У малярийных плазмодиев нашли собственные биологические часы
Цикл развития Plasmodium falciparum в эритроцитах
GovdocsGwen / flickr
Американские биологи доказали, что циклическое течение
малярии обусловлено наличием у малярийного плазмодия собственных циркадных
ритмов, которые можно наблюдать даже в культуре in vitro и во время инфекции «асинхронного»
хозяина. Результаты экспериментов изложены сразу в двух статьях в Science (1, 2).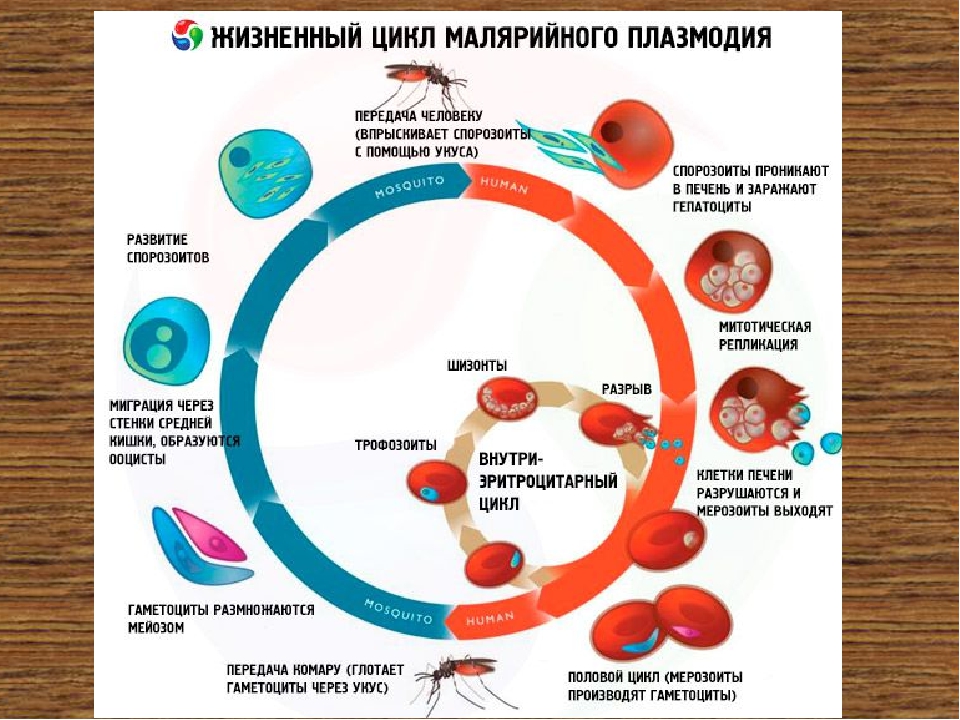
Малярия — тяжелое инфекционное заболевание, которое вызывается простейшим из рода Plasmodium. Возбудитель попадает в кровь жертвы с укусом определенного вида комара, и начинает размножаться внутри сначала клеток печени, а затем в эритроцитах — красных кровяных клетках. Течение малярии характеризуется периодической лихорадкой, которая происходит раз в 24, 48 или 72 часа. Цикличность болезни соответствует цикличности развития плазмодиев, которые одновременно разрушают эритроциты и выходят наружу.
Раньше считалось, что циклы развития паразита регулируются циркадными ритмами хозяина, однако ученые выяснили, что плазмодии в основном руководствуются собственными биологическими часами, которые, тем не менее, могут подстраиваться под часы их хозяина. Две разные исследовательские группы на двух видах плазмодия проследили за экспрессией генов паразитов и убедились в ее цикличности вне зависимости от внешних условий.
Ученые из Юго-западного медицинского центра Университета
Техаса работали на мышиной модели малярии, которая вызывается простейшим Plasmodium chabaudi, и изучали транкриптом паразита каждые 3 часа в
течение трех 24-часовых циклов.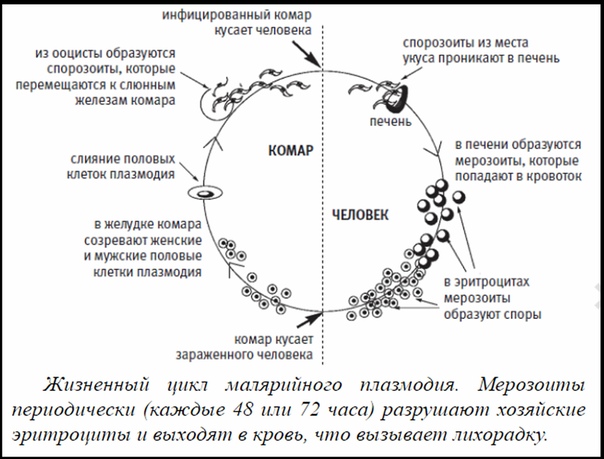 Они выяснили, что экспрессия 60 процентов генов
плазмодия подвержена регулярным колебаниям в рамках цикла 24 часа, которые
сохранились у мышей со сбитым режимом. В первом случае инфицированных мышей держали
в темноте, нарушив их режим сна, а во втором случае хаотично кормили в течение
суток, нарушив режим питания. Оказалось, что ни то, ни другое не влияет на цикл
развития плазмодия.
Они выяснили, что экспрессия 60 процентов генов
плазмодия подвержена регулярным колебаниям в рамках цикла 24 часа, которые
сохранились у мышей со сбитым режимом. В первом случае инфицированных мышей держали
в темноте, нарушив их режим сна, а во втором случае хаотично кормили в течение
суток, нарушив режим питания. Оказалось, что ни то, ни другое не влияет на цикл
развития плазмодия.
Цикличность плазмодия наблюдалась даже у мутантных мышей с
выключенным геном Cry,
у которых с рождения отсутствовал какой-либо режим дня. Тем не менее через 5-7
дней инфекции синхронность циклов паразитов у таких мышей пропала. В мутантных
мышах с удлиненными до 26 часов сутками (т.е. их ежедневная жизнедеятельность укладывалась
в 25,7 часов, в то время как у нормальных мышей в 23,7 часов) жизненный цикл
плазмодия растянулся соответственно хозяйскому. Исследователи сделали вывод,
что каждый малярийный плазмодий регулирует свой жизненный цикл самостоятельно,
однако способен воспринимать и сигналы со стороны организма хозяина, которые,
видимо, помогают синхронизировать популяцию паразитов.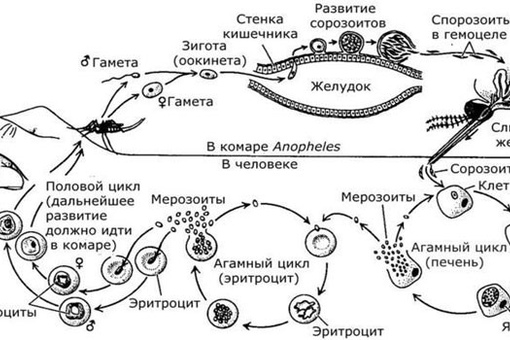
Исследовательская группа из Университета Дьюка изучала поведение четырех штаммов возбудителя человеческой малярии Plasmodium falciparum в культуре in vitro. Ученые также анализировали состав РНК плазмодиев с интервалом в 3 часа в течение 60-70 часов. Они обнаружили, что как минимум 87 процентов транскриптов плазмодия подвержено циклическим колебаниям. Нужно отметить, что генов, гомологичных основным циркадным регуляторам животных, у плазмодиев нет, тем не менее, даже в культуре их жизненный цикл составил 48 часов с небольшими вариациями от штамма к штамму.
Наблюдения ученых соответствуют существующей теории о том,
что в составе организма каждая клетка имеет собственные биологические часы
(циркадные ритмы), однако их цикл не обязательно составляет ровно 24 часа. Для
подстройки всех клеточных часов нужен «мастер-регулятор», который у млекопитающих
расположен в гипоталамусе. За открытие биологических часов и механизма работы в
2017 году была присуждена Нобелевская премия по физиологии и медицине, почитать
об этом подробнее можно в нашем материале «Ход часов лишь однозвучный».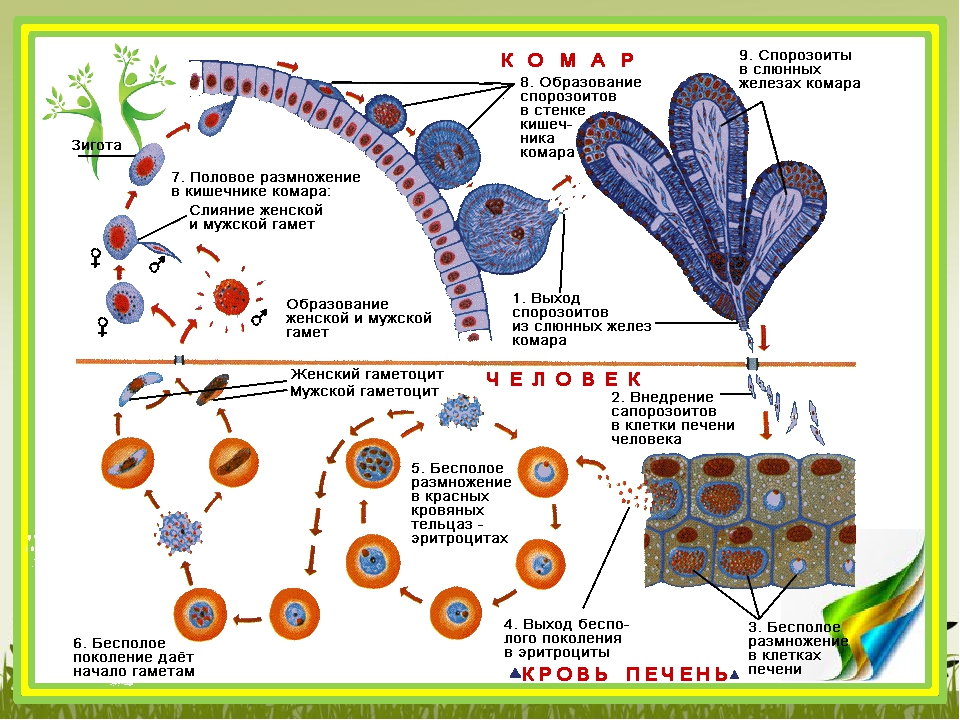
Дарья Спасская
Ход развития малярийного плазмодия / Зоология для учителя
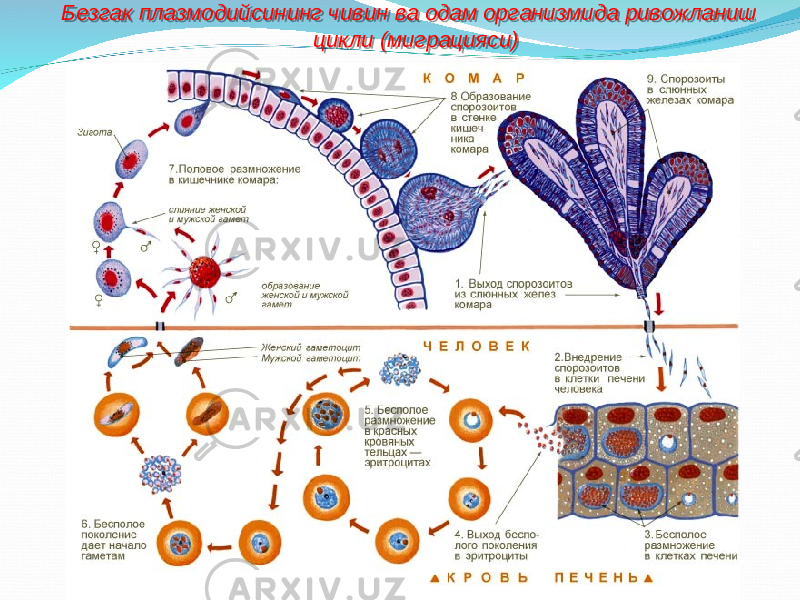
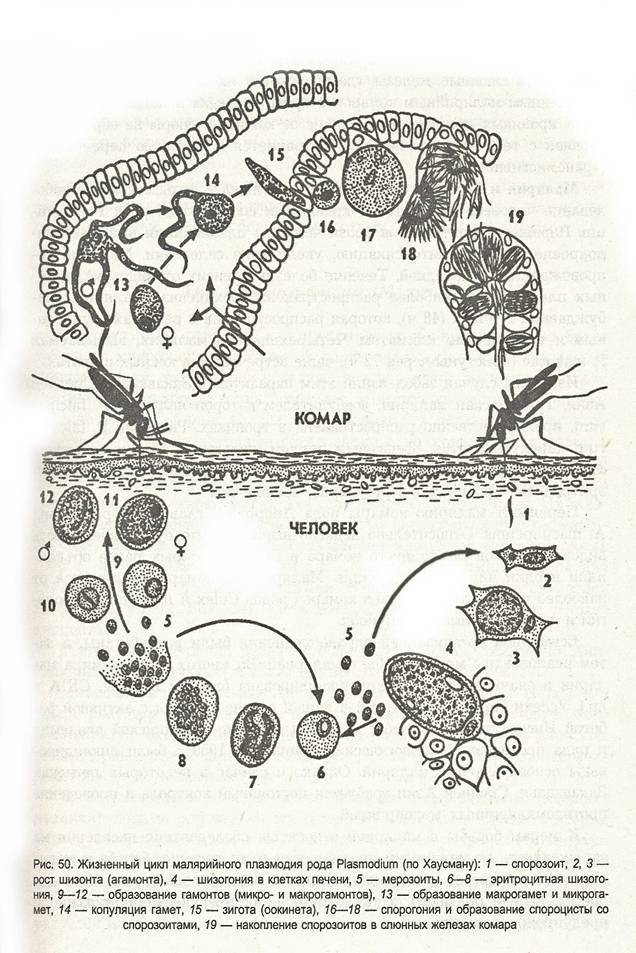
Оказалось, что возбудитель малярии — малярийный плазмодий — передаётся человеку исключительно через укусы комаров, принадлежащих к роду анофелес (см. рис. 245), и притом только в тех случаях, если такой комар сам был заражён малярийным плазмодием, насосавшись где-нибудь крови малярийного больного.
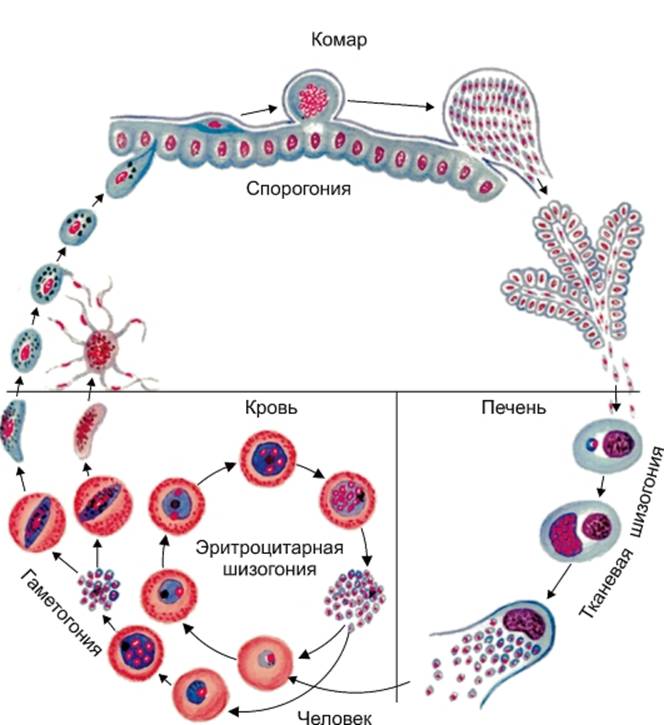
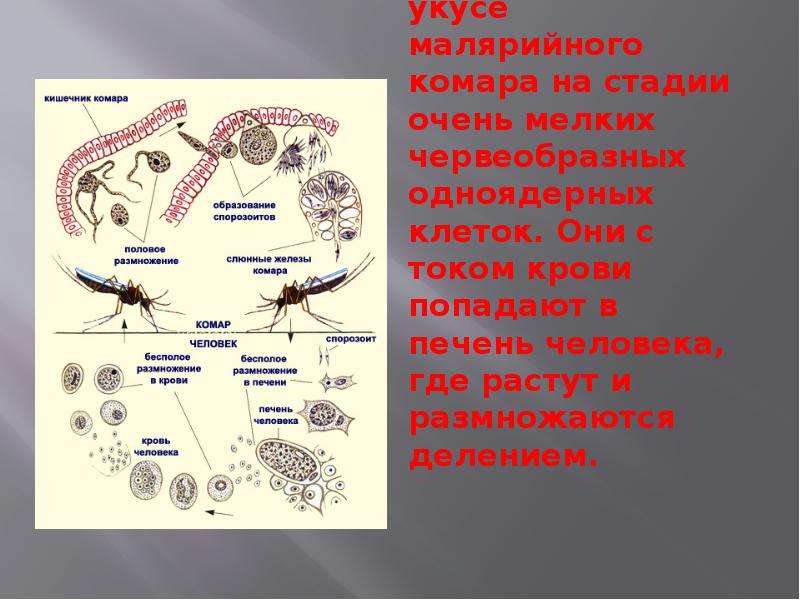
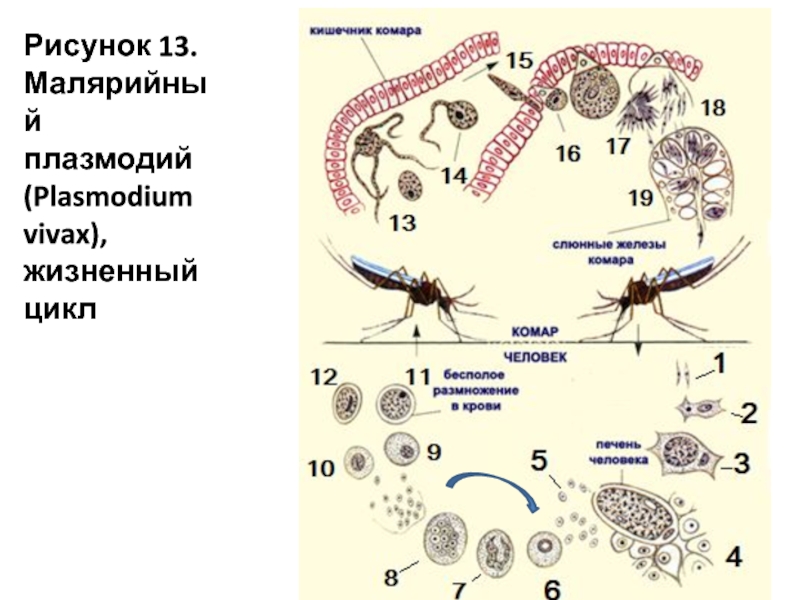
В организм человека возбудители малярии попадают.вместе со слюной заражённого комара анофелеса в виде тонких веретеновидных клеточек — так называемых спорозоитов (рис. 40, 1). Кровеносная система разносит спорозоиты по всему телу, и часть их через печёночную артерию попадает в печень и там внедряется в печёночные клетки (рис. 40, 2–9) — это пока ещё скрытый период малярии, когда заражённый человек ещё не ощущает приступов лихорадки.
В клетках печени паразиты разрастаются, а затем распадаются на мелкие дольки — мерозоиты, которые попадают в общее кровяное русло и уже в таком виде внедряются в красные кровяные тельца — эритроциты.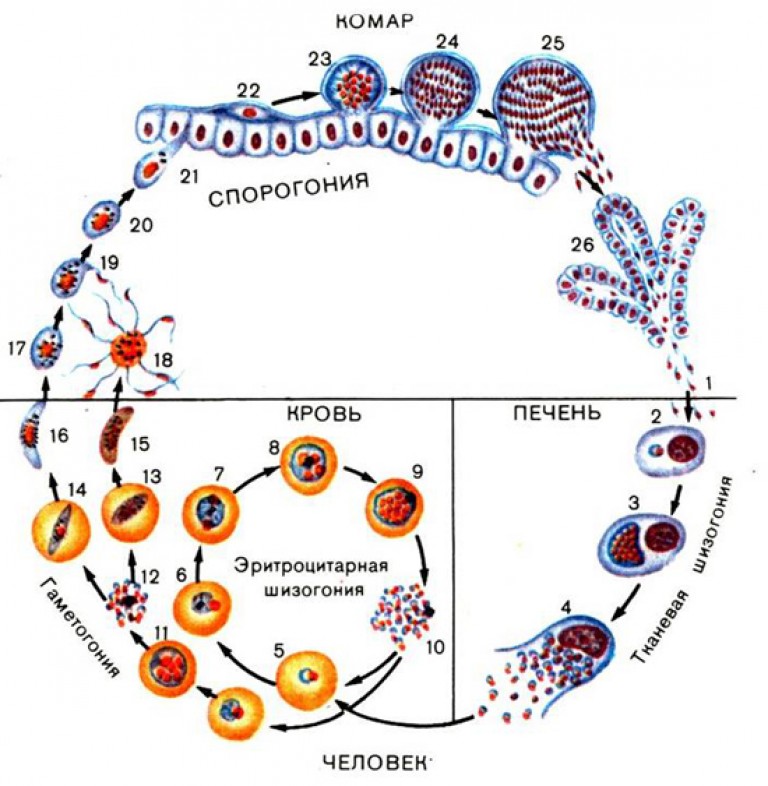 Внутри эритроцита паразит изменяет свои внешний вид (рис. 40, 10–15) и становится похожим на амёбу (отсюда и его старое название «кровяная амёба»).
Внутри эритроцита паразит изменяет свои внешний вид (рис. 40, 10–15) и становится похожим на амёбу (отсюда и его старое название «кровяная амёба»).
Он заполняет собой весь объем поражённого им кровяного тельца, а затем дробится сразу на мелкие дольки — мерозоиты (рис. 40, 16). Тогда кровяные тельца, в которых развивались паразиты, окончательно разрушаются, молодые мерозоиты попадают в кровяную жидкость (рис. 40, 16), а затем проникают в новые кровяные тельца.
Там повторяется то же самое (рис. 40, 10–16), и всякий раз, когда новые поколения мерозоитов выходят из разрушенных ими эритроцитов, больной чувствует приступ лихорадки.
Так в крови больного одни поколения плазмодиев сменяются другими, вызывая периодически повторяющиеся приступы «перемежающейся лихорадки», и этот процесс может продолжаться в течение многих лет, пока больной не излечится от своего недуга. Однако некоторая часть малярийных плазмодиев, разросшихся внутри эритроцитов, не приступает к дроблению, а превращается в особые «полулунные тельца» (рис.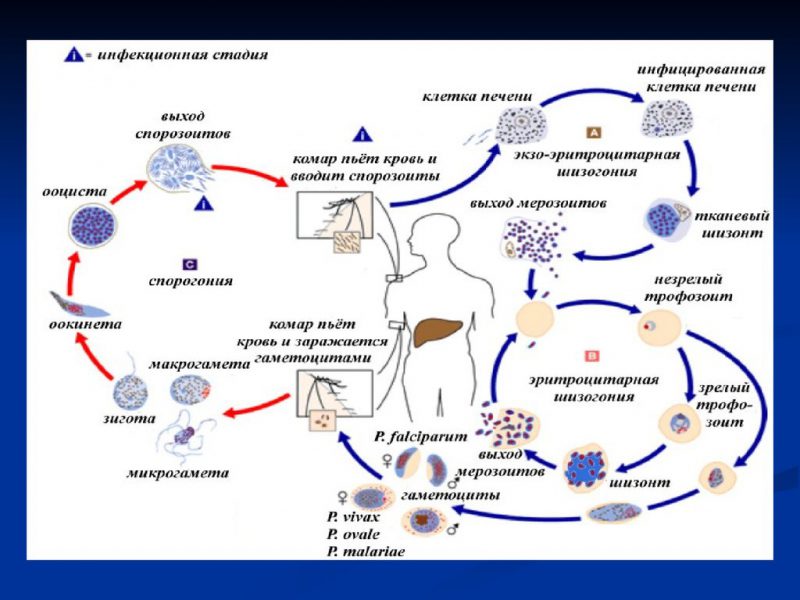 40, 17–20), и дальнейшее развитие их пока приостанавливается.
40, 17–20), и дальнейшее развитие их пока приостанавливается.
Стадии малярийного плазмодия
Много видов простейших, обитающих в различных органах человека и животных, вызывают их опасные заболевания. Одно из них — малярия, от которой на протяжении XIX и XX веков погибло свыше 100 млн людей. Возбудители малярии — несколько видов малярийных плазмодиев.
Малярийный плазмодий имеет сложный жизненный цикл, происходящий со сменой полового и бесполых поколений. Во время жизненного цикла происходит и смена хозяев — человека и малярийного комара. Комар заражается малярийным плазмодием, напившись крови человека, больного малярией. В кишечнике комара происходит половое размножение малярийного плазмодия, то есть комар — окончательный хозяин паразита.
Окончательный хозяин — организм, в котором паразит размножается половым способом.
После полового размножения наступает бесполое: из оплодотворенной яйцеклетки образуется много подвижных клеток паразита. Через некоторое время они накапливаются в слюнных железах комара, откуда при укусе насекомым вместе со слюной попадают в кровь человека.
Промежуточный хозяин — организм, в котором паразит размножается бесполым способом или проходит только определенные стадии развития.
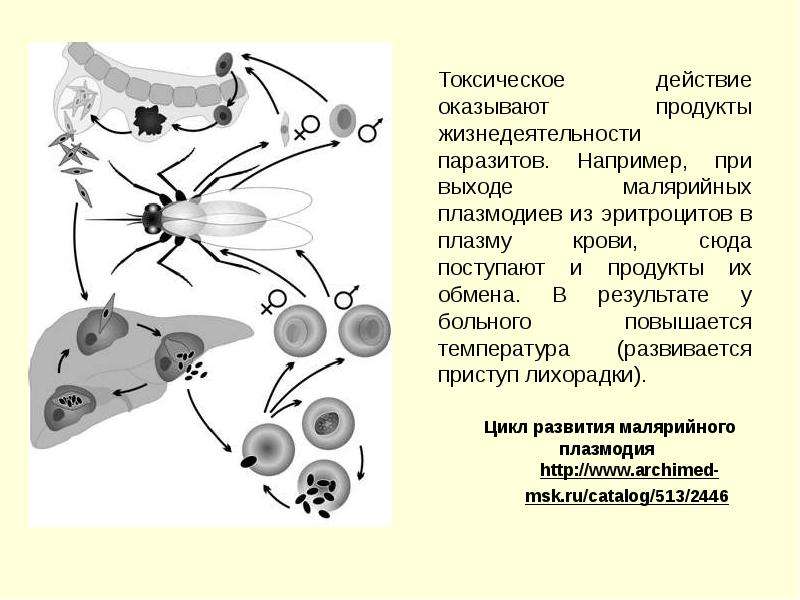
Сначала клетки паразита попадают в клетки печени человека, где интенсивно размножаются. Дальнейшее развитие паразита происходит в красных клетках крови — эритроцитах. Через определенные промежутки времени (24, 48 или 72 часа) клетки паразита разрушают эритроциты. В кровь с клетками паразита попадают токсичные продукты жизнедеятельности плазмодия, и у человека наблюдают приступ лихорадки.
Благодаря лечению больных малярией и целенаправленной борьбе с малярийными комарами и их личинками, обитающими в водоемах, малярию удалось ликвидировать на территории Европы и Северной Америки. Однако в последнее время в Украине ежегодно регистрируют сотни случаев заболевания малярией. Ее завозят люди, возвращающиеся из стран, где это заболевание встречается. Учитывая то, что в Украине обитает несколько видов малярийных комаров, в нашей стране постоянно существует опасность вспышек этого опасного заболевания.
Создан транскриптомный атлас стадий жизненного цикла малярийного плазмодия
Исследователи из института Сенгера (Кембридж) создали атлас, содержащий транскриптомные профили более чем 1700 особей Plasmodium berghei на разных стадиях жизненного цикла паразита. Ученые изолировали и очищали клетки P. berghei, взятые в разных точках жизненного цикла, начиная от момента, когда комар, укусивший млекопитающее, заражает его кровь спорозоитами. Затем паразиты собираются в печени хозяина, где развиваются и размножаются бесполым путем перед тем, как поселиться в эритроцитах. Во время эритроцитарной стадии плазмодии снова размножаются, разрывают инфицированные эритроциты изнутри и заражают новые клетки. На этой стадии они могут попасть в организмы других комаров и мигрируют к ним в среднюю кишку и слюнные железы.
Ученые утверждают, что выявили паттерны экспрессии, присутствующие у плазмодия в зависимости от стадии жизненного цикла, стратегии и хозяина.
«Мы создали атлас активности генов на протяжении всей жизни паразита. Это первый такой атлас для одноклеточного организма, — утверждает Вирджиния Ховик, постдокторант в институте Сенгера и соавтор статьи. — Жизненный цикл малярийного паразита — ключ к пониманию малярии, и клеточный атлас поможет нам разобраться в нюансах жизни плазмодия, чтобы эффективно контролировать это заболевание».
Используя модифицированную технологию секвенирования РНК под названием Smart-seq2, исследователи определили транскриптомные профили единичных клеток на каждой стадии их жизненного цикла. Они выявили экспрессию в среднем 1527 генов на клетку, с небольшими изменениями в зависимости от стадии жизненного цикла и от хозяина. К этому они добавили капельное секвенирование почти 16000 клеток малярийных паразитов на эритроцитарной стадии — представителей вида Plasmodium berghei и других, вызывающих малярию, выращенных в мышах или в культуре — P. falciparum, P. malariae, and P. knowlesi.
falciparum, P. malariae, and P. knowlesi.
Оказалось, что у разных видов плазмодия на этой стадии активируется один и тот же набор генов с небольшими различиями. Однако у паразитов, заражающих разных хозяев, эта стадия имеет различную продолжительность.
Исследователи также выделили у паразитов около 20 кластеров экспрессии генов. В двух из них находились в основном гены домашнего хозяйства (гены, необходимые для поддержания базовых жизненных функций клетки, как правило, экспрессируются на постоянном уровне), но большинство кластеров содержали гены, которые экспрессировались только на определенной стадии. Например, кластер 16 содержал гены, экспрессирующиеся в период инвазии; функции одних известны, других — возможно, будут выяснены. Предполагается, что изучение этих кластеров экспрессии поможет определить возможные мишени для лекарств, ключевые на тех или иных стадиях.
Авторы статьи считают, что этот атлас можно было бы использовать, чтобы связать транскриптомные данные с конкретными стадиями жизненного цикла паразитов, как они сделали для P.
Малярия
Малярия (Средние века итал. mala aria — «плохой воздух», ранее известная как «болотная лихорадка» ) — группа трансмиссивных инфекционных заболеваний, передаваемых человеку при укусах комаров рода Anopheles («малярийных комаров») и сопровождающихся лихорадкой, ознобами, спленомегалией (увеличением размеров селезёнки), гепатомегалией (увеличением размеров печени), анемией. Характеризуется хроническим рецидивирующим течением.
Ежегодно фиксируется 350—500 миллионов случаев заражения людей малярией, из них 1,3—3 миллиона заканчиваются смертью. 85—90 % случаев заражения приходится на районы Африки южнее Сахары, в подавляющем большинстве инфицируются дети в возрасте до 5 лет.
История
Есть предположение, что люди болеют малярией уже в течение 50 000 лет. Считается, что родиной малярии является Западная Африка (P. falciparum) и Центральная Африка (P. vivax). Молекулярно-генетические данные свидетельствуют, что предпаразитический предок плазмодия был свободноживущим простейшим, способным к фотосинтезу, который приспособился жить в кишечнике водных беспозвоночных. Также он мог жить в личинках первых кровососущих насекомых отряда Diptera, которые появились 150—200 миллионов лет назад, быстро приобретя возможность иметь двух хозяев. Древнейшие найденные окаменелости комаров с остатками малярийных паразитов имеют возраст 30 миллионов лет. С появлением человека развились малярийные паразиты, способные к смене хозяина между человеком и комарами рода Anopheles.
Выяснение причины заболевания
В 1880 году французский военный врач Шарль Луи Альфонс Лаверан, работавший в Алжире, обнаружил в кровяных шариках больного малярией живой одноклеточный организм.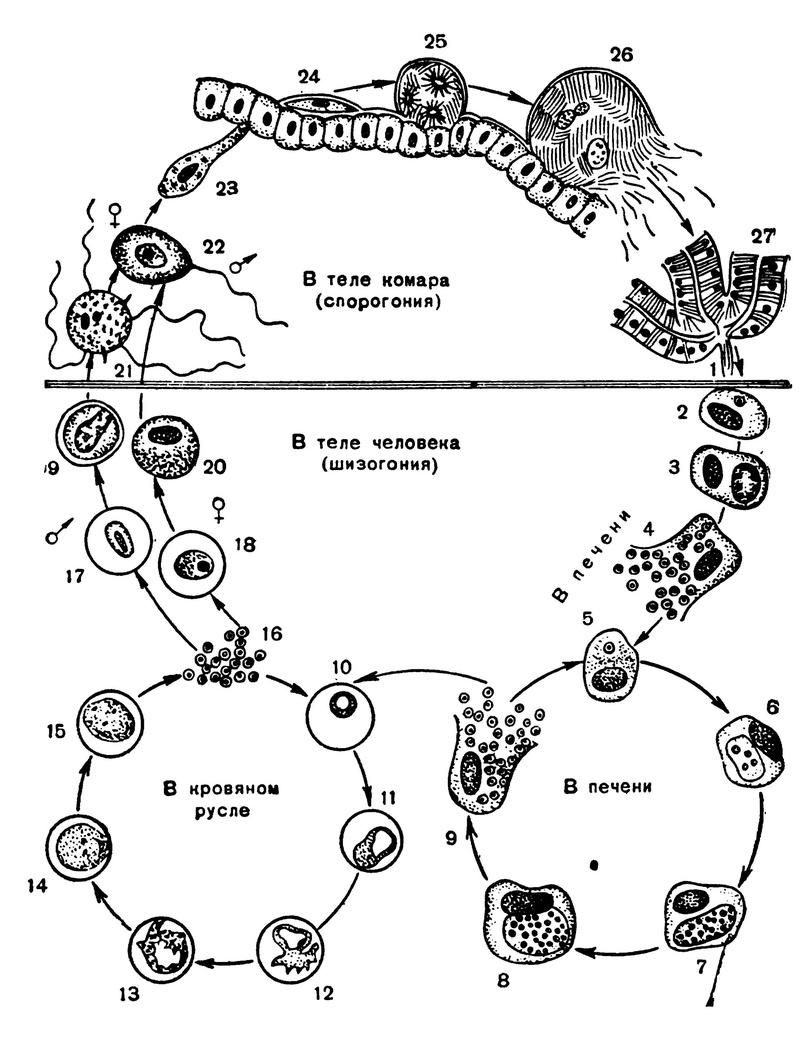 Спустя год ученый опубликовал в медицинской печати статью «Паразитарный характер заболевания малярией: описание нового паразита, найденного в крови больных малярией». Это был первый случай, когда простейшие были идентифицированы как причина болезни. За это и другие открытия он был награждён Нобелевской премией по физиологии и медицине 1907 года. Название рода паразита Plasmodium было предложено в 1895 году итальянскими учеными Этторе Маркиафава (на англ.) (итал. Ettore Marchiafava) и Анджело Челли (на англ.) (итал. Angelo Celli)[10]. В 1894 году паразитолог Патрик Мэнсон впервые предположил, что малярия может передаваться человеку комарами. В 1896 г. кубинский врач Карлос Финлей, который лечил больных жёлтой лихорадкой в Гаване, высказал ту же гипотезу. Англичанин сэр Рональд Росс, работавший в Индии, показал в 1898 году, что определённые разновидности комаров передают малярию птицам, и выделил паразитов из слюнных желез комара. Ему также удалось найти паразитов в кишечнике комаров, питавшихся кровью больных людей, но не удалось проследить передачу паразитов от комаров к человеку.
Спустя год ученый опубликовал в медицинской печати статью «Паразитарный характер заболевания малярией: описание нового паразита, найденного в крови больных малярией». Это был первый случай, когда простейшие были идентифицированы как причина болезни. За это и другие открытия он был награждён Нобелевской премией по физиологии и медицине 1907 года. Название рода паразита Plasmodium было предложено в 1895 году итальянскими учеными Этторе Маркиафава (на англ.) (итал. Ettore Marchiafava) и Анджело Челли (на англ.) (итал. Angelo Celli)[10]. В 1894 году паразитолог Патрик Мэнсон впервые предположил, что малярия может передаваться человеку комарами. В 1896 г. кубинский врач Карлос Финлей, который лечил больных жёлтой лихорадкой в Гаване, высказал ту же гипотезу. Англичанин сэр Рональд Росс, работавший в Индии, показал в 1898 году, что определённые разновидности комаров передают малярию птицам, и выделил паразитов из слюнных желез комара. Ему также удалось найти паразитов в кишечнике комаров, питавшихся кровью больных людей, но не удалось проследить передачу паразитов от комаров к человеку. Джованни Батиста Грасси в 1898 г удалось осуществить экспериментальное заражение человека малярией через укус комара (он ставил опыты на добровольцах, в том числе и на себе). Он доказал также, что только комары рода Anopheles являются переносчиками малярии в Италии, разработал и внедрил меры профилактики малярии. Однако в 1902 Нобелевскую премию по медицине за описание жизненного цикла малярийного паразита получил только Рональд Росс. Данные, полученные Финлеем и Россом, в 1900 году были подтверждены медицинским советом, который возглавлял Вальтер Рид. Рекомендации этого совета были использованы Вильямом C. Гордасом для оздоровительных мероприятий, осуществлённых на строительстве Панамского канала.
Джованни Батиста Грасси в 1898 г удалось осуществить экспериментальное заражение человека малярией через укус комара (он ставил опыты на добровольцах, в том числе и на себе). Он доказал также, что только комары рода Anopheles являются переносчиками малярии в Италии, разработал и внедрил меры профилактики малярии. Однако в 1902 Нобелевскую премию по медицине за описание жизненного цикла малярийного паразита получил только Рональд Росс. Данные, полученные Финлеем и Россом, в 1900 году были подтверждены медицинским советом, который возглавлял Вальтер Рид. Рекомендации этого совета были использованы Вильямом C. Гордасом для оздоровительных мероприятий, осуществлённых на строительстве Панамского канала.
В начале XX века, до открытия антибиотиков, практиковалось умышленное заражение малярией пациентов, больных сифилисом. Малярия обеспечивала повышенную температуру тела, при которой сифилис если и не проходил полностью, то во всяком случае снижал свою активность и переходил в латентную стадию.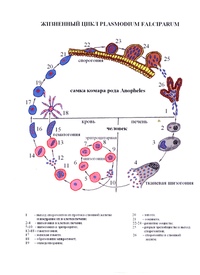 Контролируя течение лихорадки с помощью хинина, врачи таким образом пытались минимизировать негативные эффекты сифилиса. Несмотря на то, что некоторые пациенты умирали, это считалось предпочтительнее неизбежной смерти от сифилитической инфекции.
Контролируя течение лихорадки с помощью хинина, врачи таким образом пытались минимизировать негативные эффекты сифилиса. Несмотря на то, что некоторые пациенты умирали, это считалось предпочтительнее неизбежной смерти от сифилитической инфекции.
Открытие покоящейся стадии паразита
Хотя стадии жизненного цикла паразита, проходящие в кровяном русле человека и в теле комара, были описаны ещё в конце XIX — начале XX века, только в 1980-е годы стало известно о существовании покоящейся стадии. Открытие этой формы паразита окончательно объяснило, как люди, вылечившиеся от малярии, могли вновь заболевать спустя годы после исчезновения клеток плазмодия из кровяного русла.
Ареал
Малярийные комары живут почти во всех климатических зонах, за исключением субарктического, арктического поясов и пустынь. В России они обитают на всей европейской территории страны и в Западной Сибири, кроме полярных и приполярных широт. В Восточной Сибири не обитают: зимы там слишком суровые, и комары не выживают.
Однако для того, чтобы существовал риск заражения малярией, требуются, помимо малярийных комаров, условия для быстрого размножения их и переноса малярийного плазмодия. Такие условия достигаются в тех районах, где не бывает низких температур, имеются болота и выпадает много осадков. Поэтому малярия шире всего распространена в экваториальной и субэкваториальной зонах.
Также к естественному ареалу малярии относят влажный субтропический пояс: так, в районе Сочи малярия в начале XX века была большой проблемой до принятия мер по осушению заболоченной местности, нефтеванию водоёмов и проведению других мероприятий, приведших в итоге к уничтожению мест размножения малярийных комаров в курортной зоне.
В России и в СССР до начала 1950-х годов заболеваемость малярией была массовой, причем не только на Кавказе, в Закавказье и в Средней Азии, но и средней полосе европейской части (Поволжье и другие районы). Пик абсолютного числа заболевших пришелся на 1934—1935 годы, когда было зарегистрировано более 9 млн случаев малярии.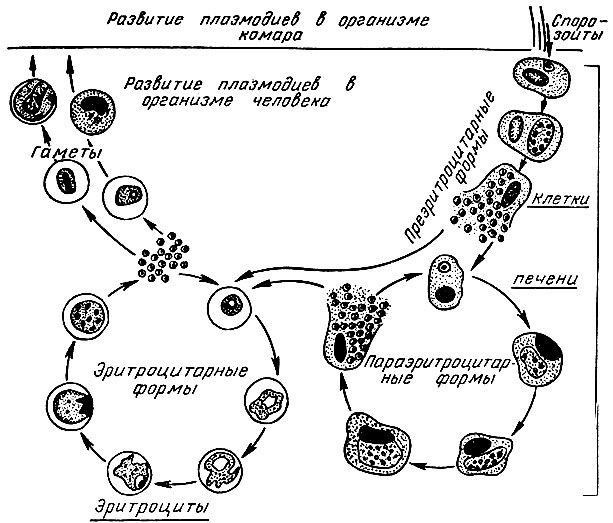
Этиология
Возбудители малярии — простейшие рода Plasmodium (плазмодии). Для человека патогенны четыре вида этого рода: P.vivax (англ.), P.ovale (англ.), P.malariae (англ.) и P.falciparum. В последние годы установлено, что малярию у человека в Юго-Восточной Азии вызывает также пятый вид — Plasmodium knowlesi. Человек заражается ими в момент инокуляции (впрыскивания) самкой малярийного комара одной из стадий жизненного цикла возбудителя (так называемых спорозоитов) в кровь или лимфатическую систему, которое происходит при кровососании.
Эритроцитарная, или клиническая стадия малярии начинается с прикрепления попавших в кровь мерозоитов к специфическим рецепторам на поверхности мембраны эритроцитов. Эти рецепторы, служащие мишенями для заражения, по-видимому, различны для разных видов малярийных плазмодиев.
Плазмодии, попадая в жертву, стимулируют выбросы ей веществ, являющихся привлекательными для комаров. К такому выводу пришли исследователи, когда провели серию экспериментов на мышах.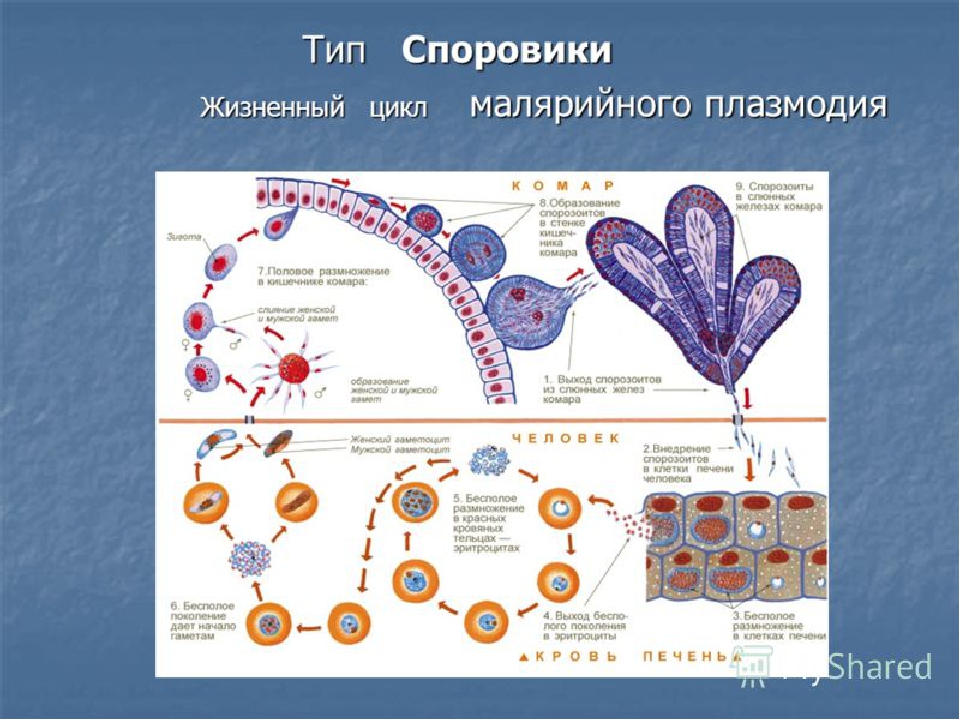 Малярийные паразиты изменяли запах тела мышей, и особенно «привлекательным» этот запах становился в период их (паразитов) полного созревания.
Малярийные паразиты изменяли запах тела мышей, и особенно «привлекательным» этот запах становился в период их (паразитов) полного созревания.
Симптомы и диагностика
Симптомы малярии обычно следующие: лихорадка, ознобы, артралгия (боль в суставах), рвота, анемия, вызванная гемолизом, гемоглобинурия (выделение гемоглобина в моче) и конвульсии. Возможно также ощущение покалывания в коже, особенно в случае малярии, вызванной P. falciparum. Также могут наблюдаться спленомегалия (увеличенная селезенка), нестерпимая головная боль, ишемия головного мозга. Малярийная инфекция смертельно опасна. Особенно уязвимы дети и беременные женщины.
Диагноз ставится на основе выявления паразитов в мазках крови. Традиционно используют два типа мазков — тонкий и толстый (или так называемую «толстую каплю»). Тонкий мазок позволяет с большей надёжностью определить разновидность малярийного плазмодия, поскольку внешний вид паразита (форма его клеток) при данном типе исследования лучше сохраняется. Толстый мазок позволяет микроскописту просмотреть больший объём крови, поэтому этот метод чувствительнее, но внешний вид плазмодия при этом изменяется, что не позволяет легко различать разновидности плазмодия. Поставить диагноз на основе микроскопического исследования зачастую бывает затруднительно, так как незрелые трофозоиты разных видов малярийного плазмодия плохо различимы, и обычно необходимо несколько плазмодиев, находящихся на разных стадиях созревания, для надёжной дифференциальной диагностики.
Толстый мазок позволяет микроскописту просмотреть больший объём крови, поэтому этот метод чувствительнее, но внешний вид плазмодия при этом изменяется, что не позволяет легко различать разновидности плазмодия. Поставить диагноз на основе микроскопического исследования зачастую бывает затруднительно, так как незрелые трофозоиты разных видов малярийного плазмодия плохо различимы, и обычно необходимо несколько плазмодиев, находящихся на разных стадиях созревания, для надёжной дифференциальной диагностики.
В настоящее время используются также быстрые диагностические тесты (RDT, Rapid Diagnostic Tests) с использованием иммунохимических наборов (более дорогие, но дающие результат через 5—15 минут и не требующие использования микроскопа) и тесты с помощью ПЦР (наиболее дорогие, но наиболее надежные)
Виды (формы) малярии
Симптоматика, течение и прогноз заболевания отчасти зависят от вида плазмодия, который является возбудителем данной формы болезни.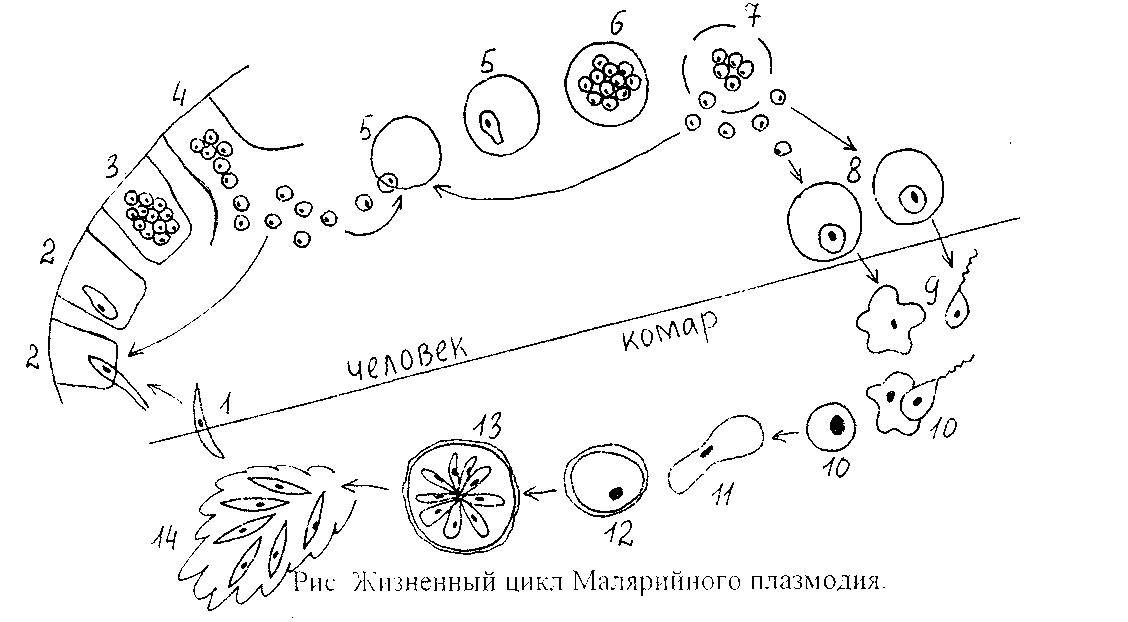
- Возбудитель тропической малярии — P. falciparum. Вызывает наиболее опасную форму, часто протекающую с осложнениями и имеющую высокую смертность. Эта же форма наиболее широко распространена (91 % всех случаев малярии в 2006 г.).
- Возбудитель четырехдневной малярии — Plasmodium malariae. Приступы происходят обычно через 72 часа.
- Возбудители трехдневной малярии и похожей на неё овале-малярии — соответственно, Plasmodium vivax и Plasmodium ovale. Приступы происходят через каждые 40—48 часов.
Эти формы малярии различаются также по длительности инкубационного периода, продолжительности разных стадий жизненного цикла плазмодиев, симптоматике и течению.
Противомалярийный иммунитет
Иммунный ответ против малярийной инфекции развивается медленно. Он характеризуется малой эффективностью и практически не защищает от повторного инфицирования. Приобретённый иммунитет развивается после нескольких заболеваний малярией за несколько лет.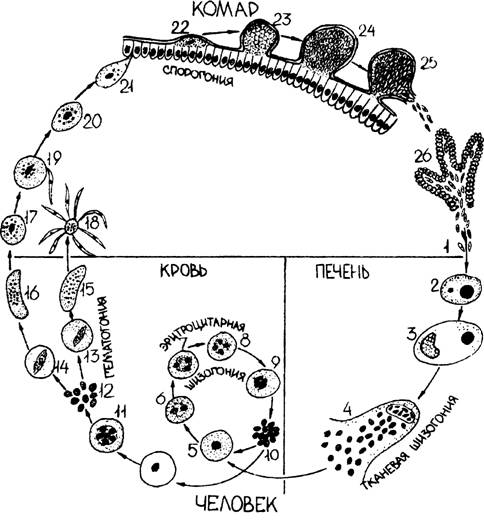 Этот иммунитет специфичен к стадии заболевания, к виду и даже к конкретному штамму малярийного плазмодия. Но клинические проявления и симптомы уменьшаются с развитием специфического противомалярийного иммунитета.
Этот иммунитет специфичен к стадии заболевания, к виду и даже к конкретному штамму малярийного плазмодия. Но клинические проявления и симптомы уменьшаются с развитием специфического противомалярийного иммунитета.
Среди возможных объяснений такого слабого иммунного ответа называют нахождение малярийного плазмодия в клетках на протяжении большей части его жизненного цикла, общее угнетение иммунной системы, присутствие антигенов, которые не опознаются T-клетками, подавление пролиферации B-клеток, значительный полиморфизм малярийного плазмодия и быстрая смена потенциальных антигенов на его поверхности.
Лечение
Самым распространенным медикаментом для лечения малярии сегодня, как и раньше, является хинин. На некоторое время он был заменен хлорохином, но ныне снова приобрел популярность. Причиной этому стало появление в Азии и затем распространение по Африке и другим частям света Plasmodium falciparum с мутацией устойчивости к хлорохину.
Также существует несколько других веществ, которые используются для лечения и иногда для профилактики малярии.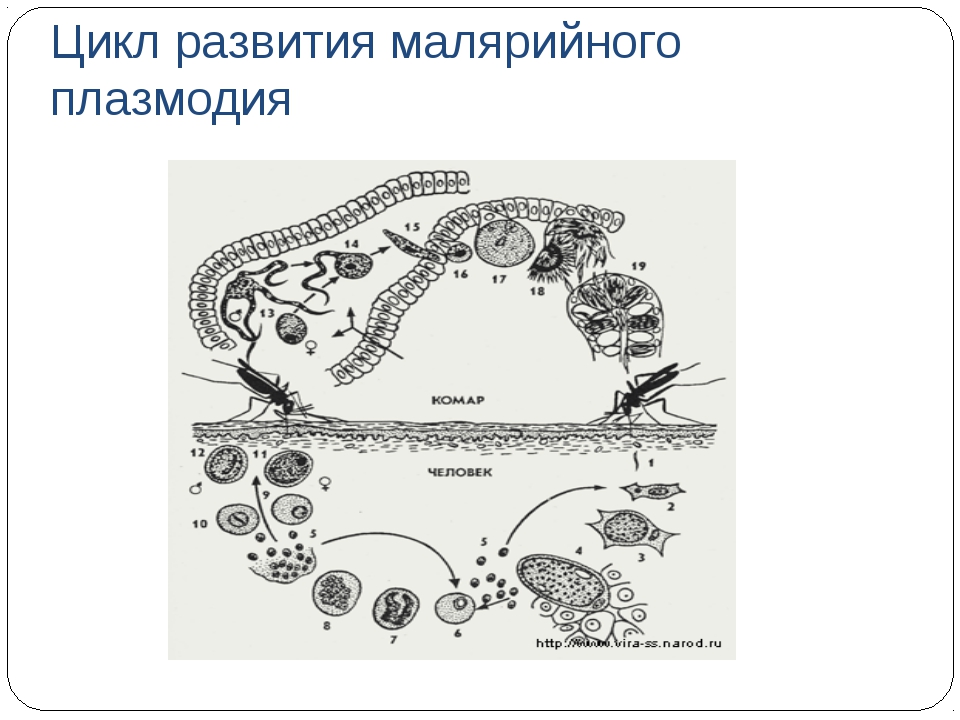 Многие из них могут использоваться для обеих целей. Их использование зависит преимущественно от устойчивости к ним паразитов в области, где используется тот или другой препарат.
Многие из них могут использоваться для обеих целей. Их использование зависит преимущественно от устойчивости к ним паразитов в области, где используется тот или другой препарат.
В настоящее время наиболее эффективными лекарственными средствами для лечения являются комбинированные с артемезинином лекарственные средства. Резолюция ВОЗ WHA60.18 (май 2007) настаивает на использовании этих препаратов, однако в РФ они до сих пор не зарегистрированы и не применяются.
| Основные антималярийные препараты | ||||
| Препарат | Англ. название | Профилактика | Лечение | Примечания |
| Артеметер-люмефантрин | Artemether-lumefantrine | — | + | коммерческое название Коартем |
| Артезунат-амодиахин | Artesunate-amodiaquine | + | — | |
| Атовакуон-прогуанил | Atovaquone-proguanil | + | + | коммерческое название Маларон |
| Хинин | Quinine | — | + | |
| Хлорохин | Chloroquine | + | + | после появления резистентности использование ограничено коммерческое название Делагил |
| Котрифазид | Cotrifazid | + | + | |
| Доксициклин | Doxycycline | + | + | |
| Мефлохин | Mefloquine | + | + | коммерческое название Лариам |
| Прогуанил | Proguanil | — | + | |
| Примахин | Primaquine | + | — | |
| Сульфадоксин-пириметамин | Sulfadoxine-pyrimethamine | + | + | коммерческое название Фансидар |
Экстракты растения Artemisia annua (Полынь однолетняя), которые содержат вещество артемизинин и его синтетические аналоги, имеют высокую эффективность, но их производство дорого.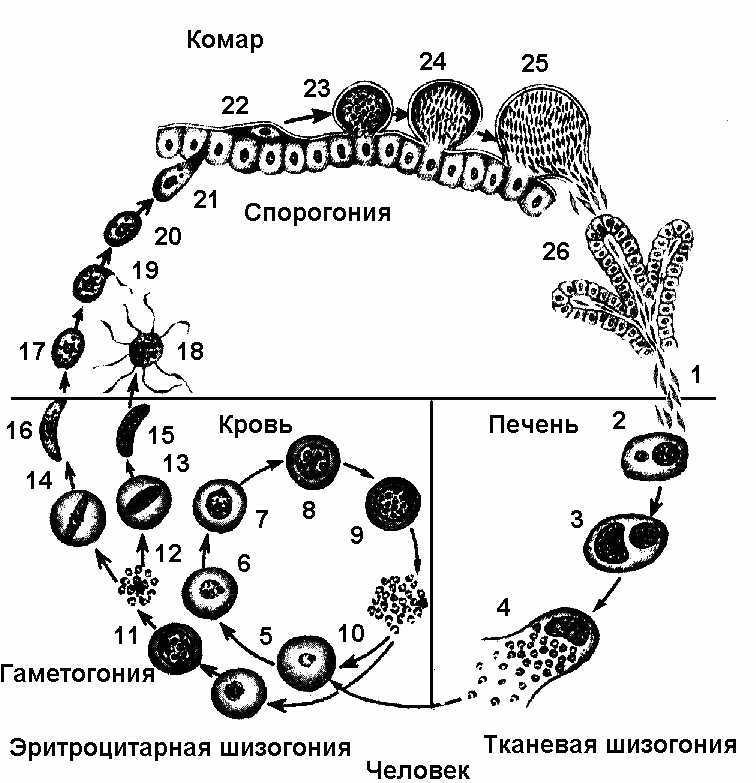 В настоящее время (2006) изучаются клинические эффекты и возможность производства новых препаратов на основе артемизинина. Другая работа команды французских и южноафриканских исследователей разработала группу новых препаратов, известных как G25 и TE3, успешно испытанных на приматах.
В настоящее время (2006) изучаются клинические эффекты и возможность производства новых препаратов на основе артемизинина. Другая работа команды французских и южноафриканских исследователей разработала группу новых препаратов, известных как G25 и TE3, успешно испытанных на приматах.
Хотя противомалярийные лекарства присутствуют на рынке, болезнь представляет угрозу для людей, которые живут в эндемичных областях, где нет надлежащего доступа к эффективным препаратам. Согласно данным организации «Врачи без границ», средние затраты на лечение человека, зараженного малярией, в некоторых африканских странах составляют всего 0,25—2,40 $.
Профилактика
Методы, которые используются для предотвращения распространения болезни или для защиты в областях, эндемичных для малярии, включают профилактические лекарственные средства, уничтожение комаров и средства для предотвращения укусов комаров. В настоящий момент нет вакцины против малярии, но ведутся активные исследования для её создания.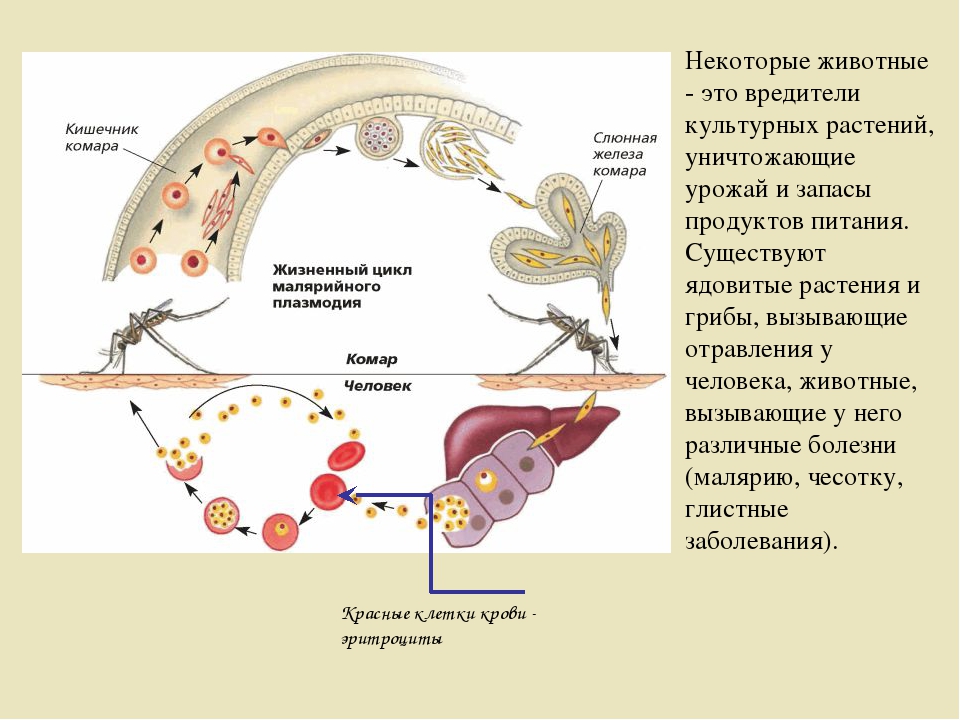
Разработка вакцины
Ведутся разработки и начаты клинические испытания вакцин от малярии.
В марте 2013 года, после серии неудачных экспериментов, ученые из США успешно испытали на мышах быстродействующее средство против малярии, новый препарат готовят к испытанию на людях.
Малярия всегда была и остается одной из опаснейших болезней человека. К известным личностям, которые умерли от малярии, принадлежат: Александр Македонский, Аларих (король вестготов), Чингисхан, святой Августин, как минимум 5 римских пап, итальянский поэт Данте, император Священной Римской империи Карл V, Христофор Колумб, Оливер Кромвель, Микеланджело Меризи Караваджо, лорд Байрон и многие другие.
Современные данные о малярии:
- В районах распространения малярии проживает 2,4 миллиарда человек, или 34 % населения мира.
- Ежегодно 300—500 миллионов человек заражается малярией, и, согласно ВОЗ, это число ежегодно возрастает на 16 %.
 90 % случаев регистрируется в Африке, из остальных — 70 % случаев приходится на Индию, Бразилию, Шри-Ланку, Вьетнам, Колумбию и Соломоновы Острова.
90 % случаев регистрируется в Африке, из остальных — 70 % случаев приходится на Индию, Бразилию, Шри-Ланку, Вьетнам, Колумбию и Соломоновы Острова. - Ежегодно 1,5—3 миллиона человек умирает от малярии (в 15 раз больше, чем от ВИЧ/СПИД).
- За последнее десятилетие с третьего места по числу смертельных случаев за год (после пневмонии и туберкулёза) малярия вышла на первое среди инфекционных заболеваний.
- Каждый год около 30 000 человек, посещающих опасные районы, заболевают малярией, 1 % из них умирает.
 90 % случаев регистрируется в Африке, из остальных — 70 % случаев приходится на Индию, Бразилию, Шри-Ланку, Вьетнам, Колумбию и Соломоновы Острова.
90 % случаев регистрируется в Африке, из остальных — 70 % случаев приходится на Индию, Бразилию, Шри-Ланку, Вьетнам, Колумбию и Соломоновы Острова.Составлен атлас активности генов возбудителя малярии
Международный исследовательский коллектив под руководством учёных из Института Сенгера (Великобритания) сделал первое в истории описание генной активности малярийного плазмодия на всех стадиях его сложного жизненного цикла. Полученные знания помогут разработать принципиально новые способы борьбы с малярией.
Возбудитель малярии — одноклеточное существо, малярийный плазмодий, паразитирующее внутри клеток крови. Плазмодиев существует более 250 видов, но, к счастью, они узкоспецифичны, и болезнь человека вызывают лишь пять: Plasmodium vivax, Plasmodium falciparum, Plasmodium ovale, Plasmodium knowlesi и Plasmodium malariae. О развитии плазмодия в организмах комара и человека можно узнать из отдельного очерка. Первая часть нынешнего исследования была сделана на Plasmodium berghei, обитающем в крови крыс, вторая – Plasmodium falciparum и Plasmodium knowlesi, полученных от трёх кенийцев, больных малярией.
Плазмодиев существует более 250 видов, но, к счастью, они узкоспецифичны, и болезнь человека вызывают лишь пять: Plasmodium vivax, Plasmodium falciparum, Plasmodium ovale, Plasmodium knowlesi и Plasmodium malariae. О развитии плазмодия в организмах комара и человека можно узнать из отдельного очерка. Первая часть нынешнего исследования была сделана на Plasmodium berghei, обитающем в крови крыс, вторая – Plasmodium falciparum и Plasmodium knowlesi, полученных от трёх кенийцев, больных малярией.
Вирджиния Ховик (Virginia Howick) из Института Сенгера говорит: «Мы создали атлас активности генов, охватывающий полный жизненный цикл малярийного плазмодия. Это первый атлас такого типа для одноклеточных организмов. Жизненный цикл паразита малярии является ключом к исследованию этой болезни, и атлас генной активности поможет нам по-настоящему узнать её возбудителя, чтобы эффективно бороться с малярией».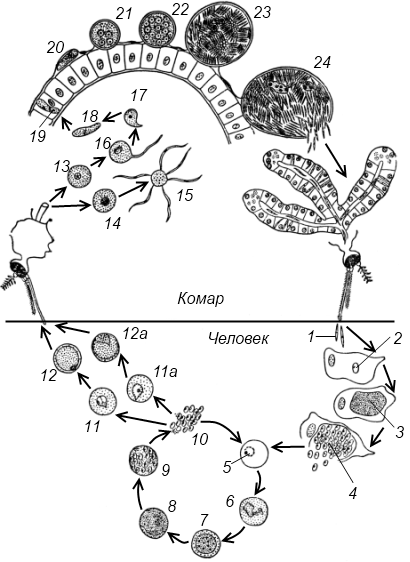
Схематическое изображение жизненного цикла малярийного плазмодия. Фото: Alex Cagan
Чтобы выполнить задачу, учёные измерили активность генов у 1787 отдельных малярийных плазмодиев на десяти стадиях развития. Например, они давали комарам, заражённым малярией, искусственную кровь, чтобы выделить плазмодиев, вышедших при питании комара, и сравнить их с теми, что остались в слюнных железах. В результате выяснилось, что у 40 % генов плазмодия, активных на разных этапах его жизни, функция пока остаётся неизвестной. Возможно, некоторые из этих генов могут стать мишенями новых методов терапии. Полученные результаты авторы исследования сделали доступными для всех коллег, опубликовав атлас генной активности малярийного плазмодия на сайте Института Сенгера.
Статья об исследовании опубликована в журнале Science.
виды, строение, размножение и цикл развития
Особенности организации Простейших
Плазмодий является представителем самой примитивной группы животных — подцарства Одноклеточные или Простейшие. Для них характерны следующие признаки:
Для них характерны следующие признаки:
- тело состоит из одной клетки, которая выполняет функции целого организма;
- наличие ядра;
- отсутствие плотной клеточной стенки;
- передвижение с помощью специализированных структур: ресничек, ложноножек, жгутиков;
- наличие пищеварительных и сократительных вакуолей;
- осуществление газообмена через поверхность клетки;
- половое и бесполое размножение.
 youtube.com/embed/zcJbMQHu1L8?feature=oembed» frameborder=»0″ allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»/>
youtube.com/embed/zcJbMQHu1L8?feature=oembed» frameborder=»0″ allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»/>Паразитические Одноклеточные
Простейшие освоили многие среды обитания: почву, пресные и соленые водоемы, болота. Среди них есть также группа паразитических одноклеточных. Они развиваются в различных органах животных и человека, при этом вызывая серьезные заболевания.
Одним из них является малярийный плазмодий. Размеры этого паразита микроскопические. В зависимости от вида клетка плазмодия может иметь вид шара, вытянутого тяжа или перстня. Всего в природе их насчитывают около 180. Но в организме человека паразитируют 10 видов, из них 5 — самые опасные.
Всего в природе их насчитывают около 180. Но в организме человека паразитируют 10 видов, из них 5 — самые опасные.
Лечение малярии
Основная задача терапии этого заболевания заключается в предотвращении возникновения/повторения приступов, полном уничтожении возбудителя. Болезнь малярия или болотная лихорадка чаще встречается в эндемичных районах, поэтому путешественникам следует принимать профилактические меры заранее. Лечение малярии проводят при помощи медикаментозной терапии, используют, как правило, Примахин, Хлорохин, Атабрин (хинакрин гидрохлорид), Акрихин.
Лекарства от малярии
Медикаментозная терапия при данном заболевании считается эффективным методом. Существуют проверенные лекарства от малярии, которые применяются на протяжении длительного времени. Примером таких медикамента выступает Хинин, который был заменен Хлорохином на некоторое время, но затем снова начал активно использоваться. Причиной этому послужило возникновение, а затем распространение в Азии и Африке Plasmodium falciparum, который имел резистентность к Хлорохину.
Примером таких медикамента выступает Хинин, который был заменен Хлорохином на некоторое время, но затем снова начал активно использоваться. Причиной этому послужило возникновение, а затем распространение в Азии и Африке Plasmodium falciparum, который имел резистентность к Хлорохину.
В зависимости от региона, где произошло заражение, могут быть использованы те или иные препараты против плазмодия малярийного. Большинство из них подходят как для лечения, так и профилактики. Экстракт полыни однолетней, содержащие артемизинин и аналоги синтетического происхождения, обладают высокой эффективностью, но и высокую стоимость. Большую опасность болезнь представляет для жителей, которые проживают в эндемических областях, где доступа к препаратам нет. В развитых странах проблем с приобретением медикаментов не наблюдается.
- Витамин A в капсулах — инструкция по применению Показания к приему витамина А в капсулах и противопоказания
- ГСГ маточных труб
- Гетры спицами описание и схемы
 youtube.com/embed/t_XfqtRg5AI?feature=oembed» frameborder=»0″ allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»/>
youtube.com/embed/t_XfqtRg5AI?feature=oembed» frameborder=»0″ allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»/>История заболевания
Малярийный плазмодий, жизненный цикл которого мы рассматриваем, вызывает тяжелое инфекционное заболевание. В переводе с итальянского языка его название означает «плохой воздух». Долгое время ее называли болотной лихорадкой.
Ученые предполагают, что первые случаи заболевания малярией были известные еще 50 тысяч лет тому назад в Центральной и Западной Африке. Эволюционные исследования привели к выводу, что предок современного вида сначала был свободноживущим. С течением времени он приспособился к обитанию в кишечнике беспозвоночных, а потом и первых кровососущих.
Причина этого смертельного заболевания долгое время была неизвестна. Только в конце XIX века французский врач Шарль Лаверан, который практиковался в Алжире, обнаружил в эритроцитах больного неизвестные клетки. За это открытие он был удостоен Нобелевской премии.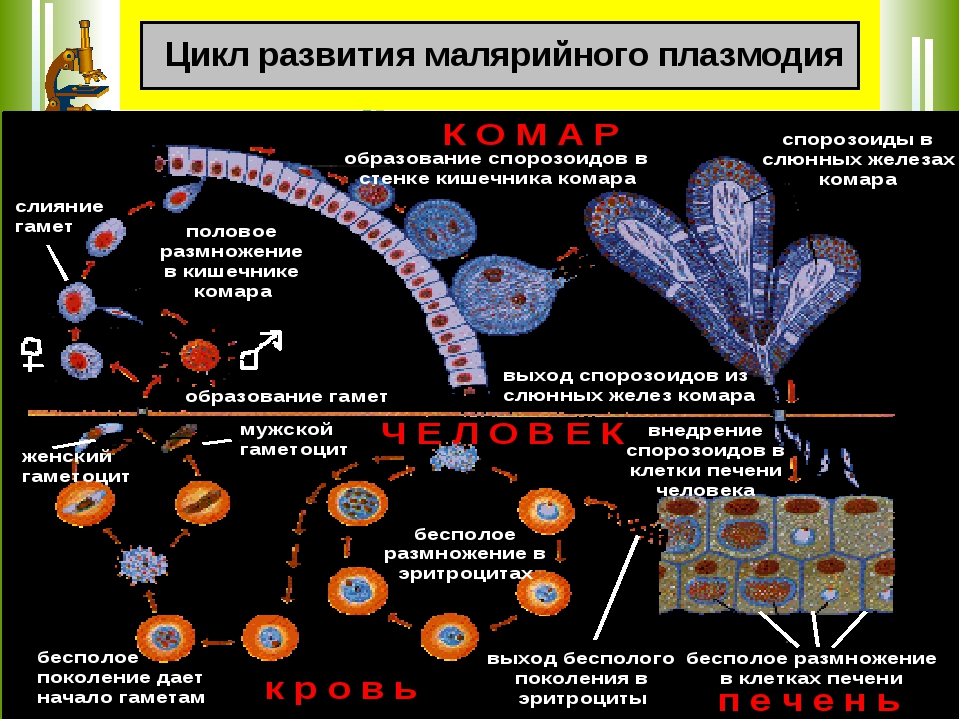
Цикл малярийного плазмодия: схема
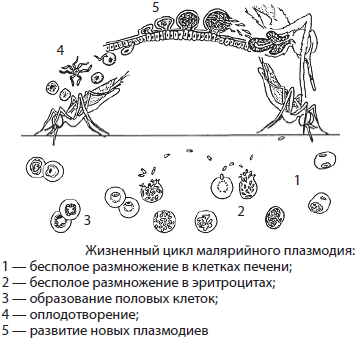
Как и все представители типа Споровики, данный паразит имеет сложный жизненный цикл. Протекает он со сменой хозяев: промежуточного и окончательного. В организме первого паразит размножается бесполым способом. Продолжается цикл развития малярийного плазмодия в теле окончательного хозяина. Там происходит половое размножение паразитического животного.
В ходе своего развития малярийный плазмодий проходит следующие стадии:
- Клетки печени человека.
- Эритроциты.
- Кровь комара.

Далее, через укус паразит вновь попадает в организм человека и цикл повторяется.
Профилактика
Важным условием проведения мероприятий против распространения малярии является своевременное выявление больных с помощью анализов, а также незамедлительное лечение их в стационаре.
Кроме того, используются следующие действия по предупреждению размножения малярийного плазмодия:
- Осушение болот, обеззараживание водоемов.
- Уничтожение комаров репеллентами.
- Защита жилых и общественных помещений специальными сетками.
- Дезинфицирующая обработка предметов и инструментов для медицинских инъекций.
Эффективной вакцины против малярии сегодня не существует, поэтому профилактические меры для предотвращения заражения плазмодием очень важны.
Развитие малярии при отсутствии лечения угрожает человеку образованием тромбов из эритроцитов, которые могут заблокировать кровеносные сосуды.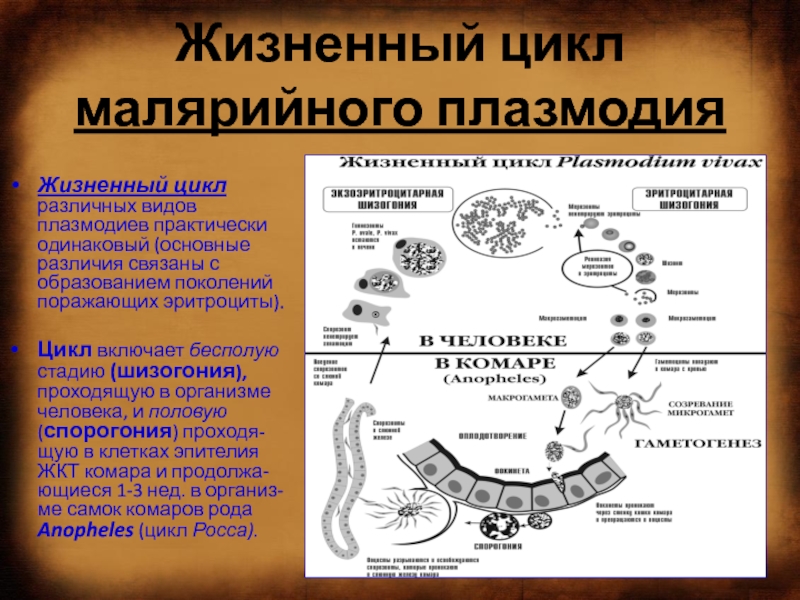 Как следствие: нарушения состава крови, гипоксия клеток печени, легких, мозга, почек. Малярийный плазмодий поражает системы кровообращения, пищеварения, дыхания. Дисфункции органов — это кома и смерть.
Как следствие: нарушения состава крови, гипоксия клеток печени, легких, мозга, почек. Малярийный плазмодий поражает системы кровообращения, пищеварения, дыхания. Дисфункции органов — это кома и смерть.
Бесполое размножение
Промежуточным хозяином паразита является человек. Именно в его организме плазмодий размножается бесполым способом. Происходит это путем деления клетки надвое.
Как попадает этот организм в кровь человека? Это происходит при укусе комара, зараженного малярийным плазмодием. Стадии его развития, которые происходят в организме промежуточного хозяина, можно разделить на несколько этапов.
Сначала плазмодий с током крови попадает в клетки печени человека. Здесь он размножается способом шизогонии. В ходе этого процесса многократно делится ядро, вокруг каждой из образовавшихся частей обосабливается цитоплазма. После этого вновь образованные клетки делятся снова.
После нескольких таких делений клетки паразита поступают в кровь. Их развитие происходит в эритроцитах.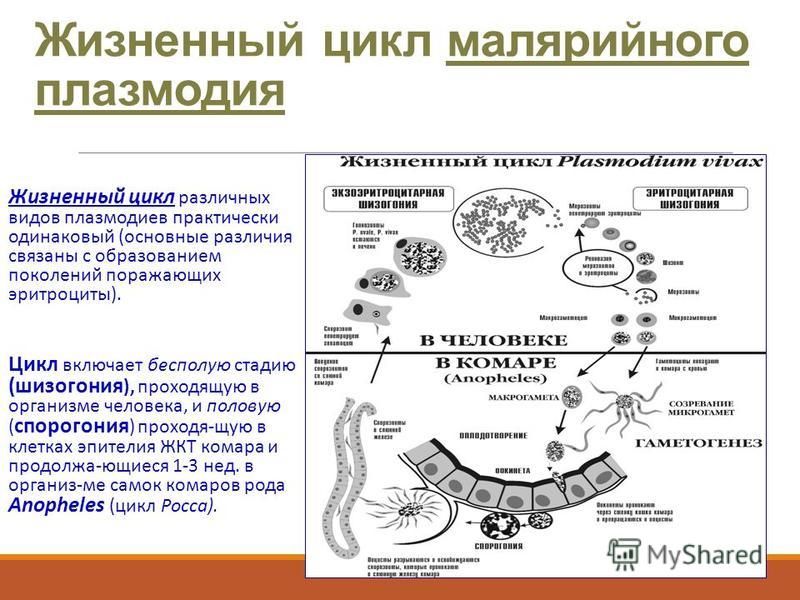 Плазмодий продолжает многократно делиться, при этом разрушая клетки крови. Их поражение наступает достаточно быстро — максимум через 72 часа.
Плазмодий продолжает многократно делиться, при этом разрушая клетки крови. Их поражение наступает достаточно быстро — максимум через 72 часа.
При разрушении эритроцитов в кровь поступают токсичные продукты жизнедеятельности плазмодия. Именно они вызывают у человека приступы лихорадки. В результате нескольких последовательных делений из клеток паразита образуются предшественники половых клеток, которые называются гаметоцитами.
[править]Этиология и эпидемиология
Малярия представляет собой паразитарную болезнь, которую вызывают 5 видов простейших из рода Plasmodium (плазмодии): Plasmodium vivax, Plasmodium ovale, Plasmodium malariae, Plasmodium falciparum и Plasmodium knowlesi.
Plasmodium vivax (Grassi et Feletti, 1890) (Labbe, 1899) — возбудитель трёхдневной малярии; Plasmodium malariae (Laveran, 1881) (Grassi et Feletti, 1890) — возбудитель четырёхдневной малярии; Plasmodium falciparum (Welch, 1897) — возбудитель тропической малярии; Plasmodium ovale (Stephens, 1922) — возбудитель овале малярияи.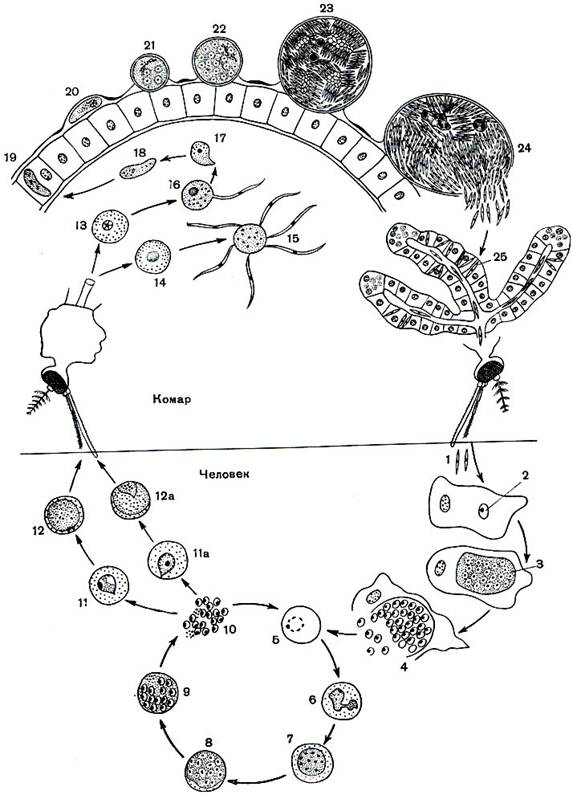
Plasmodium knowlesi представляет собой новый вид патогенных для человека плазмодиев. Он поражает людей в Юго-Восточной Азии.
Возбудители Малярия различаются по ряду признаков: вирулентности, устойчивости к воздействию химиотерапевтических препаратов, длительности инкубационного периода, иммунологический характеристикам, способности заражать комаров и пр.
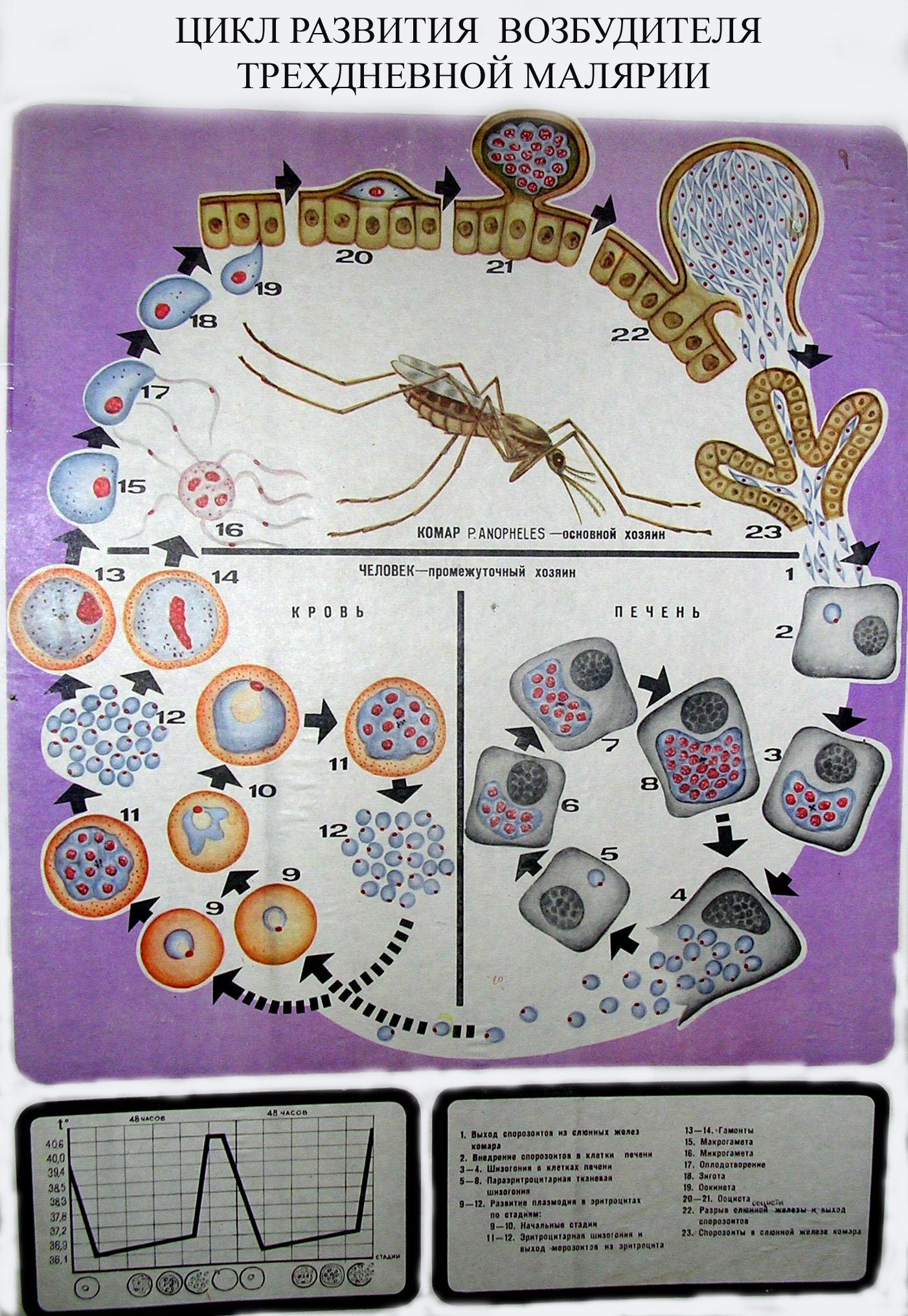
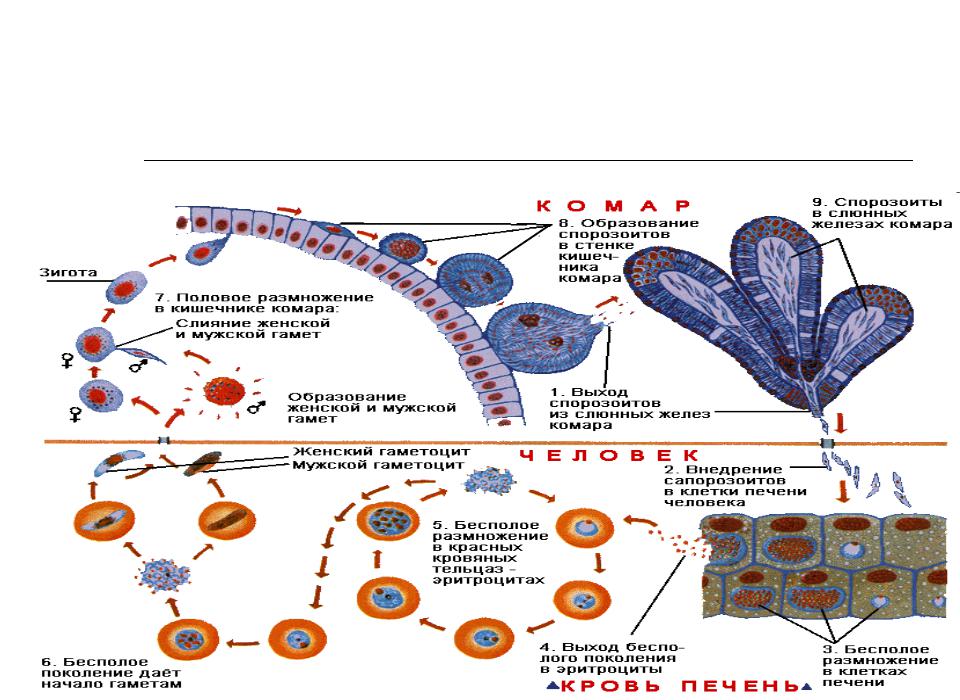
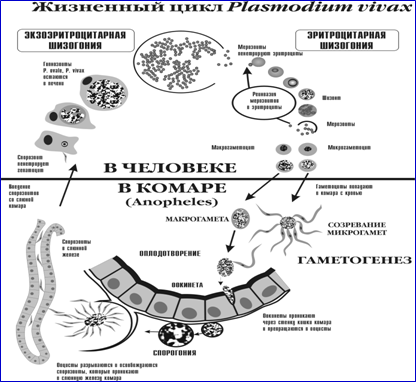
Возбудители инфекции характеризуются сложным циклом развития со сменой хозяина. Бесполое развитие (шизогония) происходит в организме человека, половое развитие (спорогония) — в организме самок комаров рода Anopheles.
В организме человека различают две фазы развития возбудителей малярии: в клетках печени — экзоэритроцитарную (внеэритроцитарную, тканевую) шизогонию и в эритроцитах — эритроцитарнуюшизогонию.
Человек заражается в момент инокуляции (впрыскивания) самкой малярийного комара одной из стадий жизненного цикла возбудителя (так называемых спорозоитов) в кровь или лимфатическую систему, которое происходит при кровососании.
С кровью и лимфой спорозоиты заносятся в печень и внедряются в гепатоциты, округляются и превращаются в трофозоиты (начало экзоэритроцитарной шизогонии). Здесь они затем превращаются в экзоэритроцитарные шизонты.
Шизонт созревает в зависимости от вида плазмодия в течение 5−15 суток. Ядро и цитоплазма шизонта многократно делятся, образуя до 10−50 тысяч экзоэритроцитарных мерозоитов овальной или удлинённой формы, длиной около 2,5 мкм и шириной 5 мкм, которые выходят в плазму крови.
У возбудителей малярии человека экзоэритроцитарная фаза развития ограничена одной генерацией. Продолжительность её у P. falciparum около 6 суток, у P. vivax — 8, у P. ovale — 9, у P. malariae — 15 суток.
Малярия — трансмиссивная болезнь, её передают человеку членистоногие переносчики — малярийные комары Anopheles, имеющих широкое распространение (из материков их нет только в Антарктиде). Известно более 450 видов этих комаров. Обитают малярийные комары и на территории РФ (примерно 10 видов), что обуславливает угрозу возвращения этой инфекции в Россию[2].

Также возможно заражение при переливании крови.
Окончательный хозяин
Цикл развития малярийного плазмодия продолжается в организме комара. Он заражается паразитом при укусе людей, больных малярией. Комар является окончательным хозяином, поскольку в нем опасные клетки размножается половым путем.
При укусе гаметоциты из эритроцитов проникают в кровь комара. Здесь они превращаются в полноценные половые клетки. Процесс оплодотворения происходит в кишечнике насекомого. В его результате образуются многочисленные подвижные клетки паразита.
В его результате образуются многочисленные подвижные клетки паразита.
Далее, жизненный цикл малярийного плазмодия повторяется. Его клетки накапливаются в слюнных железах комара, а при укусе снова попадают в кровеносное русло человека.
Опасные симптомы
Род Малярийные комары насчитывает более 400 видов. Не стоит думать, что их представители обитают только в странах Африки. Они распространены повсеместно, кроме северных районов. Характерными особенностями взрослых насекомых является вытянутое тело, длинные ноги и хоботок, короткая голова. Их крылья вдоль жилок покрыты чешуйками.
Укус такого комара является наиболее распространенным путем заражения малярией. Но причиной могут стать и нестерильные медицинские инструменты. Первым симптомом заболевания является появление лихорадки. По мере разрушения эритроцитов наблюдается увеличение размеров селезенки, уплотнение печени, развитие малокровия.
Итак, для жизненного цикла малярийного плазмодия характерна смена хозяев: промежуточного и окончательного.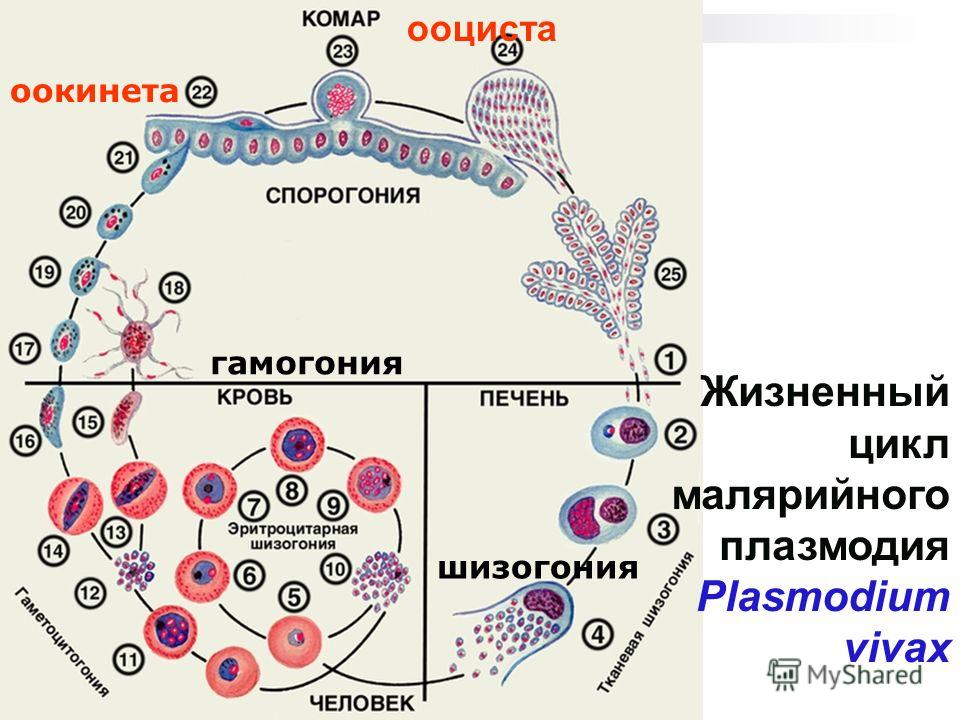 Первым является человек. В клетках его крови происходит бесполое размножение плазмодия путем шизогонии. Окончательный хозяин паразита — комар. В его организме клетки паразита размножаются половым способом. Развитие малярии происходит только в том случае, если есть возможность повторения этапов жизненного цикла. В противном случае опасная болезнь не развивается.
Первым является человек. В клетках его крови происходит бесполое размножение плазмодия путем шизогонии. Окончательный хозяин паразита — комар. В его организме клетки паразита размножаются половым способом. Развитие малярии происходит только в том случае, если есть возможность повторения этапов жизненного цикла. В противном случае опасная болезнь не развивается.
Источники малярийной инфекции
Малярийный плазмодий относится к типу Apicomplexa (класса Споровики). Комары с плазмодиями, опасными для человека, обитают в странах, где умеренные, тропические, субтропические климатические условия. Минимальная суточная температура, благоприятная для размножения + 16 градусов.
Источниками инфекционных заболеваний являются зараженные малярийными плазмодиями люди.
Пути передачи:
- укусы комаров Anopheles,
- процедуры переливания крови, инъекции,
- трансплацентарно — при беременности,
- от больных рожениц к детям.
Малярия отличается сезонным (летне – осенним) характером, что объясняется активизацией размножения комаров.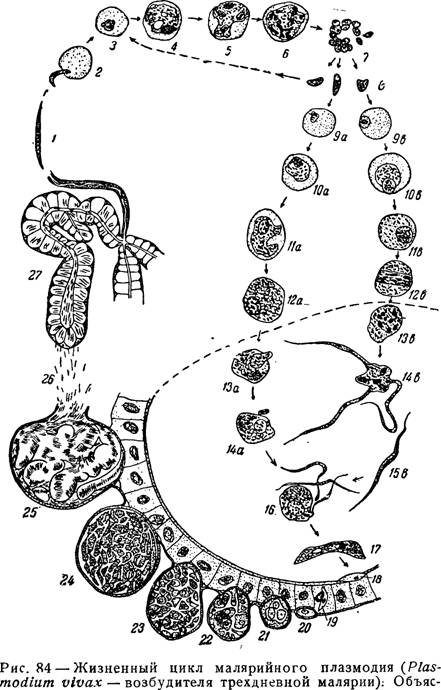
- Бразилия, Вьетнам, Шри-Ланка, Индия, Колумбия, Европа.
- Малярийный плазмодий обитает на побережьях Средиземного моря и Красного.
- А также на Украине, в Таджикистане, Казахстане, Грузии, Азербайджане, Узбекистане.
- Отмечается наличие Зараженных комаров в Белоруссии, России.
Вспышки заболеваний наблюдаются повсюду, за исключением областей Антарктиды, при условии проживания в регионе больных малярией людей и комаров, зараженных плазмодием.
Массовые миграции народа из одной страны в другую, развитие туризма приводят к пандемии (эпидемиям) малярии, поэтому число людей, подвергаемых заболеваемости малярией в мире постоянно возрастает.
CDC — Малярия — О малярии
Anopheles Mosquitoes
Малярия передается человеку самками комаров из рода Anopheles. Самки комаров принимают пищу из крови для производства яиц, и эта пища из крови является связующим звеном между человеком и хозяином комара в жизненном цикле паразита.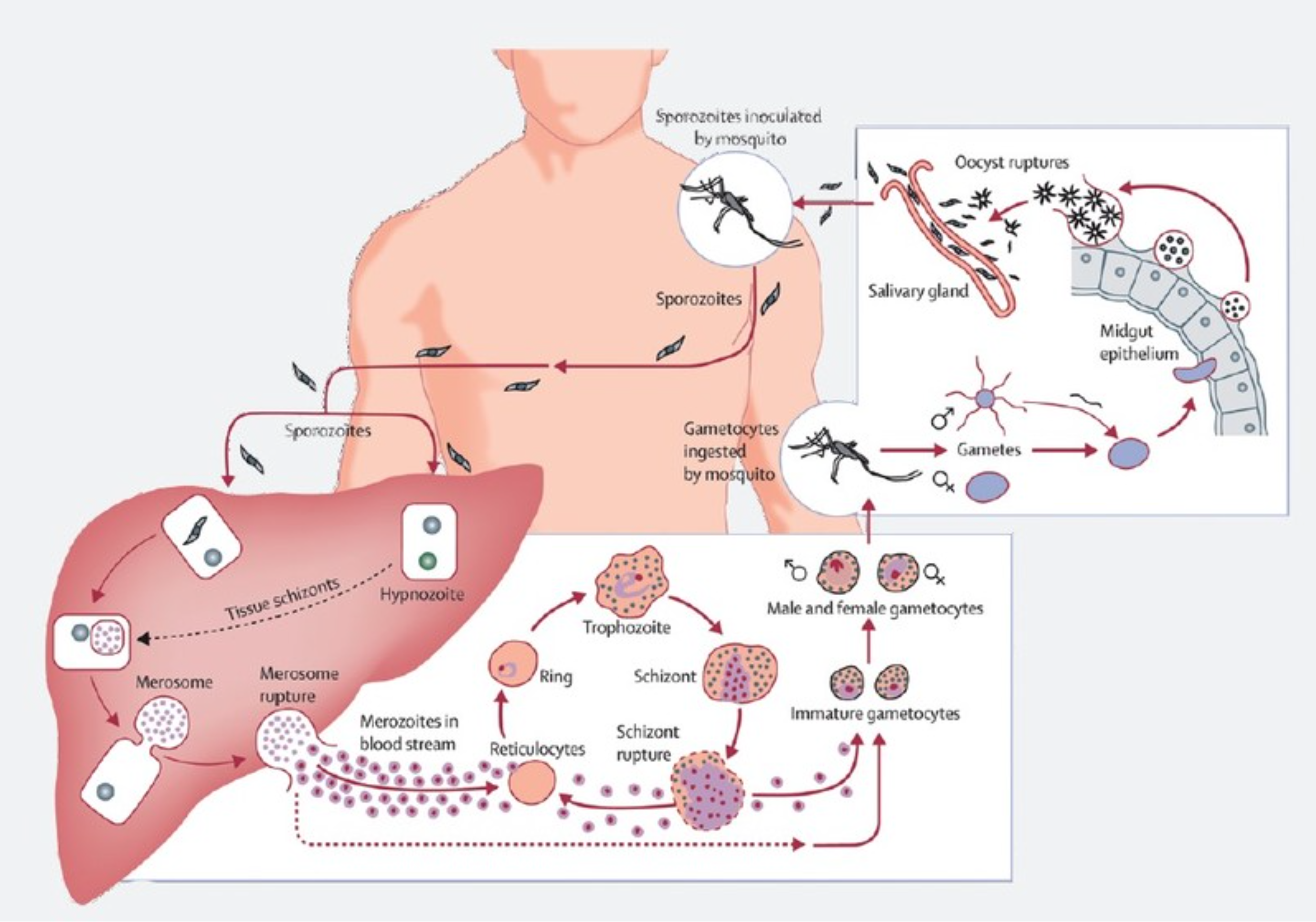 Успешное развитие малярийного паразита в комаре (от стадии «гаметоцит» до стадии «спорозоит») зависит от нескольких факторов.Наиболее важными являются температура и влажность окружающей среды (более высокие температуры ускоряют рост паразита в комаре) и то, выживет ли Anopheles достаточно долго, чтобы позволить паразиту завершить свой цикл в комаре-хозяине («спорогонический» или «внешний» цикл, продолжительность от 9 до 18 дней). В отличие от человека-хозяина, хозяин-комар не страдает от присутствия паразитов.
Успешное развитие малярийного паразита в комаре (от стадии «гаметоцит» до стадии «спорозоит») зависит от нескольких факторов.Наиболее важными являются температура и влажность окружающей среды (более высокие температуры ускоряют рост паразита в комаре) и то, выживет ли Anopheles достаточно долго, чтобы позволить паразиту завершить свой цикл в комаре-хозяине («спорогонический» или «внешний» цикл, продолжительность от 9 до 18 дней). В отличие от человека-хозяина, хозяин-комар не страдает от присутствия паразитов.
Схема взрослых самок комаров
Карта мира, показывающая распространение основных переносчиков малярии
Anopheles freeborni Комар качает кровь
Изображение большего размера
Общая информация
Насчитывается около 3 500 видов комаров, сгруппированных в 41 род.Малярия человека передается только самками рода Anopheles . Из примерно 430 видов Anopheles только 30-40 являются переносчиками малярии (т. Е. Являются «переносчиками») в природе. Остальные либо кусают людей нечасто, либо не могут поддерживать развитие малярийных паразитов.
Е. Являются «переносчиками») в природе. Остальные либо кусают людей нечасто, либо не могут поддерживать развитие малярийных паразитов.
Географическое распространение
Anophelines встречаются по всему миру, кроме Антарктиды. Малярия передается различными видами Anopheles в разных географических регионах. В пределах географических регионов разная среда поддерживает разные виды.
Anophelines, которые могут передавать малярию, обнаружены не только в эндемичных по малярии районах, но также и в районах, где малярия была ликвидирована. Таким образом, эти области подвержены риску повторного занесения болезни.
Жизненные этапы
Как и все комары, комары Anopheles проходят в своем жизненном цикле четыре стадии: яйцо, личинка, куколка и взрослая особь. Первые три этапа — водные и длятся 7-14 дней, в зависимости от вида и температуры окружающей среды. Кусающаяся самка комара Anopheles может переносить малярию.Самцы комаров не кусаются, поэтому не могут передавать малярию или другие заболевания. Взрослые самки, как правило, недолговечны, и лишь небольшая их часть живет достаточно долго (более 10 дней в тропических регионах), чтобы передавать малярию.
Взрослые самки, как правило, недолговечны, и лишь небольшая их часть живет достаточно долго (более 10 дней в тропических регионах), чтобы передавать малярию.
Яйца
Взрослые самки за одну кладку откладывают 50-200 яиц. Яйца откладывают поодиночке прямо на воду и уникальны тем, что имеют поплавки с обеих сторон. Яйца неустойчивы к высыханию и вылупляются в течение 2-3 дней, хотя в более холодном климате вылупление может занять до 2-3 недель.
Личинки
Личинки комаров имеют хорошо развитую голову с щетками для рта, используемыми для кормления, большую грудную клетку и сегментированный живот. У них нет ног. В отличие от других комаров, личинки Anopheles не имеют дыхательного сифона и поэтому располагаются так, чтобы их тело было параллельно поверхности воды.
Верх: Anopheles Яйцо; обратите внимание на боковые поплавки.
Дно: Anopheles яиц откладываются одиночно.
Личинки дышат через дыхальца, расположенные на 8-м сегменте брюшка, и поэтому должны часто выходить на поверхность.
Личинки проводят большую часть своего времени, питаясь водорослями, бактериями и другими микроорганизмами в поверхностном микрослое. Они делают это, поворачивая голову на 180 градусов и питаясь снизу микрослоя. Личинки ныряют под поверхность только при потревожении. Личинки плавают либо резкими движениями всего тела, либо движением щеток для рта.
Личинки проходят 4 стадии развития, после чего превращаются в куколок. В конце каждого возраста личинки линяют, сбрасывая свой экзоскелет или кожу, чтобы обеспечить дальнейший рост.
Anopheles Личинка. Обратите внимание на положение, параллельное поверхности воды.
Личинки встречаются в самых разных средах обитания, но большинство видов предпочитают чистую незагрязненную воду. Личинки комаров Anopheles были обнаружены в пресноводных или соленых болотах, мангровых болотах, рисовых полях, травянистых канавах, краях ручьев и рек, а также в небольших временных водоемах. Многие виды предпочитают места обитания с растительностью. Другие предпочитают среды обитания, которых нет. Некоторые размножаются в открытых, освещенных солнцем бассейнах, в то время как другие встречаются только в затененных местах размножения в лесах. Некоторые виды размножаются в дуплах деревьев или пазухах листьев некоторых растений.
Многие виды предпочитают места обитания с растительностью. Другие предпочитают среды обитания, которых нет. Некоторые размножаются в открытых, освещенных солнцем бассейнах, в то время как другие встречаются только в затененных местах размножения в лесах. Некоторые виды размножаются в дуплах деревьев или пазухах листьев некоторых растений.
Куколка при взгляде сбоку имеет форму запятой. Это переходный этап между личинкой и взрослой особью. Куколка не питается, но претерпевает радикальные метаморфозы. Голова и грудная клетка сливаются в головогруди, а брюшко изгибается снизу.Как и в случае с личинками, куколки должны часто выходить на поверхность, чтобы дышать, что они делают через пару дыхательных труб на головогруди. Через несколько дней в виде куколки спинная поверхность головогруди расщепляется, и взрослый комар выходит на поверхность воды.
Продолжительность от яйца до взрослой особи значительно варьируется у разных видов и сильно зависит от температуры окружающей среды. Комары могут превратиться из яйца в взрослую особь всего за 7 дней, но в тропических условиях обычно это занимает 10-14 дней.
Комары могут превратиться из яйца в взрослую особь всего за 7 дней, но в тропических условиях обычно это занимает 10-14 дней.
Anopheles Куколка
Anopheles Взрослые. Обратите внимание (нижний ряд) на типичное положение покоя.
Взрослые
Как и все комары, взрослые анофелии имеют стройное тело с тремя отделами: головой, грудной клеткой и брюшком.
Голова предназначена для получения сенсорной информации и кормления. На голове расположены глаза и пара длинных, многочлениковых усиков. Усики важны для обнаружения запахов хозяина, а также запахов в местах обитания водных личинок, где самки откладывают яйца.На голове также есть удлиненный, выступающий вперед хоботок, используемый для кормления, и два сенсорных щупика.
Грудная клетка предназначена для передвижения. К грудной клетке прикреплены три пары ног и одна пара крыльев.
Брюшная полость предназначена для переваривания пищи и развития яиц. Эта сегментированная часть тела значительно расширяется, когда самка ест кровь. Кровь со временем переваривается и служит источником белка для производства яиц, которые постепенно заполняют брюшную полость.
Кровь со временем переваривается и служит источником белка для производства яиц, которые постепенно заполняют брюшную полость.
Комаров Anopheles можно отличить от других комаров по пальпам, длина которых равна длине хоботка, и по наличию отдельных блоков черной и белой чешуек на крыльях. Взрослых особей Anopheles можно также идентифицировать по их типичному положению покоя: самцы и самки отдыхают так, что их брюшко торчит в воздух, а не параллельно поверхности, на которой они отдыхают .
Взрослые комары обычно спариваются в течение нескольких дней после выхода из стадии куколки.У некоторых видов самцы образуют большие стаи, обычно в сумерках, а самки летают в стаи для спаривания. Места спаривания многих видов остаются неизвестными.
Самцы живут около недели, питаясь нектаром и другими источниками сахара. Самки также питаются источниками сахара для получения энергии, но обычно им требуется кровяная мука для развития яиц. После полноценной еды самка будет отдыхать в течение нескольких дней, пока кровь переваривается и развиваются яйца. Этот процесс зависит от температуры, но в тропических условиях обычно занимает 2-3 дня.Когда яйца полностью развиваются, самка откладывает их, а затем ищет кровь, чтобы выдержать еще одну партию яиц.
Этот процесс зависит от температуры, но в тропических условиях обычно занимает 2-3 дня.Когда яйца полностью развиваются, самка откладывает их, а затем ищет кровь, чтобы выдержать еще одну партию яиц.
Цикл повторяется до самой смерти самки. Самки могут прожить до месяца (или дольше в неволе), но большинство из них не живут дольше 1-2 недель в природе. Их шансы на выживание зависят от температуры и влажности, а также от их способности успешно питаться кровью, избегая при этом защиты хозяина.
Самка Anopheles dirus кормление
Факторы, участвующие в передаче малярии и борьбе с ней
Понимание биологии и поведения комаров Anopheles может помочь в разработке соответствующих стратегий борьбы.Факторы, которые влияют на способность комара переносить малярию, включают его врожденную восприимчивость к Plasmodium , выбор хозяина и продолжительность жизни. Наиболее опасны долгожители, предпочитающие человеческую кровь и поддерживающие развитие паразитов.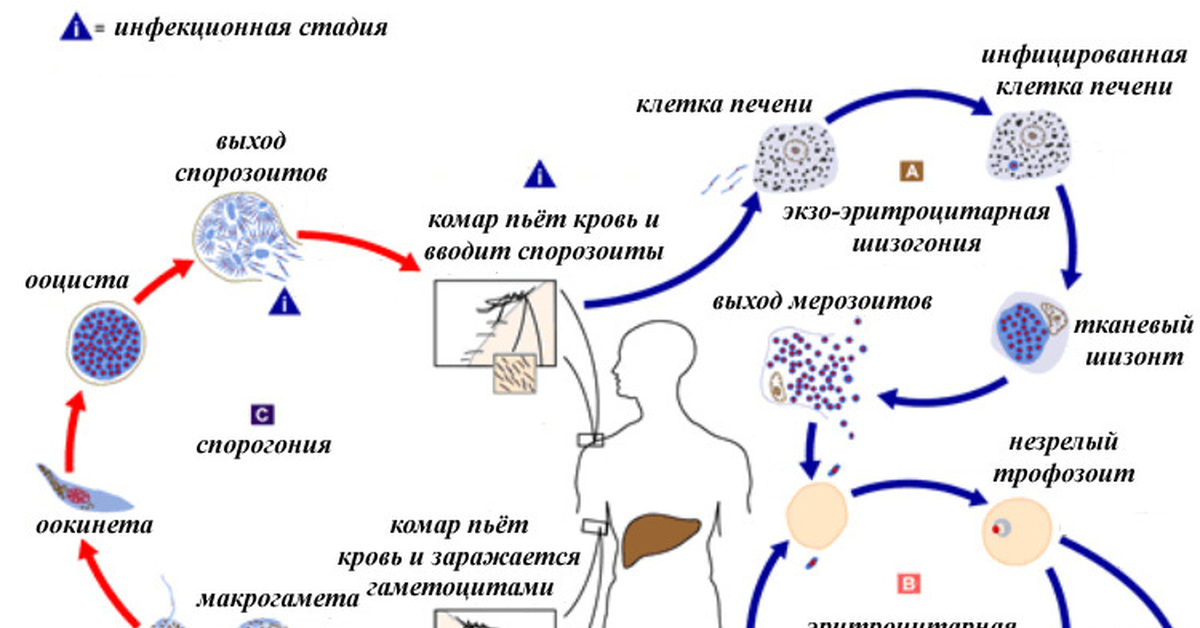 Факторы, которые следует учитывать при разработке программы борьбы, включают восприимчивость малярийных комаров к инсектицидам и предпочтительные места кормления и отдыха взрослых комаров.
Факторы, которые следует учитывать при разработке программы борьбы, включают восприимчивость малярийных комаров к инсектицидам и предпочтительные места кормления и отдыха взрослых комаров.
Подробнее: как уменьшить воздействие малярии
Предпочтительные источники крови
Одним из важных поведенческих факторов является степень, в которой вид Anopheles предпочитает питаться людьми (антропофилия) или животными, такими как крупный рогатый скот (зоофилия).Anthrophilic Anopheles с большей вероятностью передаст малярийных паразитов от одного человека к другому. Большинство комаров Anopheles не являются исключительно антропофильными или зоофильными; многие из них приспособлены и питаются любым доступным хостом. Однако основные переносчики малярии в Африке, An. gambiae и An. funestus , являются сильно антропофильными и, следовательно, являются двумя наиболее эффективными переносчиками малярии в мире.
Срок службы
Попадая в организм комара, малярийные паразиты должны развиться внутри комара, прежде чем они заразятся для человека. Время, необходимое для развития у комара (внешний инкубационный период), составляет 9 дней или больше, в зависимости от вида паразита и температуры. Если комар не проживет дольше внешнего инкубационного периода, он не сможет передавать малярийных паразитов.
Время, необходимое для развития у комара (внешний инкубационный период), составляет 9 дней или больше, в зависимости от вида паразита и температуры. Если комар не проживет дольше внешнего инкубационного периода, он не сможет передавать малярийных паразитов.
Непосредственно измерить продолжительность жизни комаров в природе невозможно, но во многих исследованиях продолжительность жизни косвенно измерялась путем изучения их репродуктивного статуса или путем маркировки, выпуска и повторного отлова взрослых комаров.Большинство комаров не живут достаточно долго, чтобы передавать малярию, но некоторые могут жить до трех недель в природе. Хотя данные свидетельствуют о том, что уровень смертности увеличивается с возрастом, большинство рабочих оценивают долголетие с точки зрения вероятности того, что комар однажды проживет. Обычно эти оценки варьируются от 0,7 до 0,9. Если выживаемость составляет 90% ежедневно, то значительная часть населения будет жить дольше 2 недель и будет способна передавать малярию.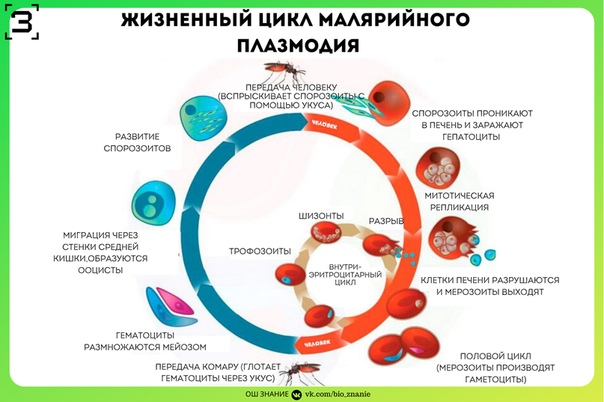 Любая мера контроля, сокращающая среднюю продолжительность жизни популяции комаров, снизит потенциал передачи. Таким образом, инсектициды не обязательно убивают комаров сразу, но могут быть эффективными, ограничивая их продолжительность жизни.
Любая мера контроля, сокращающая среднюю продолжительность жизни популяции комаров, снизит потенциал передачи. Таким образом, инсектициды не обязательно убивают комаров сразу, но могут быть эффективными, ограничивая их продолжительность жизни.
Способы кормления и отдыха
Большинство комаров Anopheles ведут сумеречный образ жизни (активны в сумерках или на рассвете) или ведут ночной образ жизни (активны ночью). Некоторые комары Anopheles питаются в помещении (эндофаги), а другие — на открытом воздухе (экзофаги).После кормления кровью около комаров Anopheles предпочитают отдыхать в помещении (эндофильный), в то время как другие предпочитают отдых на открытом воздухе (экзофильный). Укусы ночных, эндофагических комаров Anopheles можно значительно уменьшить за счет использования обработанных инсектицидами надкроватных сеток (ITN) или за счет улучшенной конструкции жилья для предотвращения проникновения комаров (например, оконных сеток).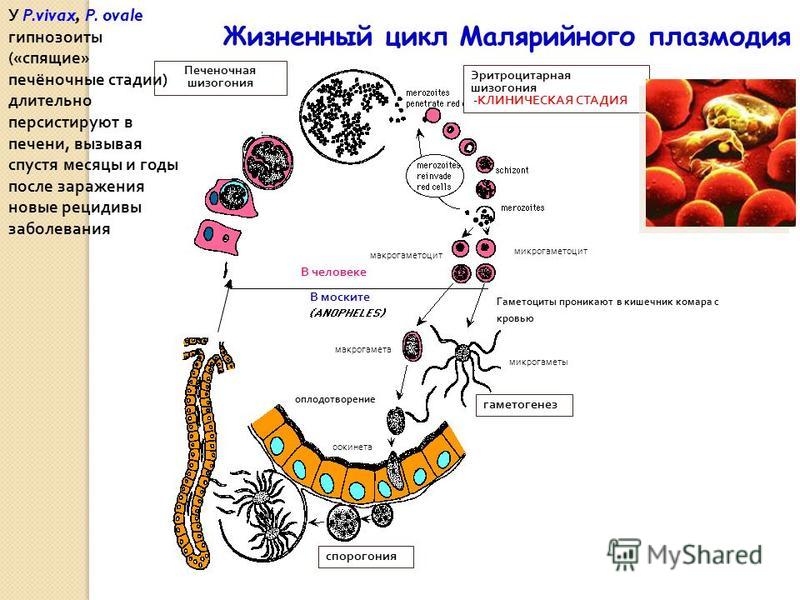 С эндофильными комарами легко бороться с помощью распыления в помещении остаточных инсектицидов. Напротив, экзофагические / экзофильные переносчики лучше всего контролируются путем сокращения источников (разрушения местообитаний личинок).
С эндофильными комарами легко бороться с помощью распыления в помещении остаточных инсектицидов. Напротив, экзофагические / экзофильные переносчики лучше всего контролируются путем сокращения источников (разрушения местообитаний личинок).
Устойчивость к инсектицидам
Меры контроля на основе инсектицидов (например, опрыскивание помещений инсектицидами, ОИС) являются основным способом уничтожения комаров, которые кусаются в помещении. Однако после длительного воздействия инсектицида в течение нескольких поколений у комаров, как и у других насекомых, может развиться устойчивость, способность выживать при контакте с инсектицидом. Поскольку комары могут иметь много поколений в год, высокий уровень устойчивости может возникнуть очень быстро. Устойчивость комаров к некоторым инсектицидам была подтверждена в течение нескольких лет после их внедрения.Существует более 125 видов комаров с подтвержденной устойчивостью к одному или нескольким инсектицидам. Развитие устойчивости к инсектицидам, используемым для остаточного опрыскивания помещений, было основным препятствием во время Глобальной кампании по искоренению малярии.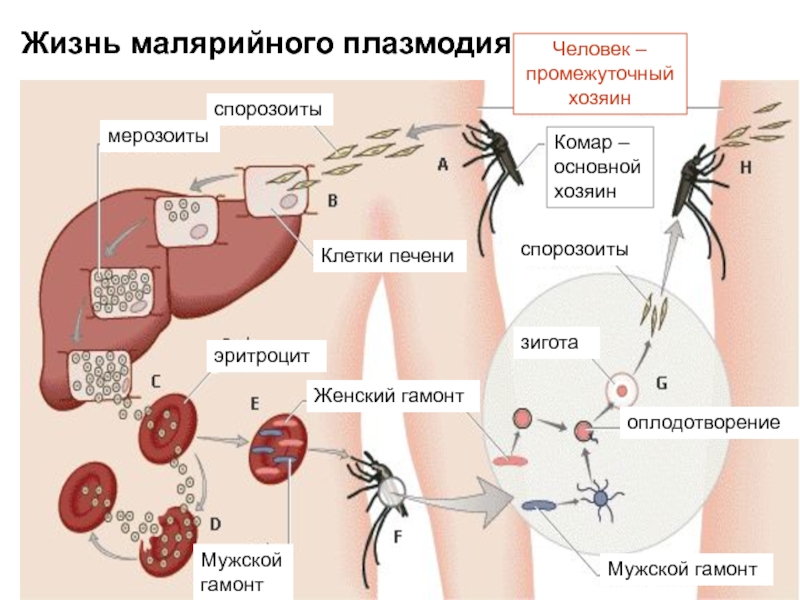 Разумное использование инсектицидов для борьбы с комарами может ограничить развитие и распространение устойчивости, особенно за счет ротации различных классов инсектицидов, используемых для борьбы. Мониторинг устойчивости важен для предупреждения программ контроля о переходе на более эффективные инсектициды.
Разумное использование инсектицидов для борьбы с комарами может ограничить развитие и распространение устойчивости, особенно за счет ротации различных классов инсектицидов, используемых для борьбы. Мониторинг устойчивости важен для предупреждения программ контроля о переходе на более эффективные инсектициды.
Восприимчивость / рефрактерность
Около видов Anopheles являются плохими переносчиками малярии, так как паразиты не развиваются (или не развиваются вообще) внутри них. Есть также вариации внутри видов. В лаборатории удалось отобрать штаммы An. gambiae , устойчивые к заражению малярийными паразитами. Эти невосприимчивые штаммы обладают иммунным ответом, который инкапсулирует и убивает паразитов после того, как они проникли в стенку желудка комара.Ученые изучают генетический механизм этой реакции. Есть надежда, что когда-нибудь генетически модифицированные комары, невосприимчивые к малярии, смогут заменить диких комаров, тем самым ограничив или исключив передачу малярии.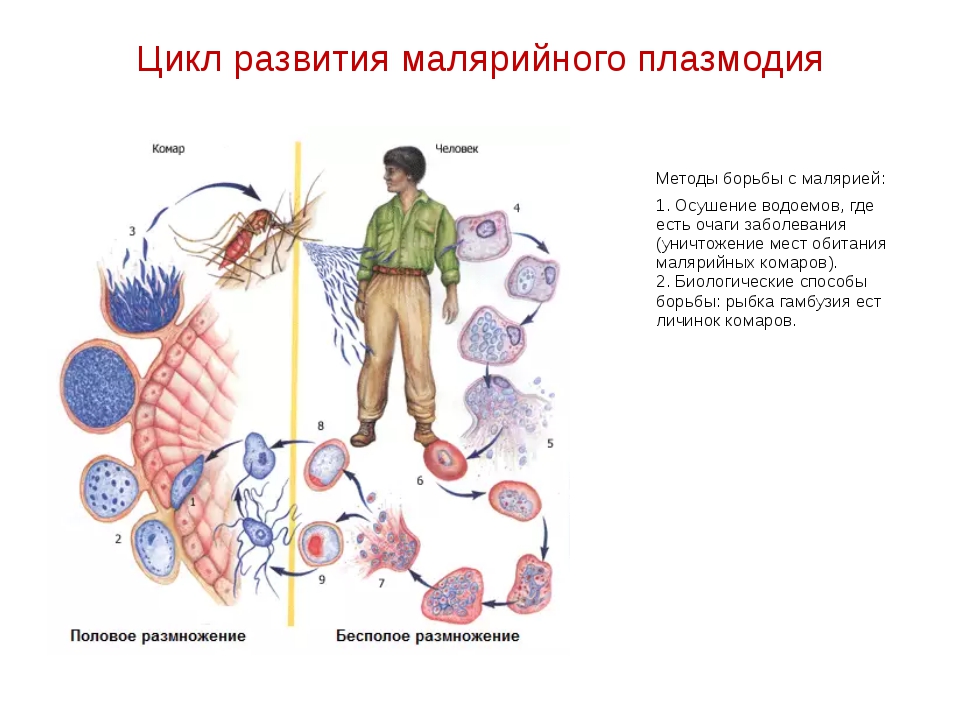
Малярийный паразит
жизненный цикл включает два хозяина. Во время кровопролития зараженный малярией
самка Anopheles комар инокулирует спорозоиты в организме человека-хозяина
.
Спорозоиты поражают клетки печени
и зрелый
в шизонтов
, который
разрыв и высвобождение мерозоитов
. (Из
обратите внимание, в P. vivax и P. ovale находится стадия покоя [гипнозоиты]
может сохраняться в печени и вызывать рецидивы, вторгаясь в кровоток
недели или даже годы спустя.) После этой первоначальной репликации в печени (экзоэритроцитарная
шизогония
),
паразиты подвергаются бесполому размножению в эритроцитах (эритроцитарные
шизогония
).
Мерозоиты заражают эритроциты
. Кольцо
стадия трофозоитов созревает в шизонты, которые разрывают высвобождение мерозоитов
. Немного
паразиты дифференцируются на половые эритроцитарные стадии (гаметоциты)
. Кровь
стадийные паразиты ответственны за клинические проявления
болезнь. Кровь
стадийные паразиты ответственны за клинические проявления
болезнь.Мужские (микрогаметоциты) и женские (макрогаметоциты) гаметоциты проглочен комаром Anopheles во время еды с кровью .В размножение паразитов у комара известно как спорогонический цикл. . Находясь в желудке комара, микрогаметы проникают в макрогаметы. создание зигот . В зиготы в свою очередь становятся подвижными и удлиненными (оокинеты) который вторгаются в стенку средней кишки комара, где они развиваются в ооцисты . В ооцисты растут, разрываются и выделяют спорозоиты , который пробиваются к слюнным железам комара.Прививка спорозоитов в нового человека-хозяина увековечивает жизненный цикл малярии . |
Бесполый рост и половое развитие плазмодиев в гематопоэтической нише хозяина
ВОЗ. Всемирный доклад о малярии, 2018 г. . https://www.who.int/malaria/publications/world-malaria-report-2018/report/en/ (2018).
Tavares, J. et al. Роль пересечения клетки-хозяина спорозоитом малярии во время инфекции печени. J. Exp. Med. 210 , 905–915 (2013).
CAS PubMed PubMed Central Google ученый
Amino, R. et al. Количественная визуализация передачи Plasmodium от комара к млекопитающему. Nat. Med. 12 , 220–224 (2006).
CAS PubMed Google ученый
Mota, M. M. et al. Миграция спорозоитов Plasmodium через клетки до заражения.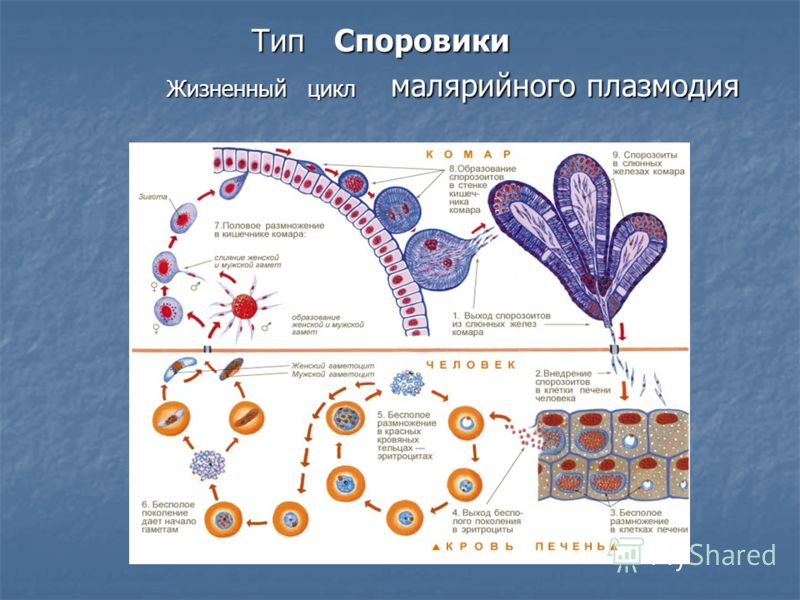 Наука 291 , 141–144 (2001).
Наука 291 , 141–144 (2001).
CAS PubMed Google ученый
Sturm, A. et al. Манипулирование гепатоцитами хозяина малярийным паразитом для доставки в синусоиды печени. Наука 313 , 1287–1290 (2006).
CAS PubMed Google ученый
Сологуб Л. и др. Протеазы малярии опосредуют выход гаметоцитов из эритроцитов наизнанку после передачи паразита комару. Cell Microbiol. 13 , 897–912 (2011).
CAS PubMed Google ученый
Янсе, К. Дж., Рамесар, Дж., Ван ден Берг, Ф. М. и Монс, Б. Plasmodium berghei : создание и отбор in vivo мутантов кариотипа и мутантов-продуцентов негаметоцитов. Exp. Паразитол. 74 , 1–10 (1992).
Plasmodium berghei : создание и отбор in vivo мутантов кариотипа и мутантов-продуцентов негаметоцитов. Exp. Паразитол. 74 , 1–10 (1992).
CAS PubMed Google ученый
Alano, P. et al. Plasmodium falciparum : паразиты, дефектные на ранних стадиях гаметоцитогенеза. Exp. Паразитол. 81 , 227–235 (1995).
CAS PubMed Google ученый
Kafsack, B. F. et al. Переключение транскрипции лежит в основе приверженности половому развитию малярийных паразитов. Природа 507 , 248–252 (2014).
CAS PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 10.
org/ScholarlyArticle»> 10.Sinha, A. et al. Каскад ДНК-связывающих белков для сексуальной ориентации и развития у Plasmodium . Природа 507 , 253–257 (2014).
CAS PubMed PubMed Central Google ученый
Brancucci, N.M.B. et al. Гетерохроматиновый белок 1 обеспечивает выживание и передачу малярийных паразитов. Клеточный микроб-хозяин 16 , 165–176 (2014).
CAS PubMed Google ученый
Coleman, B. I. et al. Гистондеацетилаза Plasmodium falciparum регулирует антигенную изменчивость и конверсию гаметоцитов. Клеточный микроб-хозяин 16 , 177–186 (2014).
CAS PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 13.
org/ScholarlyArticle»> 13.Eksi, S. et al. Plasmodium falciparum Развитие гаметоцитов 1 (Pfgdv1) и идентификация ранних генов гаметоцитогенеза и приверженность половому развитию. PLOS Pathog. 8 , e1002964 (2012).
CAS PubMed PubMed Central Google ученый
Filarsky, M. et al. GDV1 вызывает половую активность малярийных паразитов, противодействуя HP1-зависимому подавлению генов. Наука 359 , 1259–1263 (2018).
CAS PubMed PubMed Central Google ученый
Баклинг, А., Ранфорд-Картрайт, Л.С., Майлз, А. и Рид, А.Ф. Хлорохин усиливает гаметоцитогенез Plasmodium falciparum in vitro. Паразитология 118 (Pt 4), 339–346 (1999).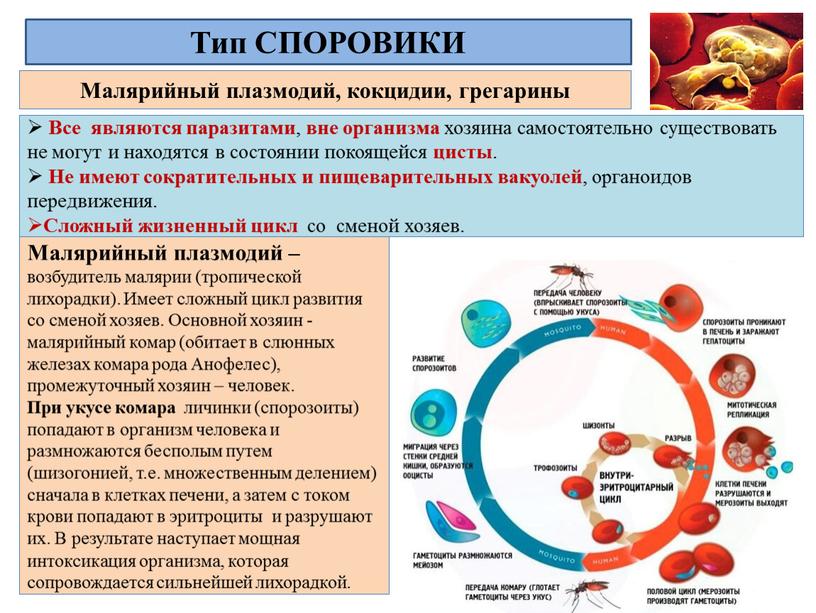
CAS PubMed Google ученый
Buchholz, K. et al. Высокопроизводительный скрининг, нацеленный на стадии передачи малярии, открывает новые возможности для разработки лекарств. J. Infect. Дис. 203 , 1445–1453 (2011).
PubMed PubMed Central Google ученый
Brancucci, N.M.B. et al. Лизофосфатидилхолин регулирует дифференциацию половых стадий у малярийного паразита человека Plasmodium falciparum . Ячейка 171 , 1532–1544.e15 (2017).
CAS PubMed PubMed Central Google ученый
Бранкучи, Н. М.B. et al. Исследование Plasmodium falciparum сексуальной активности на одноклеточном уровне. Wellcome Open Res. 3 , 70 (2018).
PubMed PubMed Central Google ученый
Poran, A. et al. Секвенирование одноклеточной РНК выявляет признак сексуальной приверженности малярийных паразитов. Nature 551 , 95–99 (2017).
PubMed PubMed Central Google ученый
Brancucci, N.M., Witmer, K., Schmid, C. & Voss, T. S. Верхний элемент гена var контролирует синтез белка на уровне инициации трансляции в Plasmodium falciparum . PLOS ONE 9 , e100183 (2014).
PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 21.
org/ScholarlyArticle»> 21.Джослинг, Г. А. и Ллинас, М. Половое развитие у паразитов Plasmodium : знание, когда пора совершать действия. Nat. Rev. Microbiol. 13 , 573–587 (2015).
CAS PubMed Google ученый
Джослинг Г. А., Уильямсон К. К. и Ллинас М. Регулирование полового влечения и гаметоцитогенеза у малярийных паразитов. Annu. Rev. Microbiol. 72 , 501–519 (2018).
CAS PubMed Google ученый
Хокинг, Ф., Wilson, M. E. & Gammage, K. Доказательства циклического развития и недолгой зрелости гаметоцитов Plasmodium falciparum . Пер. R. Soc. Троп. Med. Hyg. 65 , 549–559 (1971).
CAS PubMed Google ученый
 org/ScholarlyArticle»> 24.
org/ScholarlyArticle»> 24.Gautret, P. & Motard, A. Периодическая инфекционность гаметоцитов Plasmodium по отношению к вектору. Обзор. Паразиты 6 , 103–111 (1999).
CAS PubMed Google ученый
Dearnley, M. et al. Обратимое ремоделирование клеток-хозяев лежит в основе изменений деформируемости на стадиях половой крови малярийных паразитов. Proc. Natl Acad. Sci. США 113 , 4800–4805 (2016).
CAS PubMed Google ученый
Hliscs, M. et al. Организация и функция актинового цитоскелета в гаметоцитах Plasmodium falciparum . Cell. Microbiol. 17 , 207–225 (2015).
CAS PubMed Google ученый
 org/ScholarlyArticle»> 27.
org/ScholarlyArticle»> 27.Dearnley, M. K. et al. Происхождение, состав, организация и функция комплекса внутренней мембраны гаметоцитов Plasmodium falciparum . J. Cell Sci. 125 , 2053–2063 (2012).
CAS PubMed Google ученый
Turner, L. et al. Тяжелая форма малярии связана со связыванием паразитов с рецептором эндотелиального протеина С. Природа 498 , 502–505 (2013).
CAS PubMed PubMed Central Google ученый
Seydel, K. B. et al. Набухание мозга и смерть у детей от церебральной малярии. N. Engl. J. Med. 372 , 1126–1137 (2015).
CAS PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 30.
org/ScholarlyArticle»> 30.Фрид М. и Даффи П. Э. Привязка Plasmodium falciparum к хондроитинсульфату а в плаценте человека. Наука 272 , 1502–1504 (1996).
CAS PubMed Google ученый
Cranston, H.A. et al. Plasmodium falciparum Созревание устраняет физиологическую деформируемость эритроцитов. Наука 223 , 400–403 (1984).
CAS PubMed Google ученый
Гинзбург, Х. и Кругляк, М. Поглощение L-триптофана эритроцитами, инфицированными малярийными паразитами ( Plasmodium falciparum ). Biochim. Биофиз. Acta 729 , 97–103 (1983).
CAS PubMed Google ученый
 org/ScholarlyArticle»> 33.
org/ScholarlyArticle»> 33.Вествебер Д. Как лейкоциты проникают через эндотелий сосудов. Nat. Rev. Immunol. 15 , 692–704 (2015).
CAS PubMed Google ученый
Пиявка, Дж. Х., Барнуэлл, Дж. У., Миллер, Л. Х. и Ховард, Р. Дж. Идентификация штамм-специфического малярийного антигена, экспонированного на поверхности эритроцитов, инфицированных Plasmodium falciparum . J. Exp. Med. 159 , 1567–1575 (1984).
CAS PubMed Google ученый
Su, X. Z. et al. Большое разнообразие генов семейства var кодирует белки, участвующие в цитоадгезии и антигенной вариации эритроцитов, инфицированных Plasmodium falciparum . Cell 82 , 89–100 (1995).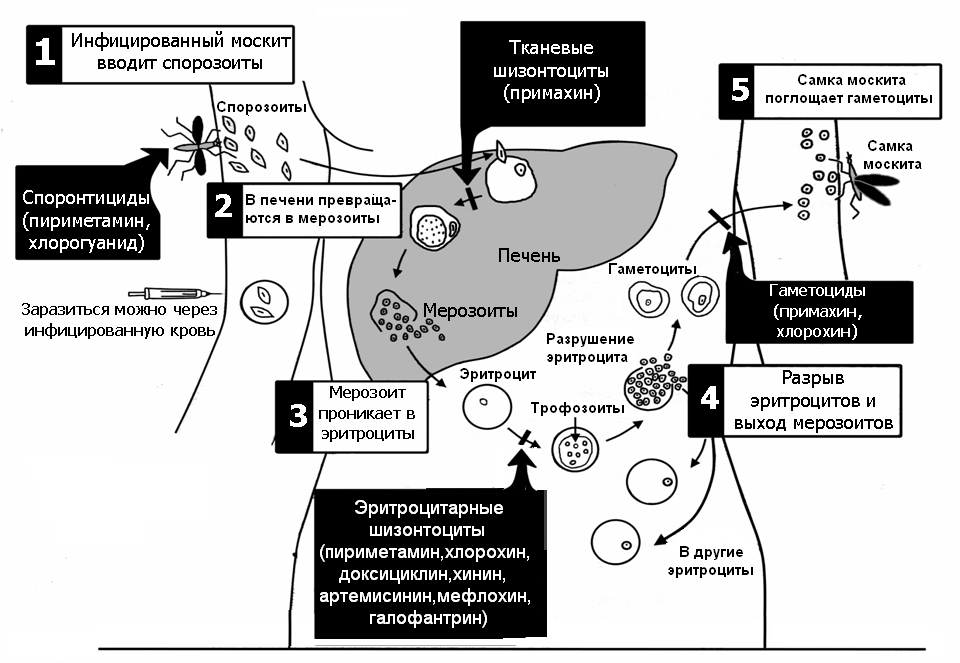
CAS PubMed Google ученый
Smith, J. D. et al. Идентификация связывающего домена молекулы-1 межклеточной адгезии Plasmodium falciparum : признак адгезии паразитов, связанный с церебральной малярией. Proc. Natl Acad. Sci. США 97 , 1766–1771 (2000).
CAS PubMed Google ученый
Salanti, A. et al. Доказательства участия VAR2CSA в связанной с беременностью малярии. J. Exp. Med. 200 , 1197–1203 (2004).
CAS PubMed PubMed Central Google ученый
Flick, K. et al. Роль неиммунного IgG, связанного с PfEMP1, при плацентарной малярии. Наука 293 , 2098–2100 (2001).
Наука 293 , 2098–2100 (2001).
CAS PubMed Google ученый
Chan, J. A. et al. Мишени антител против эритроцитов, инфицированных Plasmodium falciparum , при иммунитете к малярии. J. Clin. Вкладывать деньги. 122 , 3227–3238 (2012).
CAS PubMed PubMed Central Google ученый
Kyes, S. A., Rowe, J. A., Kriek, N. & Newbold, C. I. Rifins: второе семейство клонально вариантных белков, экспрессируемых на поверхности эритроцитов, инфицированных Plasmodium falciparum . Proc. Natl Acad. Sci. США 96 , 9333–9338 (1999).
CAS PubMed Google ученый
 org/ScholarlyArticle»> 41.
org/ScholarlyArticle»> 41.Cheng, Q. et al. Stevor и Rif — это мультикопийные семейства генов Plasmodium falciparum , которые потенциально кодируют вариантные антигены. Мол. Biochem. Паразитол. 97 , 161–176 (1998).
CAS PubMed Google ученый
Niang, M. et al. STEVOR — это связывающий эритроциты белок Plasmodium falciparum , который опосредует инвазию и розетку мерозоитов. Клеточный микроб-хозяин 16 , 81–93 (2014).
CAS PubMed PubMed Central Google ученый
Goel, S. et al. RIFIN — это адгезины, вызывающие тяжелую малярию Plasmodium falciparum . Nat. Med. 21 , 314–317 (2015).
CAS PubMed Google ученый
 org/ScholarlyArticle»> 44.
org/ScholarlyArticle»> 44.Смит, К. Д., Браун, А. Е., Накадзава, С., Фудзиока, Х. и Айкава, М. Секвестрация мультиорганных эритроцитов и экспрессия лиганда у макак-резусов, инфицированных малярией Plasmodiumcoatneyi . Am. J. Trop. Med. Hyg. 55 , 379–383 (1996).
CAS PubMed Google ученый
Handayani, S. et al. Высокая деформируемость эритроцитов, инфицированных Plasmodium vivax , в микрофлюидных условиях. J. Infect. Дис. 199 , 445–450 (2009).
PubMed PubMed Central Google ученый
Marchiafava, E. & Bignami, A. Sulle febbri estivo aumnali ( E. Loescher , 1894).
 org/ScholarlyArticle»> 47.
org/ScholarlyArticle»> 47.Baro, B. et al. Plasmodium vivax гаметоцитов в костном мозге больного острой малярией и изменения в профиле миРНК эритроидных клеток. PLOS Negl. Троп. Дис. 11 , e0005365 (2017).
PubMed PubMed Central Google ученый
Фарфур, Э., Шарлотта, Ф., Сеттеграна, К., Мияра, М. и Баффет, П. Внесосудистый отсек костного мозга: ниша для созревания гаметоцитов Plasmodium falciparum ? Малар. J. 11 , 285 (2012).
PubMed PubMed Central Google ученый
Смолли М. Э., Абдалла С. и Браун Дж. Распределение Plasmodium falciparum в периферической крови и костном мозге детей Гамбии. Пер. R. Soc. Троп.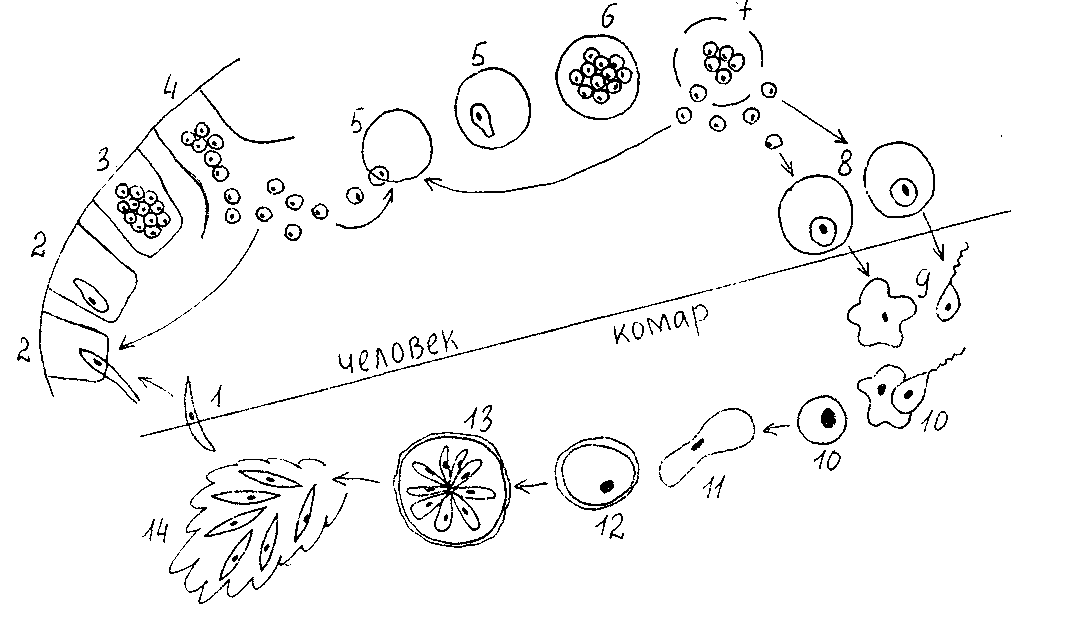 Med. Hyg. 75 , 103–105 (1981).
Med. Hyg. 75 , 103–105 (1981).
CAS PubMed Google ученый
Томсон, Дж. Г. и Робертсон, А. Структура и развитие гаметоцитов Plasmodium falciparum во внутренних органах и периферическом кровообращении. Пер. R. Soc. Троп. Med. Hyg. 14 , 31–40 (1935).
Google ученый
Joice, R. et al. Plasmodium falcipa Этапы передачи рома накапливаются в костном мозге человека. Sci. Пер. Med. 6 , 244re245 (2014). Это исследование предоставляет количественные доказательства P. falciparum обогащения гаметоцитов в костном мозге человека .
Google ученый
 org/ScholarlyArticle»> 52.
org/ScholarlyArticle»> 52.Aguilar, R. et al. Молекулярные доказательства локализации незрелых гаметоцитов Plasmodium falciparum в костном мозге. Кровь 123 , 959–966 (2014).
CAS PubMed PubMed Central Google ученый
Эллис, Р. Э. Распределение активного костного мозга у взрослых. Phys.Med. Биол. 5 , 255–258 (1961).
CAS PubMed Google ученый
Woodard, H. Q. & Holodny, E. Обобщение данных Mechanik о распределении костного мозга человека. Phys. Med. Биол. 5 , 57–59 (1960).
CAS PubMed Google ученый
 org/ScholarlyArticle»> 55.
org/ScholarlyArticle»> 55.Номбела-Арриета, К. и Манц, М. Г. Количественная оценка и трехмерная микроанатомическая организация костного мозга. Blood Adv. 1 , 407–416 (2017).
CAS PubMed PubMed Central Google ученый
Часис, Дж. А. и Мохандас, Н. Эритробластические острова: ниши для эритропоэза. Кровь 112 , 470–478 (2008).
CAS PubMed PubMed Central Google ученый
Obaldia, N., 3rd et al. Костный мозг является основным резервуаром паразитов для инфекции Plasmodium vivax . MBio 8 , 1–16 (2018). Это исследование предоставляет количественные доказательства P. vivax обогащения гаметоцитами и бесполого резервуара в костном мозге нечеловеческих приматов .
Google ученый
De Niz, M. et al. Plasmodium гаметоцитов обнаруживают самонаведение и трансмиграцию сосудов в костный мозг хозяина. Sci. Adv. 4 , eaat3775 (2018). Это исследование демонстрирует P. berghei бесполый резервуар и резервуар гаметоцитов в костном мозге и селезенке инфицированных мышей и дает первое описание событий трансмиграции паразитов через эндотелиальный барьер .
PubMed PubMed Central Google ученый
Duffier, Y. et al. Гуманизированная модель мыши для секвестрации половых стадий Plasmodium falciparum и оценки гаметоцитидных препаратов in vivo. Sci. Отчет 6 , 35025 (2016). Это исследование демонстрирует P. faciparum обогащение гаметоцитов в костном мозге и селезенке гуманизированных мышей .
Это исследование демонстрирует P. faciparum обогащение гаметоцитов в костном мозге и селезенке гуманизированных мышей .
CAS PubMed PubMed Central Google ученый
Ли, Р. С., Уотерс, А. П. и Брюер, Дж. М. Скрытый цикл в гематопоэтических нишах способствует инициации передачи малярии и уклонению от химиотерапии. Nat. Commun. 9 , 1689 (2018). Это исследование предоставляет доказательства того, что бесполый P. berghei резервуар паразитов в селезенке инфицированных мышей может привести к рецидиву .
PubMed PubMed Central Google ученый
Lopes, S.C. et al. Недостаточность зрелых шизонтов Plasmodium vivax в периферической крови связана с их повышенным цитоадгезивным потенциалом.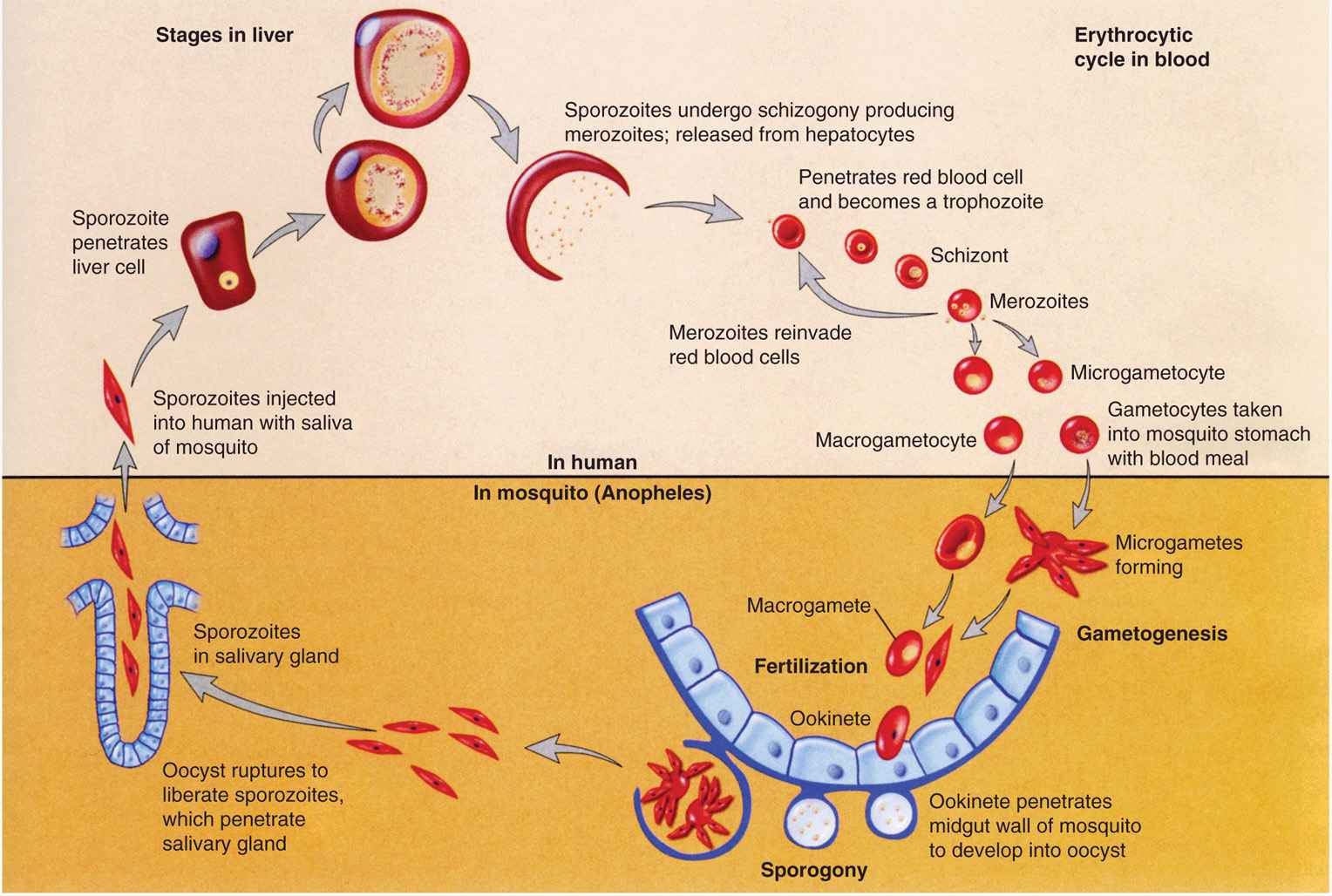 J. Infect. Дис. 209 , 1403–1407 (2014).
J. Infect. Дис. 209 , 1403–1407 (2014).
PubMed Google ученый
Lim, C. et al. Предпочтение ретикулоцитов и стадия развития изолятов Plasmodium vivax . J. Infect. Дис. 214 , 1081–1084 (2016).
PubMed PubMed Central Google ученый
Fonseca, L. L., Joyner, C.Дж., Ма, Х. С., Галински, М. Р. и Войт, Е. О. Модель маскировки Plasmodium vivax на основе инфекции Plasmodium cynomolgi в Macaca mulatta . Малар. Дж. 16 , 375 (2017).
PubMed PubMed Central Google ученый
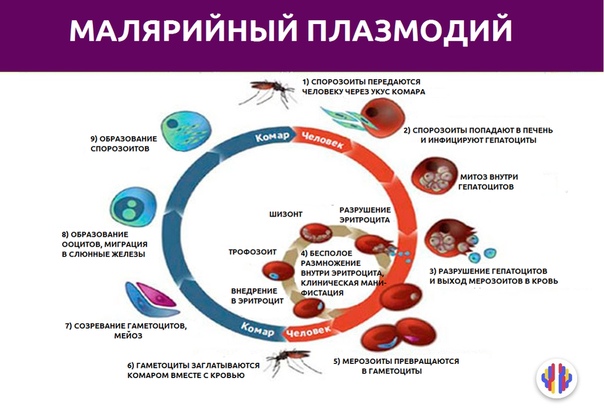 org/ScholarlyArticle»> 64.
org/ScholarlyArticle»> 64.Usui, M. et al. Plasmodium falciparum Половая дифференциация у больных малярией связана с факторами хозяина и генами, зависимыми от GDV1. Nat. Commun. 10 , 2140 (2019).
PubMed PubMed Central Google ученый
Фарид Р., Диксон М. В., Тилли Л. и Маккарти Дж. С. Инициирование гаметоцитогенеза при очень низкой плотности паразитов при инфекции Plasmodium falciparum . J. Infect. Дис. 215 , 1167–1174 (2017).
CAS PubMed PubMed Central Google ученый
Rono, M. K. et al. Адаптация Plasmodium falciparum к среде его передачи. Nat. Ecol. Evol. 2 , 377–387 (2018).
PubMed Google ученый
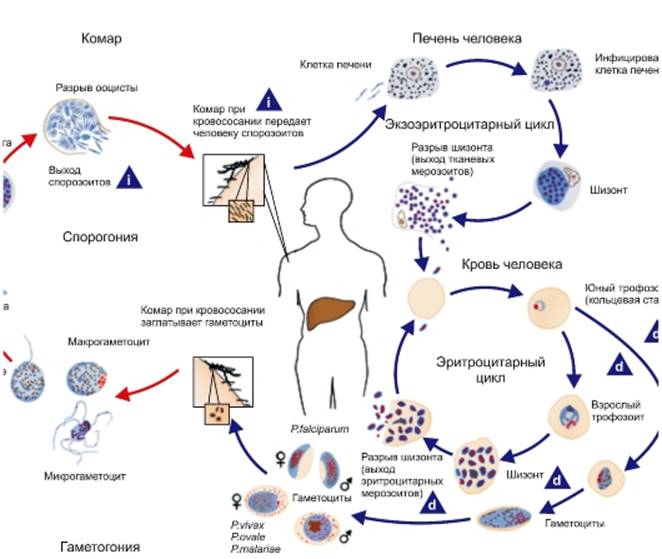 org/ScholarlyArticle»> 67.
org/ScholarlyArticle»> 67.Pelle, K. G. et al. Транскрипционное профилирование определяет динамику секвестрации тканей паразита при заражении малярией. Genome Med. 7 , 19 (2015).
PubMed PubMed Central Google ученый
Perrin, A.J., Bartholdson, S.J. и Wright, G.J. P-селектин является рецептором хозяина для лигандов Plasmodium MSP7. Малар. J. 14 , 238 (2015).
PubMed PubMed Central Google ученый
Avril, M., Brazier, A.J., Melcher, M., Sampath, S. & Smith, J. D. Гены DC8 и DC13 var, связанные с тяжелой формой малярии, активно связываются с различными эндотелиальными клетками. PLOS Pathog. 9 , e1003430 (2013).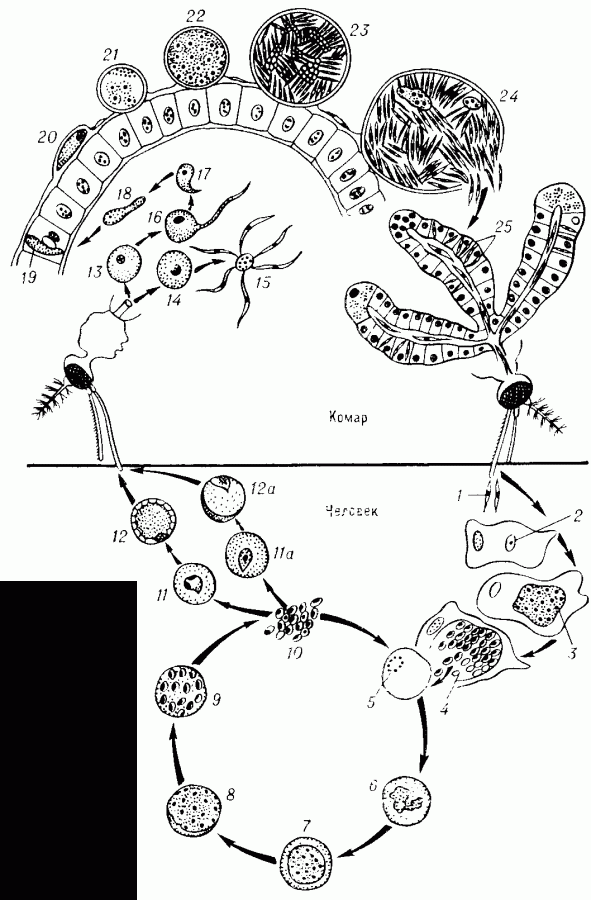
CAS PubMed PubMed Central Google ученый
Rogers, NJ, Hall, BS, Obiero, J., Targett, GA & Sutherland, CJ Модель секвестрации стадий передачи Plasmodium falciparum : адгезия эритроцитов, инфицированных гаметоцитами, к костному мозгу человека клетки. Заражение. Иммун. 68 , 3455–3462 (2000).
CAS PubMed PubMed Central Google ученый
Tiburcio, M. et al. Ранние гаметоциты малярийного паразита Plasmodium falciparum специфически изменяют адгезионные свойства инфицированной поверхности эритроцитов. Cell. Microb. 15 , 647–659 (2012).
Google ученый
 org/ScholarlyArticle»> 72.
org/ScholarlyArticle»> 72.Silvestrini, F., Tiburcio, M., Bertuccini, L. & Alano, P. Дифференциальные адгезивные свойства изолированной бесполой и половой стадий Plasmodium falcipa рома на эндотелиальных клетках человека не зависят от ткани. PLOS ONE 7 , e31567 (2012).
CAS PubMed PubMed Central Google ученый
Fraschka, S.A. et al. Сравнительное профилирование гетерохроматина выявляет консервативные и уникальные сигнатуры эпигенома, связанные с адаптацией и развитием малярийных паразитов. Клеточный микроб-хозяин 23 , 407–420.e8 (2018).
CAS PubMed PubMed Central Google ученый
Messina, V. et al. Гаметоциты малярийного паразита Plasmodium falciparum взаимодействуют с мезенхимальными клетками костного мозга и стимулируют их секретирование ангиогенетических факторов. Фронт. Клетка. Заразить. Microbiol. 8 , 50 (2018). Это исследование предоставляет доказательства связывания бесполых iRBC и гаметоцитов с МСК костного мозга .
Фронт. Клетка. Заразить. Microbiol. 8 , 50 (2018). Это исследование предоставляет доказательства связывания бесполых iRBC и гаметоцитов с МСК костного мозга .
PubMed PubMed Central Google ученый
Neveu, G.и другие. Plasmodium falciparum Эритроциты, инфицированные гаметоцитами, не прикрепляются к первичным эритробластам человека. Sci. Отчет 8 , 17886 (2018).
CAS PubMed PubMed Central Google ученый
Silvestrini, F. et al. Экспорт белка знаменует собой раннюю фазу гаметоцитогенеза малярийного паразита человека Plasmodium falciparum . Мол. Клетка. Протеомика 9 , 1437–1448 (2010).
CAS PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 77.
org/ScholarlyArticle»> 77.Dantzler, K. W. et al. Естественно приобретенный иммунитет против незрелых гаметоцитов Plasmodium falciparum . Sci. Пер. Med. 11 , 1–14 (2019). Это исследование идентифицирует общие антигены на асексуальной и гаметоцитарной поверхности iRBC .
Google ученый
Hermand, P. et al. Plasmodium falciparum белков, участвующих в цитоадгезии инфицированных эритроцитов к хемокину CX3CL1. Sci. Отчет 6 , 33786 (2016).
CAS PubMed PubMed Central Google ученый
Гончаренко М. и др. Стромальные клетки костного мозга человека экспрессируют особый набор биологически функциональных рецепторов хемокинов.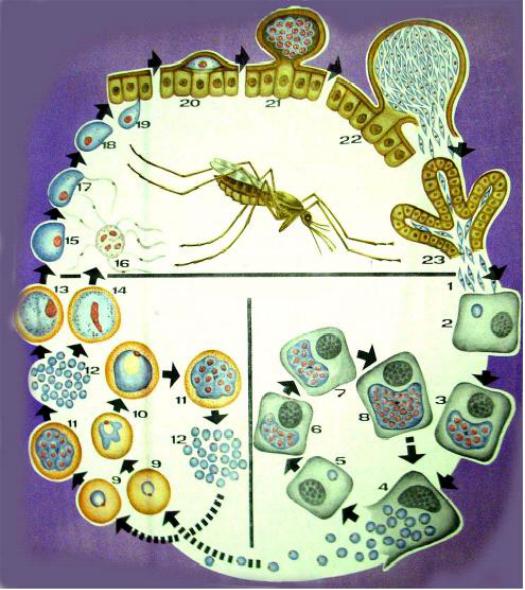 Стволовая клетка 24 , 1030–1041 (2006).
Стволовая клетка 24 , 1030–1041 (2006).
CAS Google ученый
Тамез, П. А., Лю, Х., Фернандес-Поль, С., Халдар, К. и Викрема, А. Стадия-специфическая чувствительность эритробластов человека к Plasmodium falciparum малярийной инфекции. Кровь 114 , 3652–3655 (2009).
CAS PubMed PubMed Central Google ученый
Tiburcio, M. et al.Переключение деформируемости инфицированных эритроцитов при созревании и кровообращении на этапах передачи Plasmodium falciparum . Кровь 119 , e172 – e180 (2012).
CAS PubMed PubMed Central Google ученый
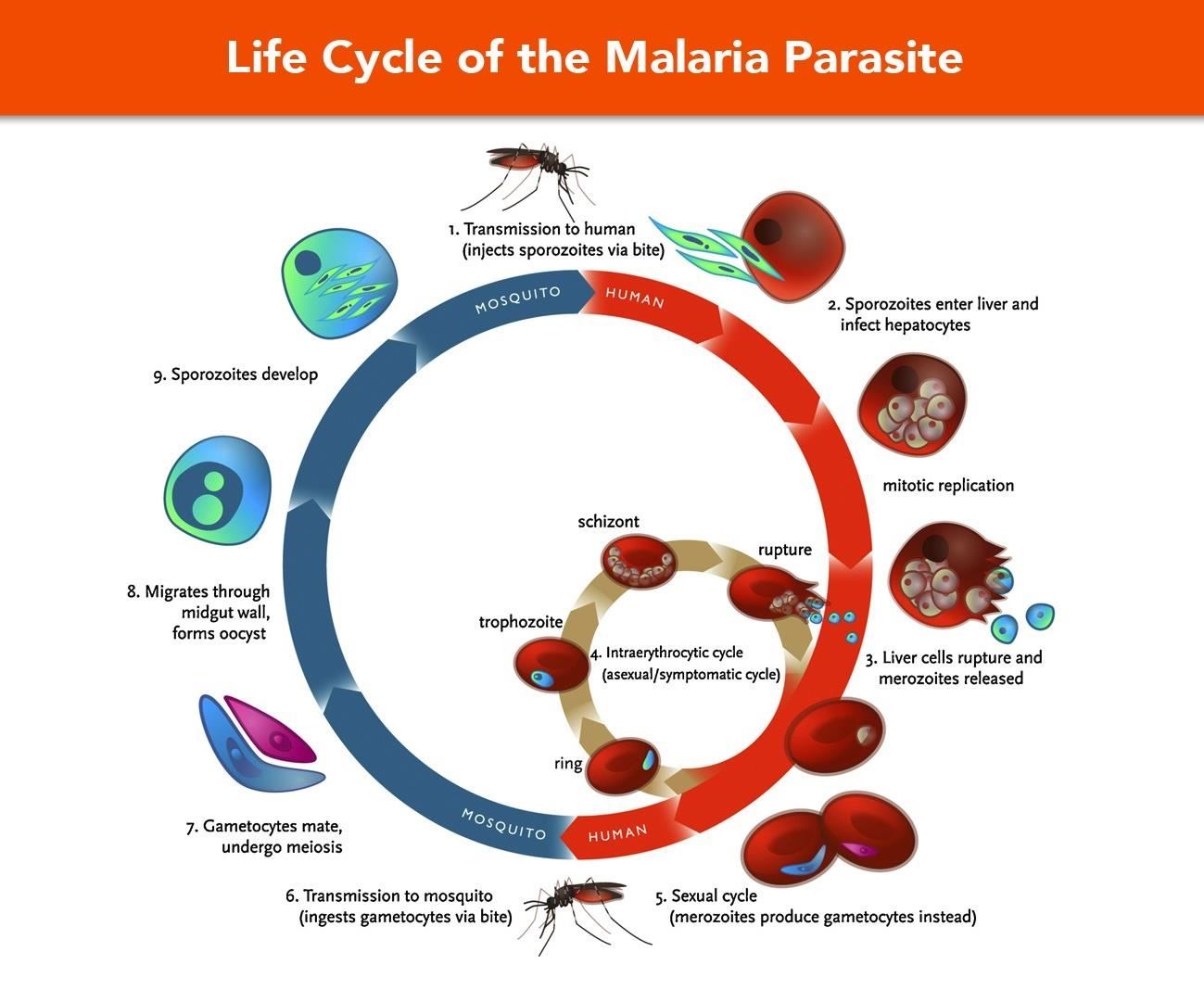 org/ScholarlyArticle»> 82.
org/ScholarlyArticle»> 82.Aingaran, M. et al. Деформируемость клетки-хозяина связана с передачей малярийного паразита человека Plasmodium falciparum . Cell. Microbiol. 14 , 983–993 (2012).
CAS PubMed PubMed Central Google ученый
Во, Р. Э. Жесткость ретикулоцитов и прохождение через поры, подобные эндотелию. Кровь 78 , 3037–3042 (1991).
CAS PubMed Google ученый
Ramdani, G. et al. Передача сигналов цАМФ регулирует деформируемость инфицированных гаметоцитами эритроцитов, необходимую для передачи паразитов малярии. PLOS Pathog. 11 , e1004815 (2015). Это исследование демонстрирует, что на деформируемость гаметоцитов можно воздействовать лекарствами .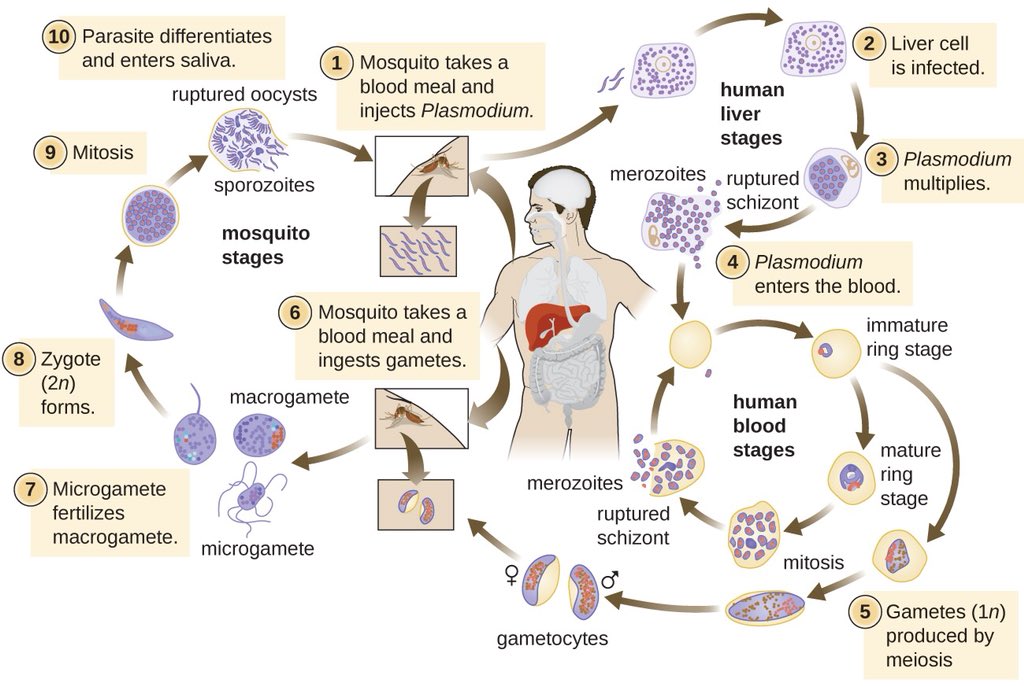
PubMed PubMed Central Google ученый
Naissant, B. et al. Plasmodium falciparum Фосфорилирование STEVOR регулирует деформируемость эритроцитов хозяина, обеспечивая передачу паразитов малярии. Кровь 127 , e42 – e53 (2016).
CAS PubMed Google ученый
Nixon, C. P. Plasmodium falciparum Транзит гаметоцитов через кожные микрососуды: новая цель для вакцин, блокирующих передачу малярии? Гум. Вакцин. Immunother 12 , 3189–3195 (2016).
PubMed PubMed Central Google ученый
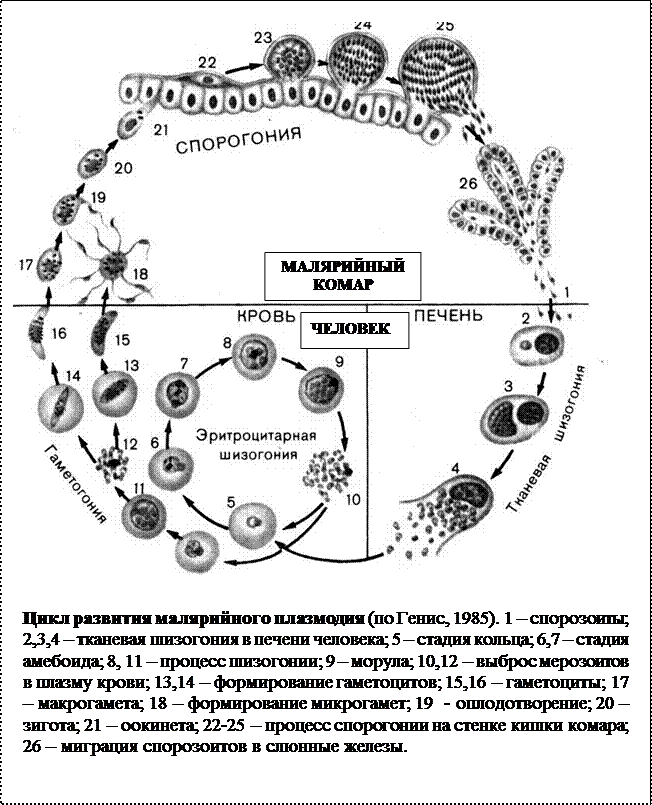
Lawniczak, M. K.И Экхофф П.А. Вычислительная линза для передачи, размножения, приспособленности и кинетики на сексуальной стадии у Plasmodium falciparum . Малар. Дж. 15 , 487 (2016).
PubMed PubMed Central Google ученый
Srivastava, A. et al. Ретикулоциты хозяина являются резервуарами обмена веществ, которые могут использоваться малярийными паразитами. PLOS Pathog. 11 , e1004882 (2015).
PubMed PubMed Central Google ученый
Mu, C. F. et al. Адресная доставка лекарств для лечения опухолей внутри костного мозга. Биоматериалы 155 , 191–202 (2018).
CAS PubMed Google ученый
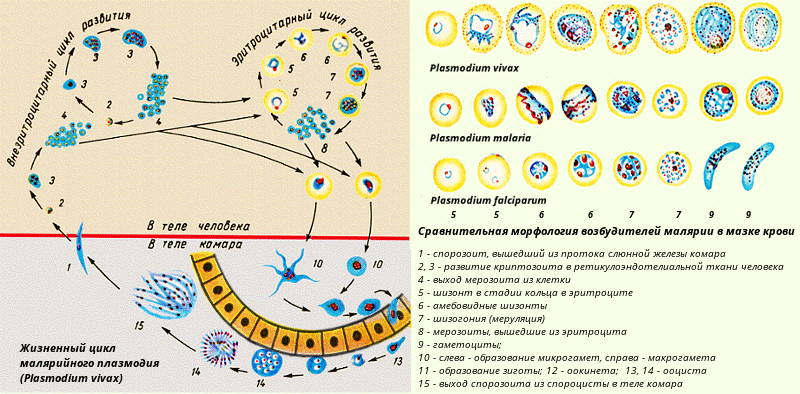 org/ScholarlyArticle»> 90.
org/ScholarlyArticle»> 90.Ng, C. L. & Fidock, D. A. Plasmodium falciparum Выбор устойчивости к лекарствам in vitro и редактирование генов. Methods Mol. Биол. 2013 , 123–140 (2019).
PubMed PubMed Central Google ученый
Camarda, G. et al. Антималярийная активность примахина действует через двухступенчатую биохимическую реле. Nat. Commun. 10 , 3226 (2019).
PubMed PubMed Central Google ученый
Duez, J. et al. Высокопроизводительная микросфильтрация для оценки деформируемости эритроцитов и выявления препаратов, блокирующих передачу малярии. Nat. Protoc. 13 , 1362–1376 (2018).
CAS PubMed Google ученый
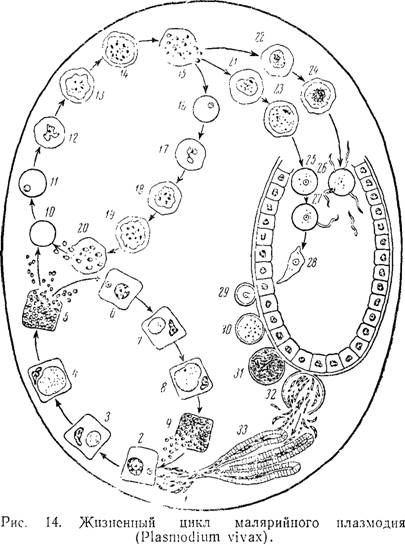 org/ScholarlyArticle»> 93.
org/ScholarlyArticle»> 93.Lavazec, C. & Naissant, B. Виагра делает Plasmodium жестким: новый способ блокировать передачу малярии? [Французский]. Med. Sci. 31 , 826–828 (2015).
Google ученый
Немази, Д. Механизмы центральной толерантности для В-клеток. Nat. Rev. Immunol. 17 , 281–294 (2017).
CAS PubMed PubMed Central Google ученый
Vainieri, M. L. et al. Систематическое отслеживание измененного гематопоэза во время развития малярии, опосредованной спорозоитом, выявляет множественные точки ответа. Открыть Биол . 6 , 1–13 (2016).
Google ученый
 org/ScholarlyArticle»> 96.
org/ScholarlyArticle»> 96.Бокстал В., Геуртс Н. и Магез С. Острое нарушение В-лимфопоэза костного мозга и апоптоз В-клеток переходной и маргинальной зоны в селезенке после инфицирования мышей на стадии крови Plasmodium chabaudi . J. Parasitol. Res. 2011 , 534697 (2011).
PubMed PubMed Central Google ученый
Nduati, E. W. et al. Четкая кинетика ответов В-клеток памяти и плазматических клеток в периферической крови после инфицирования мышей на стадии Plasmodium chabaudi в крови. PLOS ONE 5 , e15007 (2010).
PubMed PubMed Central Google ученый
Маркус, М. Б. Новые доказательства независимости от гипнозоитов Plasmodium vivax рецидивов малярии.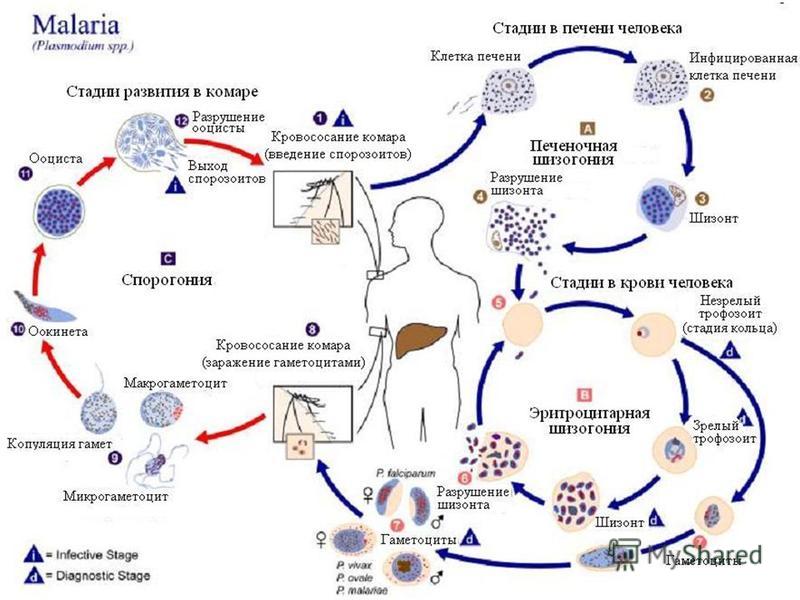 Trends Parasitol. 34 , 1015–1016 (2018).
Trends Parasitol. 34 , 1015–1016 (2018).
PubMed Google ученый
Nothelfer, K., Sansonetti, P.J.& Фалипон, А. Манипуляции с В-клетками патогенами: лучшая защита — это хорошее нападение. Nat. Rev. Microbiol. 13 , 173–184 (2015).
CAS PubMed Google ученый
Das, B. et al. CD271 + мезенхимальные стволовые клетки костного мозга могут обеспечивать нишу для покоящихся Mycobacterium tuberculosis . Sci. Пер. Med. 5 , 170ра113 (2013).
Google ученый
Варма, Н. и Насим, С. Гематологические изменения при висцеральном лейшманиозе / кала азар. Indian J. Hematol. Переливание крови. 26 , 78–82 (2010).
и Насим, С. Гематологические изменения при висцеральном лейшманиозе / кала азар. Indian J. Hematol. Переливание крови. 26 , 78–82 (2010).
PubMed PubMed Central Google ученый
Pinho, S. & Frenette, P. S. Активность гемопоэтических стволовых клеток и взаимодействия с нишей. Nat. Преподобный Мол. Cell Biol. 20 , 303–320 (2019).
CAS PubMed PubMed Central Google ученый
Torisawa, Y. S. et al. Костный мозг-на-чипе воспроизводит физиологию гематопоэтической ниши in vitro. Nat. Meth. 11 , 663–669 (2014).
CAS Google ученый
 org/ScholarlyArticle»> 104.
org/ScholarlyArticle»> 104.Sieber, S. et al. Костный мозг-на-чипе: долгосрочная культура гемопоэтических стволовых клеток человека в трехмерной микрофлюидной среде. J. Tissue Eng. Regen. Med. 12 , 479–489 (2018). Это исследование описывает первую платформу человеческого костного мозга на чипе.
CAS PubMed PubMed Central Google ученый
Rahmig, S. et al. Улучшенный эритропоэз человека и образование тромбоцитов у гуманизированных мышей NSGW41. Stem Cell Rep. 7 , 591–601 (2016).
CAS Google ученый
Baryawno, N. et al. Клеточная систематика стромы костного мозга при гомеостазе и лейкемии. Ячейка 177 , 1915–1932. e16 (2019). Это исследование представляет одноклеточный атлас ниши человеческого костного мозга .
e16 (2019). Это исследование представляет одноклеточный атлас ниши человеческого костного мозга .
CAS PubMed Google ученый
Howick, V. M. et al. Атлас клеток малярии: транскриптомы отдельных паразитов на протяжении полного жизненного цикла Plasmodium . Наука 365 , 1–11 (2019). Это исследование представляет собой одноклеточный атлас жизненного цикла P. falciparum .
Google ученый
Телфорд, С. Р. in Гемопаразиты рептилий: Атлас цветов и текст 376 стр. (CRC, 2009).
Garnham, P. C. C. in Malaria Parasites and Other Haemosporidia 1132 pp. (Blackwell, 1966).
(Blackwell, 1966).
Валкюнас, Г. в Птичьи малярийные паразиты и другие гемоспоридии 946 стр. (CRC Press, 2005).
Палинаускас В. и др. Описание, молекулярная характеристика, диагностика и жизненный цикл Plasmodium elongatum (линия pERIRUB01), вирулентного птичьего малярийного паразита. Внутр. J. Parasitol. 46 , 697–707 (2016). Эта работа является одним из нескольких исследований птичьей малярии, в которых описываются эритроцитарные шизонты в костном мозге инфицированных птиц .
CAS PubMed Google ученый
Перкинс, С. Л. Множество друзей Малярии: прошлое, настоящее и будущее систематиков отряда Haemosporida.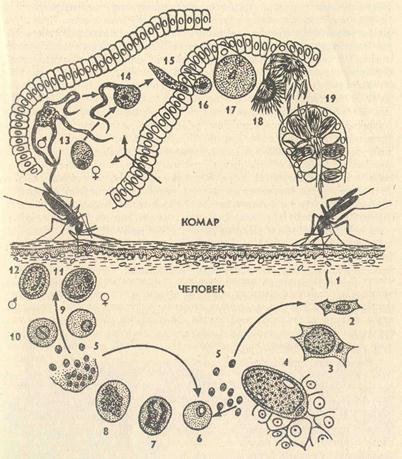 J. Parasitol. 100 , 11–25 (2014).
J. Parasitol. 100 , 11–25 (2014).
PubMed Google ученый
MacCallum, W.G. О жгутиковых формах малярийного паразита. Ланцет 11 , 1240–1241 (1897).
Google ученый
Aragão, H. B. Über den Entwicklungsgang и die Übertragung von Haemoproteus columbae. Arch. Protistenkunde 12 , 154–167 (1908).
Google ученый
Джеймс, С. П. и Тейт, П. Новые знания о жизненном цикле малярийных паразитов. Природа 139 , 545 (1937).
Google ученый
 org/ScholarlyArticle»> 116.
org/ScholarlyArticle»> 116.Росс, Р.Отчет о культивировании Proteosoma, Labbé у серых комаров. Indian Med. Газ. 33 , 401–408 (1898).
Google ученый
Коатни, Г. Р., Купер, В. К., Эдди, Н. Б. и Гринберг, Дж. Обзор противомалярийных средств: химиотерапия инфекций, вызванных Plasmodium gallinaceum ; токсичность; соотношение структуры и действия. Моногр. Общественного здравоохранения. 9 , 1–322 (1953).
CAS PubMed Google ученый
Хокинг Ф. Культура тканей малярийных паразитов ( Plasmodium gallinaceum ). Ланцет 246 , 693–694 (1944).
Google ученый
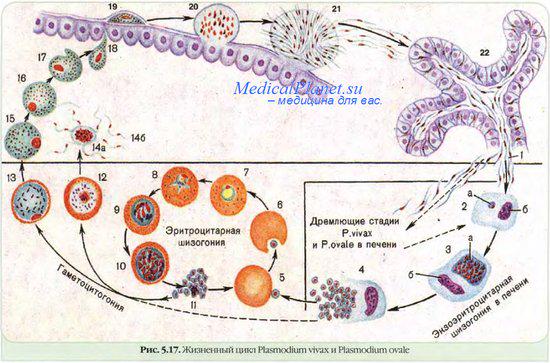 org/ScholarlyArticle»> 119.
org/ScholarlyArticle»> 119.Ilgunas, M. et al. Паттерны вирулентности Plasmodium homocircumflexum у экспериментально зараженных воробьиных птиц. Малар.Дж. 18 , 174 (2019).
PubMed PubMed Central Google ученый
Валкюнас, Г. и Иежова, Т. А. Экзоэритроцитарное развитие малярии птиц и родственных гемоспоридических паразитов. Малар. Дж. 16 , 101 (2017). Настоящий обзор — первый, посвященный экзоэритроцитарному развитию малярийных паразитов птиц, включая клетки гематопоэтической ниши .
PubMed PubMed Central Google ученый
Бен-Харел С. Исследования птичьей малярии в связи с механизмом рецидива. Am. J. Hyg. 3 , 652 (1923).
Google ученый
Ilgunas, M., Palinauskas, V., Platonova, E., Iezhova, T. & Valkiunas, G. Экспериментальное исследование восприимчивости обычных европейских певчих птиц к Plasmodium elongatum (линия pGRW6), a широко распространенный птичий малярийный паразит. Малар. J. 18 , 290 (2019). Это исследование предоставляет доказательства того, что вирулентность связана с повреждением клеток костного мозга во время птичьей малярии .
PubMed PubMed Central Google ученый
Лейнсон, Р. в Атлас простейших паразитов амазонской фауны Бразилии . Том 1: Гемоспоридии рептилий 81 стр. (Instituto Evandro Chagas, 2012).
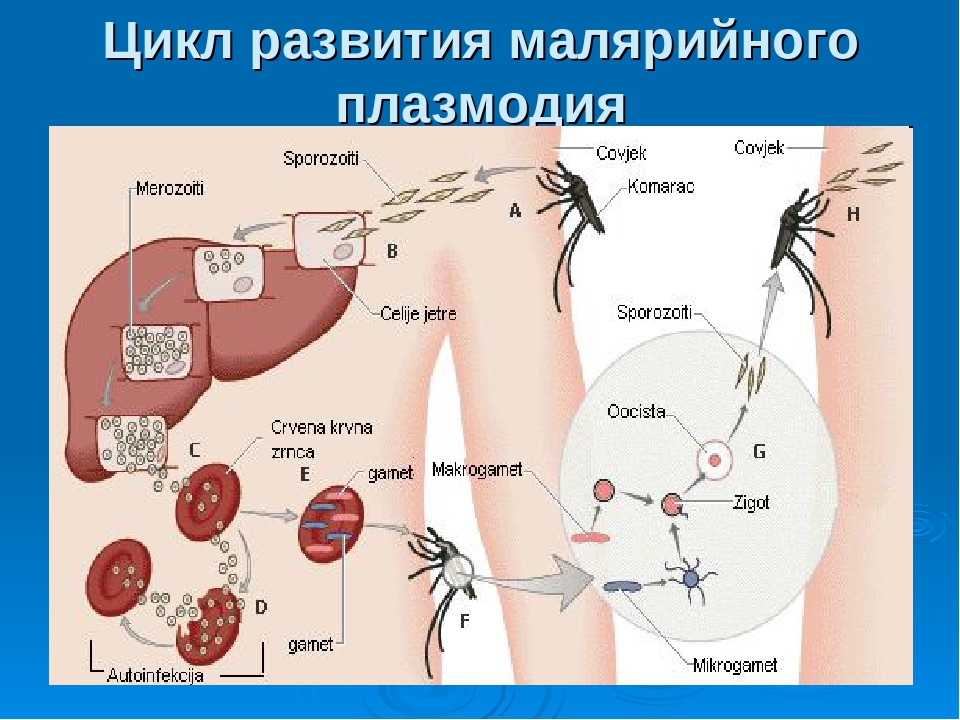 org/ScholarlyArticle»> 124.
org/ScholarlyArticle»> 124.Schaer, J. et al. Никтерии-паразиты афротропических насекомоядных летучих мышей. Внутр. J. Parasitol. 45 , 375–384 (2015).
CAS PubMed Google ученый
Schaer, J. et al. Летучие мыши с эполетами демонстрируют исключительно высокий уровень инфицирования комплексом видов Hepatocystis в Южном Судане. Sci. Отчет 7 , 6928 (2017).
PubMed PubMed Central Google ученый
Гален С.C. et al. Полифилия Plasmodium : всесторонний филогенетический анализ паразитов малярии (отряд Haemosporida) выявил широко распространенный таксономический конфликт. R. Soc. Открыть. Sci. 5 , 171780 (2018). Это исследование представляет собой всесторонний филогенетический анализ гемоспоридий, предлагающий полифилетический статус рода Plasmodium .
PubMed PubMed Central Google ученый
Borner, J. et al. Филогения гемоспоридий паразитов крови, выявленная с помощью мультигенного подхода. Мол. Филогенет. Evol. 94 , 221–231 (2016).
CAS PubMed Google ученый
Гален, С.С., Нунес, Р., Свит, П.Р. и Перкинс, С.Л. Объединение границ сливающихся видов с анализом специфичности хозяина показывает обширное скрытое разнообразие, несмотря на минимальное расхождение митохондрий у малярийных паразитов рода Leucocytozoon. BMC Evol. Биол. 18 , 128 (2018).
PubMed PubMed Central Google ученый
% PDF-1.5
%
4 0 obj
>
эндобдж
xref
4 1707
0000000016 00000 н.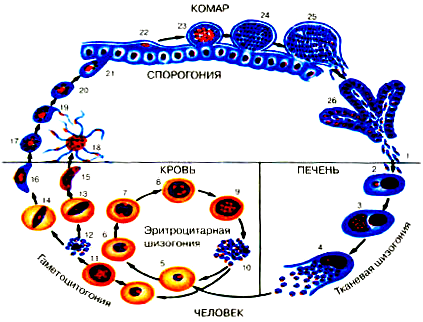 0000037480 00000 п.
0000037576 00000 п.
0000060624 00000 п.
0000060670 00000 п.
0000060715 00000 п.
0000060761 00000 п.
0000060808 00000 п.
0000060855 00000 п.
0000060901 00000 п.
0000060947 00000 п.
0000060993 00000 п.
0000061039 00000 п.
0000061085 00000 п.
0000061132 00000 п.
0000061179 00000 п.
0000061225 00000 п.
0000061272 00000 п.
0000061318 00000 п.
0000061364 00000 п.
0000061410 00000 п.
0000061457 00000 п.
0000061503 00000 п.
0000061550 00000 п.
0000061596 00000 п.
0000061643 00000 п.
0000061689 00000 п.
0000061736 00000 п.
0000061782 00000 п.
0000061829 00000 п.
0000061876 00000 п.
0000061922 00000 п.
0000061968 00000 п.
0000062014 00000 п.
0000062060 00000 п.
0000062105 00000 п.
0000062152 00000 п.
0000062198 00000 п.
0000062245 00000 п.
0000062292 00000 п.
0000062339 00000 п.
0000062386 00000 п.
0000062433 00000 п.
0000062480 00000 п.
0000062526 00000 п.
0000062572 00000 п.
0000062619 00000 п.
0000062665 00000 п.
0000062711 00000 п.
0000037480 00000 п.
0000037576 00000 п.
0000060624 00000 п.
0000060670 00000 п.
0000060715 00000 п.
0000060761 00000 п.
0000060808 00000 п.
0000060855 00000 п.
0000060901 00000 п.
0000060947 00000 п.
0000060993 00000 п.
0000061039 00000 п.
0000061085 00000 п.
0000061132 00000 п.
0000061179 00000 п.
0000061225 00000 п.
0000061272 00000 п.
0000061318 00000 п.
0000061364 00000 п.
0000061410 00000 п.
0000061457 00000 п.
0000061503 00000 п.
0000061550 00000 п.
0000061596 00000 п.
0000061643 00000 п.
0000061689 00000 п.
0000061736 00000 п.
0000061782 00000 п.
0000061829 00000 п.
0000061876 00000 п.
0000061922 00000 п.
0000061968 00000 п.
0000062014 00000 п.
0000062060 00000 п.
0000062105 00000 п.
0000062152 00000 п.
0000062198 00000 п.
0000062245 00000 п.
0000062292 00000 п.
0000062339 00000 п.
0000062386 00000 п.
0000062433 00000 п.
0000062480 00000 п.
0000062526 00000 п.
0000062572 00000 п.
0000062619 00000 п.
0000062665 00000 п.
0000062711 00000 п.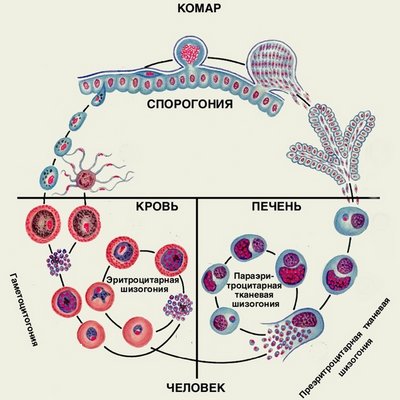 0000062757 00000 п.
0000062804 00000 п.
0000062851 00000 п.
0000062897 00000 п.
0000062944 00000 п.
0000062991 00000 п.
0000063038 00000 п.
0000063084 00000 п.
0000063130 00000 н.
0000063177 00000 п.
0000063223 00000 п.
0000063269 00000 п.
0000063315 00000 п.
0000063360 00000 п.
0000063407 00000 п.
0000063454 00000 п.
0000063500 00000 п.
0000063547 00000 п.
0000063594 00000 п.
0000063639 00000 п.
0000063685 00000 п.
0000063730 00000 п.
0000063777 00000 п.
0000063824 00000 п.
0000063870 00000 п.
0000063916 00000 п.
0000063962 00000 п.
0000064008 00000 п.
0000064053 00000 п.
0000064098 00000 п.
0000064144 00000 п.
0000064190 00000 п.
0000064236 00000 п.
0000064282 00000 п.
0000064328 00000 н.
0000064374 00000 п.
0000064420 00000 н.
0000064466 00000 н.
0000064512 00000 п.
0000064558 00000 п.
0000064604 00000 п.
0000064650 00000 п.
0000064696 00000 п.
0000064743 00000 п.
0000064789 00000 п.
0000064834 00000 п.
0000064881 00000 п.
0000064928 00000 п.
0000062757 00000 п.
0000062804 00000 п.
0000062851 00000 п.
0000062897 00000 п.
0000062944 00000 п.
0000062991 00000 п.
0000063038 00000 п.
0000063084 00000 п.
0000063130 00000 н.
0000063177 00000 п.
0000063223 00000 п.
0000063269 00000 п.
0000063315 00000 п.
0000063360 00000 п.
0000063407 00000 п.
0000063454 00000 п.
0000063500 00000 п.
0000063547 00000 п.
0000063594 00000 п.
0000063639 00000 п.
0000063685 00000 п.
0000063730 00000 п.
0000063777 00000 п.
0000063824 00000 п.
0000063870 00000 п.
0000063916 00000 п.
0000063962 00000 п.
0000064008 00000 п.
0000064053 00000 п.
0000064098 00000 п.
0000064144 00000 п.
0000064190 00000 п.
0000064236 00000 п.
0000064282 00000 п.
0000064328 00000 н.
0000064374 00000 п.
0000064420 00000 н.
0000064466 00000 н.
0000064512 00000 п.
0000064558 00000 п.
0000064604 00000 п.
0000064650 00000 п.
0000064696 00000 п.
0000064743 00000 п.
0000064789 00000 п.
0000064834 00000 п.
0000064881 00000 п.
0000064928 00000 п.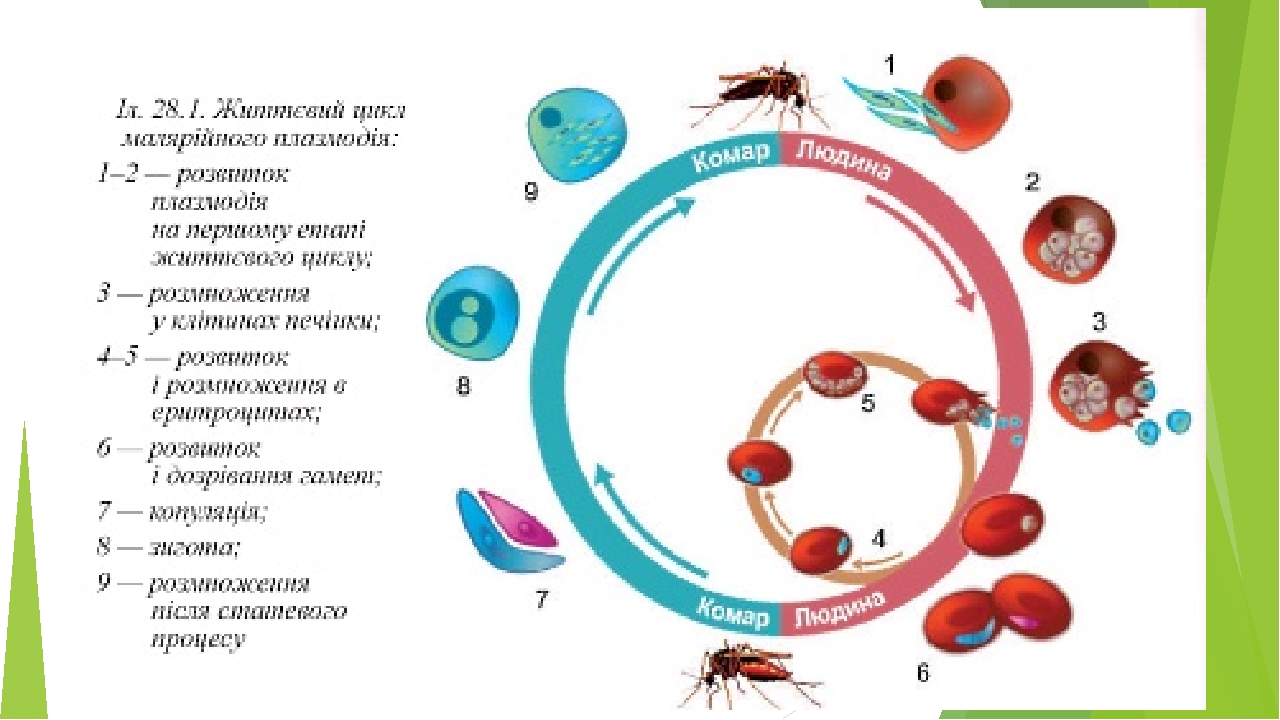 0000064975 00000 п.
0000065022 00000 п.
0000065059 00000 п.
0000065105 00000 п.
0000065153 00000 п.
0000065200 00000 н.
0000065248 00000 п.
0000065296 00000 п.
0000065344 00000 п.
0000065391 00000 п.
0000065438 00000 п.
0000065484 00000 п.
0000065532 00000 п.
0000065580 00000 п.
0000065628 00000 п.
0000065676 00000 п.
0000065724 00000 п.
0000065772 00000 п.
0000065819 00000 п.
0000065867 00000 п.
0000065914 00000 п.
0000065962 00000 п.
0000066010 00000 п.
0000066058 00000 п.
0000066105 00000 п.
0000066153 00000 п.
0000066201 00000 п.
0000066249 00000 п.
0000066296 00000 п.
0000066343 00000 п.
0000132917 00000 н.
0000209071 00000 н.
0000279566 00000 н.
0000348200 00000 н.
0000413758 00000 н.
0000414273 00000 н.
0000414484 00000 н.
0000480898 00000 н.
0000481126 00000 н.
0000481354 00000 н.
0000481608 00000 н.
0000481821 00000 н.
0000482034 00000 н.
0000482245 00000 н.
0000482460 00000 н.
0000482645 00000 н.
0000482920 00000 н.
0000483167 00000 н.
0000064975 00000 п.
0000065022 00000 п.
0000065059 00000 п.
0000065105 00000 п.
0000065153 00000 п.
0000065200 00000 н.
0000065248 00000 п.
0000065296 00000 п.
0000065344 00000 п.
0000065391 00000 п.
0000065438 00000 п.
0000065484 00000 п.
0000065532 00000 п.
0000065580 00000 п.
0000065628 00000 п.
0000065676 00000 п.
0000065724 00000 п.
0000065772 00000 п.
0000065819 00000 п.
0000065867 00000 п.
0000065914 00000 п.
0000065962 00000 п.
0000066010 00000 п.
0000066058 00000 п.
0000066105 00000 п.
0000066153 00000 п.
0000066201 00000 п.
0000066249 00000 п.
0000066296 00000 п.
0000066343 00000 п.
0000132917 00000 н.
0000209071 00000 н.
0000279566 00000 н.
0000348200 00000 н.
0000413758 00000 н.
0000414273 00000 н.
0000414484 00000 н.
0000480898 00000 н.
0000481126 00000 н.
0000481354 00000 н.
0000481608 00000 н.
0000481821 00000 н.
0000482034 00000 н.
0000482245 00000 н.
0000482460 00000 н.
0000482645 00000 н.
0000482920 00000 н.
0000483167 00000 н. 0000483788 00000 н.
0000484025 00000 н.
0000484192 00000 н.
0000551215 00000 н.
0000613669 00000 н.
0000616318 00000 н.
0000616355 00000 н.
0000616506 00000 н.
0000616654 00000 н.
0000617027 00000 н.
0000617390 00000 н.
0000617464 00000 н.
0000617796 00000 н.
0000618403 00000 п.
0000618663 00000 н.
0000618794 00000 н.
0000618940 00000 н.
0000619056 00000 н.
0000619172 00000 п.
0000619322 00000 н.
0000619439 00000 н.
0000619636 00000 п.
0000619845 00000 н.
0000620056 00000 н.
0000620280 00000 н.
0000620499 00000 н.
0000620705 00000 н.
0000620773 00000 н.
0000620838 00000 н.
0000621019 00000 н.
0000621140 00000 н.
0000621323 00000 н.
0000621665 00000 н.
0000621991 00000 н.
0000622315 00000 н.
0000622855 00000 н.
0000623548 00000 н.
0000624105 00000 н.
0000624381 00000 п.
0000624936 00000 н.
0000625507 00000 н.
0000626216 00000 н.
0000626302 00000 н.
0000626893 00000 н.
0000627190 00000 н.
0000627747 00000 н.
0000628266 00000 н.
0000628822 00000 н.
0000483788 00000 н.
0000484025 00000 н.
0000484192 00000 н.
0000551215 00000 н.
0000613669 00000 н.
0000616318 00000 н.
0000616355 00000 н.
0000616506 00000 н.
0000616654 00000 н.
0000617027 00000 н.
0000617390 00000 н.
0000617464 00000 н.
0000617796 00000 н.
0000618403 00000 п.
0000618663 00000 н.
0000618794 00000 н.
0000618940 00000 н.
0000619056 00000 н.
0000619172 00000 п.
0000619322 00000 н.
0000619439 00000 н.
0000619636 00000 п.
0000619845 00000 н.
0000620056 00000 н.
0000620280 00000 н.
0000620499 00000 н.
0000620705 00000 н.
0000620773 00000 н.
0000620838 00000 н.
0000621019 00000 н.
0000621140 00000 н.
0000621323 00000 н.
0000621665 00000 н.
0000621991 00000 н.
0000622315 00000 н.
0000622855 00000 н.
0000623548 00000 н.
0000624105 00000 н.
0000624381 00000 п.
0000624936 00000 н.
0000625507 00000 н.
0000626216 00000 н.
0000626302 00000 н.
0000626893 00000 н.
0000627190 00000 н.
0000627747 00000 н.
0000628266 00000 н.
0000628822 00000 н.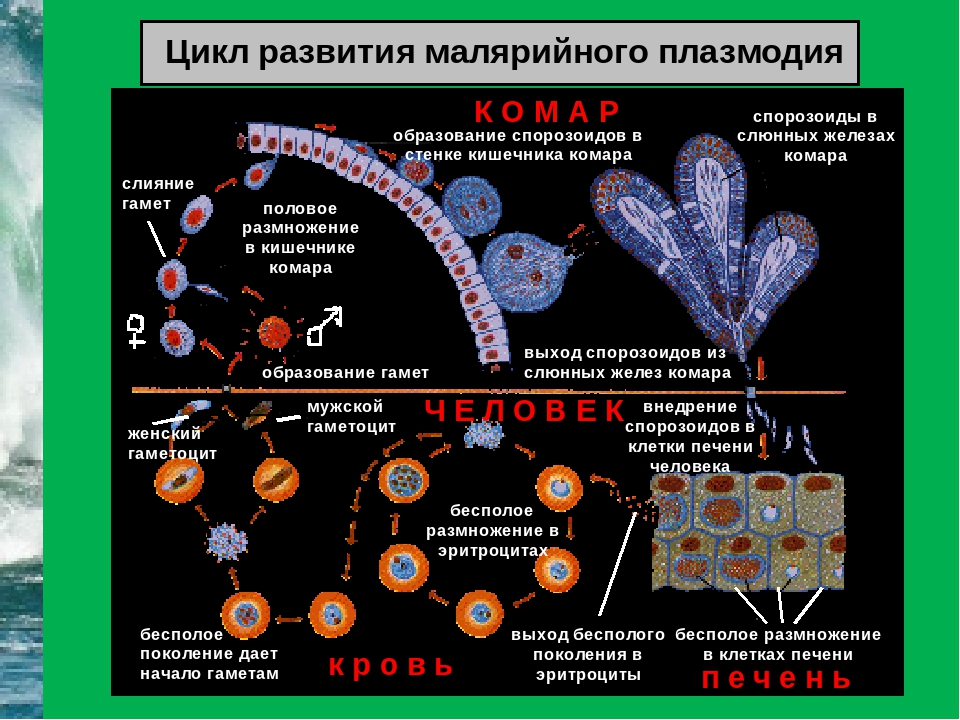 0000629424 00000 н.
0000629790 00000 н.
0000630358 00000 п.
0000630410 00000 н.
0000630593 00000 н.
0000630755 00000 н.
0000631170 00000 н.
0000631690 00000 н.
0000632206 00000 н.
0000632502 00000 н.
0000633115 00000 п.
0000633170 00000 н.
0000633484 00000 н.
0000633567 00000 н.
0000634110 00000 н.
0000634256 00000 н.
0000634911 00000 п.
0000635067 00000 н.
0000635622 00000 н.
0000635749 00000 п.
0000636036 00000 н.
0000636399 00000 н.
0000636542 00000 н.
0000636856 00000 н.
0000637158 00000 п.
0000637273 00000 н.
0000637331 00000 п.
0000637669 00000 н.
0000637968 00000 н.
0000638527 00000 н.
0000638643 00000 п.
0000638994 00000 п.
0000639646 00000 н.
0000640247 00000 н.
0000640506 00000 н.
0000640781 00000 н.
0000640885 00000 н.
0000641003 00000 н.
0000641146 00000 н.
0000641478 00000 н.
0000641777 00000 н.
0000642098 00000 н.
0000642795 00000 н.
0000642982 00000 н.
0000643089 00000 н.
0000643399 00000 н.
0000644031 00000 н.
0000644196 00000 н.
0000629424 00000 н.
0000629790 00000 н.
0000630358 00000 п.
0000630410 00000 н.
0000630593 00000 н.
0000630755 00000 н.
0000631170 00000 н.
0000631690 00000 н.
0000632206 00000 н.
0000632502 00000 н.
0000633115 00000 п.
0000633170 00000 н.
0000633484 00000 н.
0000633567 00000 н.
0000634110 00000 н.
0000634256 00000 н.
0000634911 00000 п.
0000635067 00000 н.
0000635622 00000 н.
0000635749 00000 п.
0000636036 00000 н.
0000636399 00000 н.
0000636542 00000 н.
0000636856 00000 н.
0000637158 00000 п.
0000637273 00000 н.
0000637331 00000 п.
0000637669 00000 н.
0000637968 00000 н.
0000638527 00000 н.
0000638643 00000 п.
0000638994 00000 п.
0000639646 00000 н.
0000640247 00000 н.
0000640506 00000 н.
0000640781 00000 н.
0000640885 00000 н.
0000641003 00000 н.
0000641146 00000 н.
0000641478 00000 н.
0000641777 00000 н.
0000642098 00000 н.
0000642795 00000 н.
0000642982 00000 н.
0000643089 00000 н.
0000643399 00000 н.
0000644031 00000 н.
0000644196 00000 н.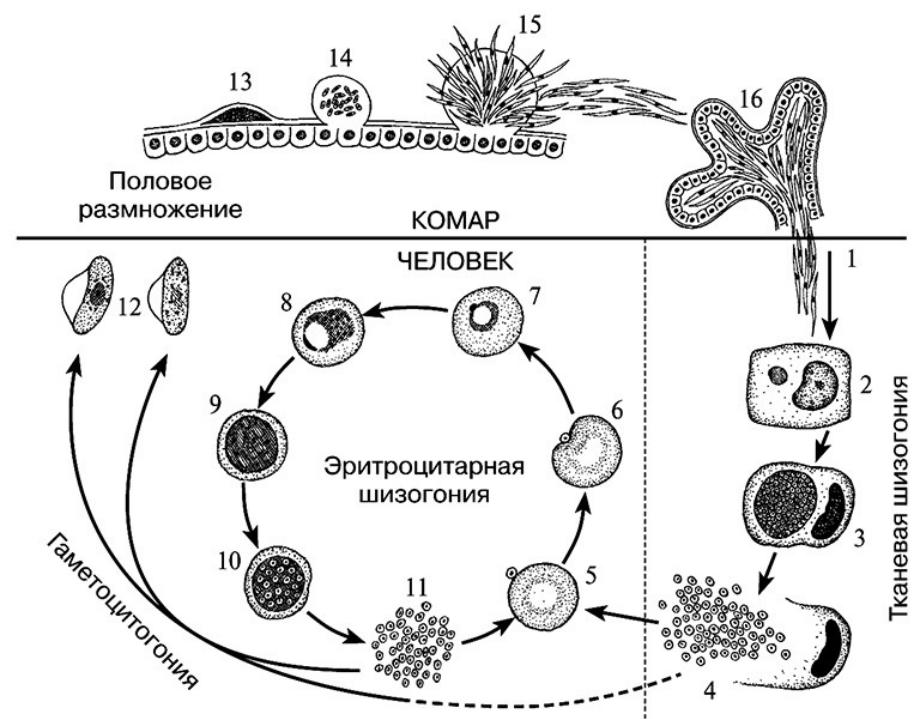 0000644279 00000 н.
0000644860 00000 н.
0000644973 00000 н.
0000645161 00000 п.
0000645292 00000 п.
0000645369 00000 п.
0000645941 00000 п.
0000646458 00000 н.
0000646682 00000 н.
0000647220 00000 н.
0000647685 00000 н.
0000647762 00000 н.
0000647881 00000 н.
0000647936 00000 н.
0000648467 00000 н.
0000649015 00000 н.
0000649483 00000 н.
0000649859 00000 н.
0000650234 00000 н.
0000650599 00000 н.
0000650972 00000 н.
0000651339 00000 н.
0000651457 00000 н.
0000651751 00000 н.
0000652129 00000 н.
0000652303 00000 н.
0000652594 00000 н.
0000652870 00000 н.
0000653466 00000 н.
0000653920 00000 н.
0000654267 00000 н.
0000654637 00000 н.
0000655018 00000 н.
0000655132 00000 н.
0000655163 00000 н.
0000655238 00000 п.
0000661386 00000 н.
0000661710 00000 н.
0000661776 00000 н.
0000661892 00000 н.
0000661923 00000 н.
0000661998 00000 н.
0000667873 00000 п.
0000668196 00000 п.
0000668262 00000 н.
0000668378 00000 п.
0000668409 00000 н.
0000668484 00000 н.
0000644279 00000 н.
0000644860 00000 н.
0000644973 00000 н.
0000645161 00000 п.
0000645292 00000 п.
0000645369 00000 п.
0000645941 00000 п.
0000646458 00000 н.
0000646682 00000 н.
0000647220 00000 н.
0000647685 00000 н.
0000647762 00000 н.
0000647881 00000 н.
0000647936 00000 н.
0000648467 00000 н.
0000649015 00000 н.
0000649483 00000 н.
0000649859 00000 н.
0000650234 00000 н.
0000650599 00000 н.
0000650972 00000 н.
0000651339 00000 н.
0000651457 00000 н.
0000651751 00000 н.
0000652129 00000 н.
0000652303 00000 н.
0000652594 00000 н.
0000652870 00000 н.
0000653466 00000 н.
0000653920 00000 н.
0000654267 00000 н.
0000654637 00000 н.
0000655018 00000 н.
0000655132 00000 н.
0000655163 00000 н.
0000655238 00000 п.
0000661386 00000 н.
0000661710 00000 н.
0000661776 00000 н.
0000661892 00000 н.
0000661923 00000 н.
0000661998 00000 н.
0000667873 00000 п.
0000668196 00000 п.
0000668262 00000 н.
0000668378 00000 п.
0000668409 00000 н.
0000668484 00000 н. 0000673699 00000 н.
0000674020 00000 н.
0000674086 00000 н.
0000674202 00000 н.
0000674392 00000 н.
0000676860 00000 н.
0000677236 00000 п.
0000677349 00000 н.
0000678420 00000 н.
0000678734 00000 н.
0000679032 00000 н.
0000682281 00000 н.
0000682755 00000 н.
0000683295 00000 н.
0000683419 00000 п.
0000684951 00000 н.
0000685278 00000 н.
0000685659 00000 н.
0000685800 00000 н.
0000687752 00000 н.
0000688089 00000 н.
0000688514 00000 н.
0000688609 00000 н.
00006
0000673699 00000 н.
0000674020 00000 н.
0000674086 00000 н.
0000674202 00000 н.
0000674392 00000 н.
0000676860 00000 н.
0000677236 00000 п.
0000677349 00000 н.
0000678420 00000 н.
0000678734 00000 н.
0000679032 00000 н.
0000682281 00000 н.
0000682755 00000 н.
0000683295 00000 н.
0000683419 00000 п.
0000684951 00000 н.
0000685278 00000 н.
0000685659 00000 н.
0000685800 00000 н.
0000687752 00000 н.
0000688089 00000 н.
0000688514 00000 н.
0000688609 00000 н.
00006
00000 п. 00006
00000 н. 0000694107 00000 п. 0000694454 00000 п. 0000694575 00000 п. 0000694709 00000 н. 0000694859 00000 н. 0000696745 00000 н. 0000697110 00000 н. 0000697260 00000 н. 0000697377 00000 п. 0000698703 00000 п. 0000699015 00000 н. 0000699391 00000 п. 0000826094 00000 н. 0000826481 00000 н. 0000826578 00000 н.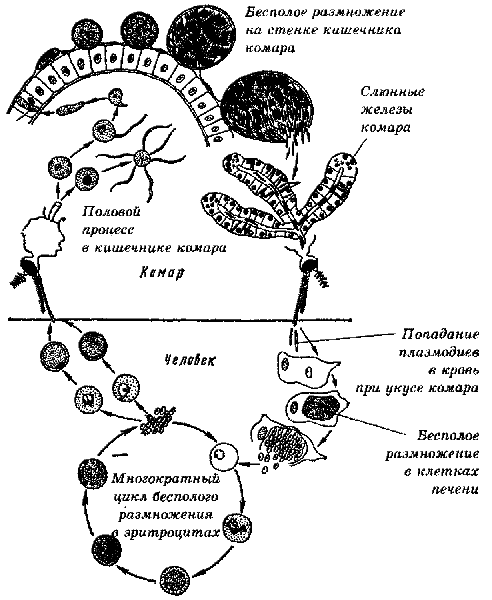 0000826724 00000 н.
0000827111 00000 н.
0000827208 00000 н.
0000827354 00000 н.
0000834359 00000 н.
0000838659 00000 н.
0000842300 00000 н.
0000846828 00000 н.
0000850992 00000 н.
0000854077 00000 н.
0000864424 00000 н.
0000873361 00000 н.
0000880710 00000 н.
0000884649 00000 н.
0000888929 00000 н.
0000893514 00000 н.
00004 00000 н.
0000
0000826724 00000 н.
0000827111 00000 н.
0000827208 00000 н.
0000827354 00000 н.
0000834359 00000 н.
0000838659 00000 н.
0000842300 00000 н.
0000846828 00000 н.
0000850992 00000 н.
0000854077 00000 н.
0000864424 00000 н.
0000873361 00000 н.
0000880710 00000 н.
0000884649 00000 н.
0000888929 00000 н.
0000893514 00000 н.
00004 00000 н.
00002 00000 н. 00006 00000 н. 0000
9 00000 п. 0000930230 00000 н. 0000934369 00000 п. 0000940379 00000 н. 0000946732 00000 н. 0000950583 00000 н. 0000956412 00000 н. 0000960297 00000 н. 0000967293 00000 п. 0000970938 00000 п. 0000981932 00000 н. 0000988038 00000 н. 0001011792 00000 п. 0001022746 00000 п. 0001027243 00000 п. 0001028756 00000 п. 0001033175 00000 п. 0001037345 00000 п. 0001057349 00000 п. 0001062381 00000 п. 0001067040 00000 п. 0001074438 00000 п. 0001079558 00000 п. 0001085415 00000 п. 00010 00000 п. 0001095644 00000 п. 0001101409 00000 п. 0001105533 00000 п. 0001109553 00000 п.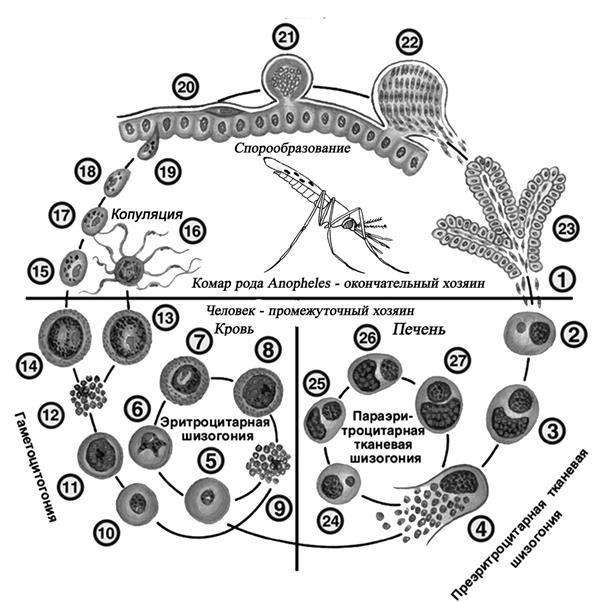 0001111694 00000 п.
0001118034 00000 п.
0001129987 00000 п.
0001134389 00000 п.
0001138191 00000 п.
0001144201 00000 п.
0001150679 00000 п.
0001152945 00000 п.
0001284636 00000 п.
0001315701 00000 п.
0001367766 00000 п.
0001370375 00000 п.
0001370605 00000 п.
0001370974 00000 п.
0001371365 00000 п.
0001371511 00000 п.
0001371665 00000 п.
0001403849 00000 п.
0001458029 00000 п.
00014
0001111694 00000 п.
0001118034 00000 п.
0001129987 00000 п.
0001134389 00000 п.
0001138191 00000 п.
0001144201 00000 п.
0001150679 00000 п.
0001152945 00000 п.
0001284636 00000 п.
0001315701 00000 п.
0001367766 00000 п.
0001370375 00000 п.
0001370605 00000 п.
0001370974 00000 п.
0001371365 00000 п.
0001371511 00000 п.
0001371665 00000 п.
0001403849 00000 п.
0001458029 00000 п.
0001400000 п. 0001543898 00000 п. 0001555882 00000 п. 0001555979 00000 п. 0001556133 00000 п. 0001566867 00000 п. 0001577704 00000 п. 0001589001 00000 п. 00015
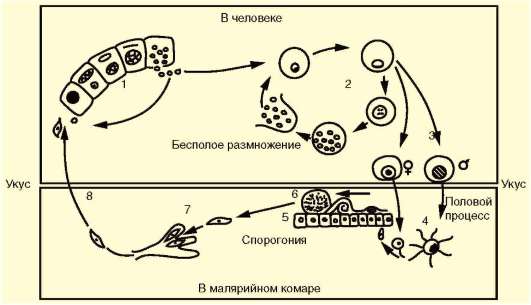 0001703596 00000 п.
0001703742 00000 п.
0001703896 00000 п.
0001736633 00000 п.
00017 00000 пн
0001824689 00000 п.
0001824918 00000 п.
0001825318 00000 п.
0001825439 00000 п.
0001825593 00000 п.
0001827576 00000 п.
0001834762 00000 п.
0001835149 00000 п.
0001835378 00000 п.
0001835499 00000 п.
0001835653 00000 п.
0001837696 00000 п.
0001839211 00000 п.
0001846410 00000 п.
0001848431 00000 н.
0001855485 00000 п.
0001857504 00000 н.
0001864555 00000 п.
0001866579 00000 п.
0001873623 00000 п.
0001875681 00000 п.
0001882760 00000 п.
0001884779 00000 п.
0001893111 00000 п.
0001
0001703596 00000 п.
0001703742 00000 п.
0001703896 00000 п.
0001736633 00000 п.
00017 00000 пн
0001824689 00000 п.
0001824918 00000 п.
0001825318 00000 п.
0001825439 00000 п.
0001825593 00000 п.
0001827576 00000 п.
0001834762 00000 п.
0001835149 00000 п.
0001835378 00000 п.
0001835499 00000 п.
0001835653 00000 п.
0001837696 00000 п.
0001839211 00000 п.
0001846410 00000 п.
0001848431 00000 н.
0001855485 00000 п.
0001857504 00000 н.
0001864555 00000 п.
0001866579 00000 п.
0001873623 00000 п.
0001875681 00000 п.
0001882760 00000 п.
0001884779 00000 п.
0001893111 00000 п.
0001 0001959760 00000 п.
0001961693 00000 п.
0001968357 00000 п.
0001970349 00000 п.
0001977130 00000 п.
0001979098 00000 н.
0001985773 00000 п.
0001987711 00000 п.
00019
0001959760 00000 п.
0001961693 00000 п.
0001968357 00000 п.
0001970349 00000 п.
0001977130 00000 п.
0001979098 00000 н.
0001985773 00000 п.
0001987711 00000 п.
0001900000 п. 0001998618 00000 п. 0002000549 00000 п. 0002007232 00000 н. 0002009158 00000 n 0002015846 00000 н. 0002017753 00000 п. 0002024500 00000 н. 0002026460 00000 п. 0002033186 00000 п. 0002035177 00000 п. 0002041932 00000 н. 0002048674 00000 н. 0002050650 00000 п. 0002057427 00000 н. 0002059391 00000 п. 0002066084 00000 п. 0002068056 00000 п. 0002074780 00000 п. 0002076736 00000 н. 0002083440 00000 п. 0002085464 00000 п. 0002092945 00000 пн 0002100052 00000 п. 0002102061 00000 п. 0002109072 00000 п. 0002111106 00000 п. 0002118167 00000 п. 0002118397 00000 п. 0002118784 00000 п. 0002118905 00000 п. 0002119072 00000 н. 0002119459 00000 п. 0002119580 00000 п. 0002119747 00000 п. 0002119977 00000 н. 0002120364 00000 п. 0002120485 00000 н. 0002120652 00000 п. 0002120806 00000 п.
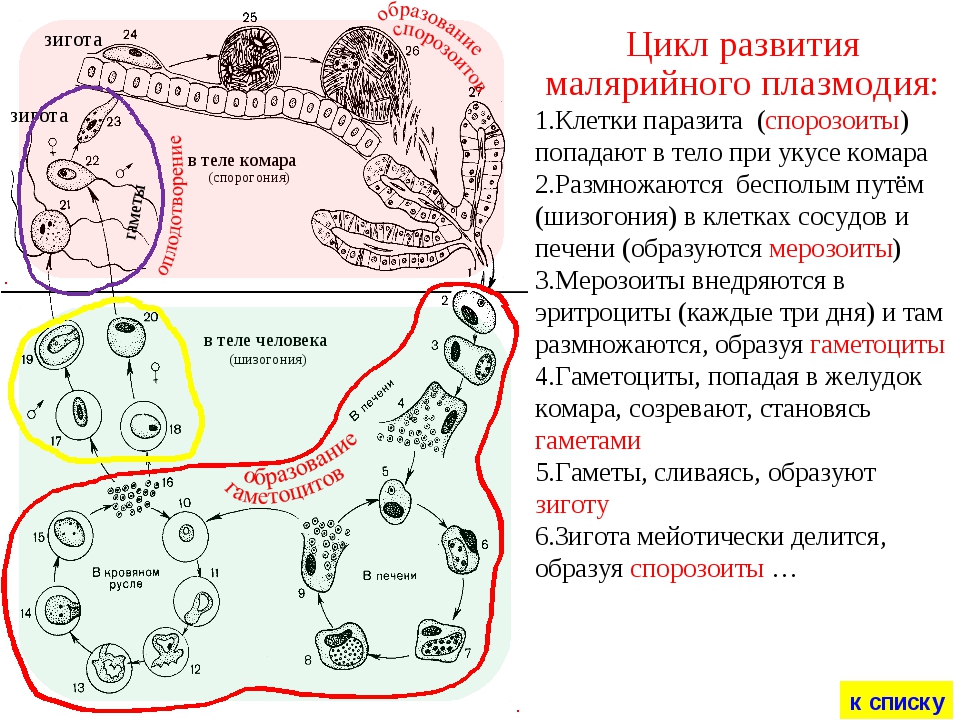 0002123002 00000 п.
0002125641 00000 п.
0002134277 00000 п.
0002136482 00000 п.
0002145297 00000 п.
0002147443 00000 п.
0002155682 00000 п.
0002157801 00000 п.
0002165840 00000 п.
0002167909 00000 п.
0002175415 00000 п.
0002177500 00000 п.
0002182581 00000 п.
00021
0002123002 00000 п.
0002125641 00000 п.
0002134277 00000 п.
0002136482 00000 п.
0002145297 00000 п.
0002147443 00000 п.
0002155682 00000 п.
0002157801 00000 п.
0002165840 00000 п.
0002167909 00000 п.
0002175415 00000 п.
0002177500 00000 п.
0002182581 00000 п.
0002100000 п. 00021
00000 п. 0002199583 00000 п. 0002199955 00000 н. 0002200342 00000 п. 0002200463 00000 п. 0002200617 00000 п. 0002200847 00000 пн 0002201232 00000 н. 0002201353 00000 п. 0002201520 00000 н. 0002201750 00000 п. 0002202137 00000 п. 0002202258 00000 п. 0002202425 00000 п. 0002202579 00000 п. 0002202809 00000 п. 0002203196 00000 п. 0002203317 00000 п. 0002203484 00000 п. 0002203835 00000 п. 0002204222 00000 п. 0002204343 00000 п. 0002204497 00000 п. 0002204664 00000 п. 0002210985 00000 п. 0002213107 00000 п. 0002221049 00000 п. 0002223156 00000 п. 0002231032 00000 п. 0002233143 00000 п. 0002241047 00000 п. 0002243159 00000 п. 0002251035 00000 п. 0002253142 00000 п. 0002260990 00000 п.
0002268177 00000 п.
0002270275 00000 п.
0002278162 00000 п.
0002280274 00000 п.
0002288264 00000 п.
00022 00000 п.
0002298317 00000 п.
0002300437 00000 п.
0002308404 00000 п.
0002310520 00000 п.
0002318516 00000 п.
0002324045 00000 п.
0002326156 00000 п.
0002334137 00000 п.
0002336251 00000 п.
0002344090 00000 п.
0002346181 00000 п.
0002354126 00000 п.
0002356214 00000 п.
0002364171 00000 п.
0002366257 00000 п.
0002374209 00000 п.
0002375981 00000 п.
0002378069 00000 п.
0002386001 00000 п.
0002388097 00000 п.
0002396116 00000 п.
0002398204 00000 п.
0002406217 00000 п.
0002408302 00000 п.
0002416291 00000 п.
0002418387 00000 п.
0002426403 00000 п.
0002426777 00000 п.
0002426874 00000 п.
0002427020 00000 n
0002432374 00000 п.
0002434479 00000 п.
0002442484 00000 п.
0002444575 00000 п.
0002452543 00000 п.
0002454646 00000 п.
0002462657 00000 п.
0002464752 00000 п.
0002472709 00000 п.
0002474784 00000 п.
0002482665 00000 п.
0002260990 00000 п.
0002268177 00000 п.
0002270275 00000 п.
0002278162 00000 п.
0002280274 00000 п.
0002288264 00000 п.
00022 00000 п.
0002298317 00000 п.
0002300437 00000 п.
0002308404 00000 п.
0002310520 00000 п.
0002318516 00000 п.
0002324045 00000 п.
0002326156 00000 п.
0002334137 00000 п.
0002336251 00000 п.
0002344090 00000 п.
0002346181 00000 п.
0002354126 00000 п.
0002356214 00000 п.
0002364171 00000 п.
0002366257 00000 п.
0002374209 00000 п.
0002375981 00000 п.
0002378069 00000 п.
0002386001 00000 п.
0002388097 00000 п.
0002396116 00000 п.
0002398204 00000 п.
0002406217 00000 п.
0002408302 00000 п.
0002416291 00000 п.
0002418387 00000 п.
0002426403 00000 п.
0002426777 00000 п.
0002426874 00000 п.
0002427020 00000 n
0002432374 00000 п.
0002434479 00000 п.
0002442484 00000 п.
0002444575 00000 п.
0002452543 00000 п.
0002454646 00000 п.
0002462657 00000 п.
0002464752 00000 п.
0002472709 00000 п.
0002474784 00000 п.
0002482665 00000 п.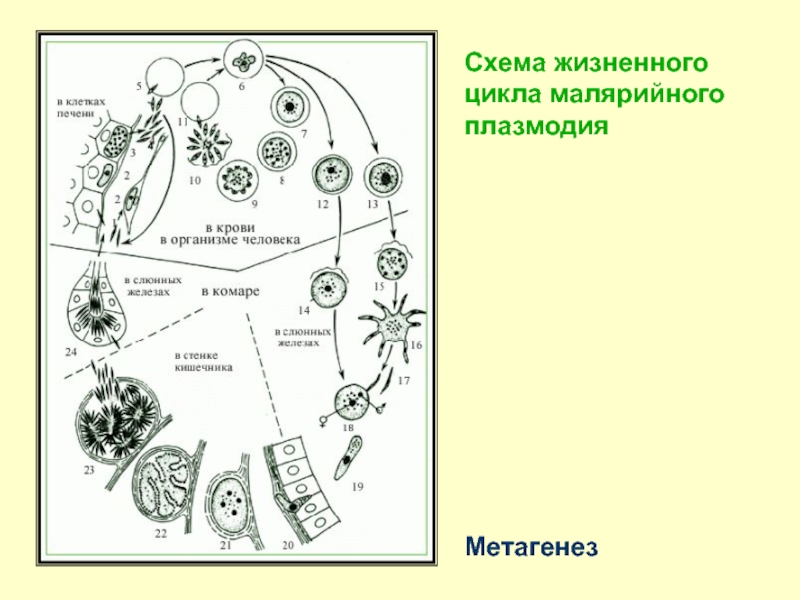 0002487362 00000 п.
0002489444 00000 п.
0002497302 00000 п.
0002499402 00000 п.
0002507339 00000 п.
0002509456 00000 п.
0002517248 00000 п.
0002519352 00000 п.
0002527177 00000 п.
0002529303 00000 п.
0002537189 00000 п.
0002546143 00000 п.
0002548265 00000 н.
0002556102 00000 п.
0002558233 00000 п.
0002566161 00000 п.
0002568285 00000 п.
0002576195 00000 п.
0002578320 00000 н.
0002586209 00000 п.
0002588281 00000 п.
0002596149 00000 n
0002602423 00000 п.
0002604510 00000 п.
0002612427 00000 п.
0002614512 00000 п.
0002622406 00000 п.
0002624504 00000 п.
0002632335 00000 п.
0002634439 00000 п.
0002642432 00000 н.
0002644555 00000 п.
0002652391 00000 п.
0002658539 00000 п.
0002660667 00000 п.
0002668383 00000 п.
0002670490 00000 п.
0002678244 00000 п.
0002680324 00000 п.
0002688081 00000 п.
00026 00000 п.
0002698013 00000 п.
0002700099 00000 н.
0002707969 00000 п.
0002714903 00000 н.
0002716988 00000 н.
0002724668 00000 н.
0002726730 00000 н.
0002487362 00000 п.
0002489444 00000 п.
0002497302 00000 п.
0002499402 00000 п.
0002507339 00000 п.
0002509456 00000 п.
0002517248 00000 п.
0002519352 00000 п.
0002527177 00000 п.
0002529303 00000 п.
0002537189 00000 п.
0002546143 00000 п.
0002548265 00000 н.
0002556102 00000 п.
0002558233 00000 п.
0002566161 00000 п.
0002568285 00000 п.
0002576195 00000 п.
0002578320 00000 н.
0002586209 00000 п.
0002588281 00000 п.
0002596149 00000 n
0002602423 00000 п.
0002604510 00000 п.
0002612427 00000 п.
0002614512 00000 п.
0002622406 00000 п.
0002624504 00000 п.
0002632335 00000 п.
0002634439 00000 п.
0002642432 00000 н.
0002644555 00000 п.
0002652391 00000 п.
0002658539 00000 п.
0002660667 00000 п.
0002668383 00000 п.
0002670490 00000 п.
0002678244 00000 п.
0002680324 00000 п.
0002688081 00000 п.
00026 00000 п.
0002698013 00000 п.
0002700099 00000 н.
0002707969 00000 п.
0002714903 00000 н.
0002716988 00000 н.
0002724668 00000 н.
0002726730 00000 н. 0002734533 00000 н.
0002736627 00000 н.
0002744419 00000 н.
0002746491 00000 п.
0002754267 00000 п.
0002756321 00000 п.
0002763925 00000 н.
0002769542 00000 п.
0002771610 00000 н.
0002779274 00000 н.
0002781336 00000 н.
0002789021 00000 н.
00027 00000 n
0002798847 00000 н.
0002800936 00000 п.
0002808750 00000 п.
0002810825 00000 п.
0002818704 00000 п.
0002825730 00000 п.
0002827855 00000 н.
0002835716 00000 п.
0002837834 00000 п.
0002845588 00000 п.
0002847712 00000 п.
0002855397 00000 п.
0002857497 00000 п.
0002865276 00000 п.
0002867383 00000 п.
0002875153 00000 п.
0002880192 00000 п.
0002882294 00000 п.
00028 00000 п.
00028
0002734533 00000 н.
0002736627 00000 н.
0002744419 00000 н.
0002746491 00000 п.
0002754267 00000 п.
0002756321 00000 п.
0002763925 00000 н.
0002769542 00000 п.
0002771610 00000 н.
0002779274 00000 н.
0002781336 00000 н.
0002789021 00000 н.
00027 00000 n
0002798847 00000 н.
0002800936 00000 п.
0002808750 00000 п.
0002810825 00000 п.
0002818704 00000 п.
0002825730 00000 п.
0002827855 00000 н.
0002835716 00000 п.
0002837834 00000 п.
0002845588 00000 п.
0002847712 00000 п.
0002855397 00000 п.
0002857497 00000 п.
0002865276 00000 п.
0002867383 00000 п.
0002875153 00000 п.
0002880192 00000 п.
0002882294 00000 п.
00028 00000 п.
0002800000 п. 0002899926 00000 н. 0002
9 00000 н. 0002
4 00000 н. 00023 00000 п. 00029 00000 п. 00028 00000 н.
0002929484 00000 н.
0002934160 00000 п.
0002936260 00000 п.
0002943873 00000 п.
0002945985 00000 п.
0002953738 00000 п.
0002955855 00000 п.
0002963613 00000 п.
0002965727 00000 н.
0002973475 00000 п. 0002975576 00000 п.
0002983026 00000 п.
0002985919 00000 п.
00029
0002975576 00000 п.
0002983026 00000 п.
0002985919 00000 п.
00029
 0003183942 00000 п.
0003186045 00000 п.
0003193977 00000 п.
0003194131 00000 п.
0003194518 00000 п.
0003194615 00000 п.
0003194761 00000 п.
0003199687 00000 п.
0003199917 00000 н.
0003200304 00000 п.
0003200425 00000 п.
0003200579 00000 п.
0003200808 00000 п.
0003201195 00000 п.
0003201316 00000 н.
0003201462 00000 н.
0003203669 00000 п.
0003212513 00000 п.
0003214707 00000 н.
0003223404 00000 п.
0003225596 00000 п.
0003234336 00000 п.
0003236522 00000 пн
0003245158 00000 п.
0003251477 00000 н.
0003253673 00000 п.
0003262346 00000 п.
0003264541 00000 п.
0003273382 00000 п.
0003275582 00000 п.
0003284274 00000 н.
0003286477 00000 н.
0003295203 00000 п.
0003297416 00000 п.
0003306146 00000 п.
0003313898 00000 п.
0003316101 00000 п.
0003324815 00000 н.
0003327024 00000 н.
0003335711 00000 п.
0003337920 00000 н.
0003346608 00000 п.
0003348822 00000 п.
0003357490 00000 н.
0003359703 00000 п.
0003368449 00000 п.
0003372986 00000 п.
0003375203 00000 п.
0003183942 00000 п.
0003186045 00000 п.
0003193977 00000 п.
0003194131 00000 п.
0003194518 00000 п.
0003194615 00000 п.
0003194761 00000 п.
0003199687 00000 п.
0003199917 00000 н.
0003200304 00000 п.
0003200425 00000 п.
0003200579 00000 п.
0003200808 00000 п.
0003201195 00000 п.
0003201316 00000 н.
0003201462 00000 н.
0003203669 00000 п.
0003212513 00000 п.
0003214707 00000 н.
0003223404 00000 п.
0003225596 00000 п.
0003234336 00000 п.
0003236522 00000 пн
0003245158 00000 п.
0003251477 00000 н.
0003253673 00000 п.
0003262346 00000 п.
0003264541 00000 п.
0003273382 00000 п.
0003275582 00000 п.
0003284274 00000 н.
0003286477 00000 н.
0003295203 00000 п.
0003297416 00000 п.
0003306146 00000 п.
0003313898 00000 п.
0003316101 00000 п.
0003324815 00000 н.
0003327024 00000 н.
0003335711 00000 п.
0003337920 00000 н.
0003346608 00000 п.
0003348822 00000 п.
0003357490 00000 н.
0003359703 00000 п.
0003368449 00000 п.
0003372986 00000 п.
0003375203 00000 п. 0003383911 00000 п.
0003386117 00000 п.
0003394813 00000 н.
0003397020 00000 п.
0003405634 00000 п.
0003407832 00000 п.
0003416506 00000 п.
0003418705 00000 п.
0003427391 00000 п.
0003432227 00000 п.
0003434434 00000 п.
0003443050 00000 п.
0003445265 00000 п.
0003453992 00000 п.
0003456208 00000 п.
0003465026 00000 п.
0003467232 00000 п.
0003475912 00000 п.
0003478112 00000 п.
0003486823 00000 п.
00034
0003383911 00000 п.
0003386117 00000 п.
0003394813 00000 н.
0003397020 00000 п.
0003405634 00000 п.
0003407832 00000 п.
0003416506 00000 п.
0003418705 00000 п.
0003427391 00000 п.
0003432227 00000 п.
0003434434 00000 п.
0003443050 00000 п.
0003445265 00000 п.
0003453992 00000 п.
0003456208 00000 п.
0003465026 00000 п.
0003467232 00000 п.
0003475912 00000 п.
0003478112 00000 п.
0003486823 00000 п.
00034 0003709633 00000 п.
0003723100 00000 п.
0003736385 00000 п.
0003749481 00000 п.
0003762170 00000 п.
0003774797 00000 п.
0003778344 00000 п.
0003778731 00000 п.
0003778828 00000 п.
0003778982 00000 п.
00037
0003709633 00000 п.
0003723100 00000 п.
0003736385 00000 п.
0003749481 00000 п.
0003762170 00000 п.
0003774797 00000 п.
0003778344 00000 п.
0003778731 00000 п.
0003778828 00000 п.
0003778982 00000 п.
0003700000 п. 0003803779 00000 п. 0003816322 00000 п. 0003828851 00000 п. 0003840773 00000 п. 0003853168 00000 п. 0003865153 00000 п. 0003875383 00000 п. 0003884222 00000 п. 0003888844 00000 п. 0003893991 00000 п. 0003
3 00000 н. 00031 00000 п. 00034 00000 п.
0003930179 00000 п.
0003939283 00000 п.
0003944149 00000 п.
0003949171 00000 п.
0003957023 00000 п.
0003965248 00000 п.
0003969886 00000 п.
0003976539 00000 п.
0003984241 00000 п.
0003988853 00000 п.
0004001878 00000 п.
0004014683 00000 п.
0004021572 00000 п.
0004049775 00000 п.
0004062321 00000 п.
0004067949 00000 п.
0004069687 00000 п.
0004074989 00000 п.
0004080128 00000 п.
0004086270 00000 п.
00040 00000 п.
0004097125 00000 п.
0004105793 00000 п.
0004111773 00000 п.
0004118661 00000 п. 0004124059 00000 п.
0004130871 00000 п.
0004137925 00000 п.
0004142871 00000 п.
0004147863 00000 п.
0004155448 00000 н.
0004169826 00000 п.
0004175360 00000 п.
0004180577 00000 п.
0004185136 00000 п.
00041 00000 n
0004199465 00000 п.
0004201914 00000 п.
0004202144 00000 п.
0004202531 00000 п.
0004202918 00000 н.
0004203321 00000 п.
0004203493 00000 п.
0004203647 00000 п.
0004207868 00000 п.
0004213265 00000 п.
0004220296 00000 п.
0004223936 00000 п.
0004224011 00000 п.
0004224136 00000 п.
0004224431 00000 п.
0004224506 00000 п.
0004224802 00000 п.
0004224877 00000 п.
0004225174 00000 п.
00042
0004124059 00000 п.
0004130871 00000 п.
0004137925 00000 п.
0004142871 00000 п.
0004147863 00000 п.
0004155448 00000 н.
0004169826 00000 п.
0004175360 00000 п.
0004180577 00000 п.
0004185136 00000 п.
00041 00000 n
0004199465 00000 п.
0004201914 00000 п.
0004202144 00000 п.
0004202531 00000 п.
0004202918 00000 н.
0004203321 00000 п.
0004203493 00000 п.
0004203647 00000 п.
0004207868 00000 п.
0004213265 00000 п.
0004220296 00000 п.
0004223936 00000 п.
0004224011 00000 п.
0004224136 00000 п.
0004224431 00000 п.
0004224506 00000 п.
0004224802 00000 п.
0004224877 00000 п.
0004225174 00000 п.
00042
00000 п.
00042 00000 п.
00042 00000 п.
0004292868 00000 п.
0004293053 00000 п.
0004293238 00000 п.
0004293946 00000 п.
0004294140 00000 п.
0004294325 00000 п.
0004295033 00000 п.
0004295239 00000 п.
0004295441 00000 п.
0004296147 00000 п.
0004297202 00000 п.
0004297908 00000 н.
0004298118 00000 п.
0004298329 00000 п.
0004299035 00000 п.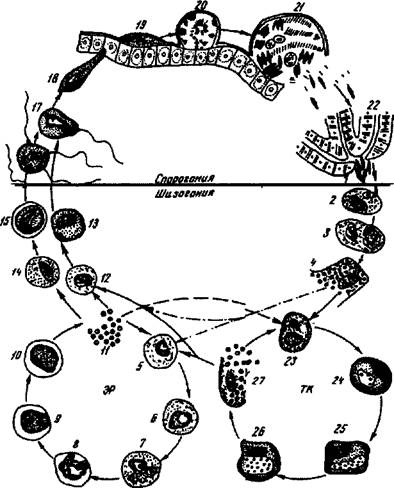 0004299741 00000 п.
0004300231 00000 п.
0004300939 00000 п.
0004301144 00000 п.
0004301329 00000 п.
0004302037 00000 п.
0004302745 00000 п.
0004303799 00000 п.
0004304507 00000 п.
0004304692 00000 п.
0004305402 00000 п.
0004305605 00000 п.
0004305799 00000 п.
0004306505 00000 п.
0004306994 00000 п.
0004307196 00000 п.
0004307402 00000 п.
0004308107 00000 п.
0004308332 00000 п.
0004308541 00000 п.
0004309247 00000 п.
0004309449 00000 п.
0004309634 00000 п.
0004310342 00000 п.
0004310544 00000 п.
0004311563 00000 п.
0004311748 00000 н.
0004312456 00000 п.
0004313164 00000 п.
0004313366 00000 п.
0004314074 00000 п.
0004314441 00000 п.
0004315149 00000 п.
0004315351 00000 п.
0004315536 00000 п.
0004316248 00000 п.
0004316467 00000 п.
0004317173 00000 п.
0004317391 00000 п.
0004317603 00000 п.
0004318309 00000 п.
0004319022 00000 п.
0004319224 00000 п.
0004319418 00000 п.
0004319628 00000 п.
0004320336 00000 п.
0004320546 00000 п.
0004320731 00000 п.
0004299741 00000 п.
0004300231 00000 п.
0004300939 00000 п.
0004301144 00000 п.
0004301329 00000 п.
0004302037 00000 п.
0004302745 00000 п.
0004303799 00000 п.
0004304507 00000 п.
0004304692 00000 п.
0004305402 00000 п.
0004305605 00000 п.
0004305799 00000 п.
0004306505 00000 п.
0004306994 00000 п.
0004307196 00000 п.
0004307402 00000 п.
0004308107 00000 п.
0004308332 00000 п.
0004308541 00000 п.
0004309247 00000 п.
0004309449 00000 п.
0004309634 00000 п.
0004310342 00000 п.
0004310544 00000 п.
0004311563 00000 п.
0004311748 00000 н.
0004312456 00000 п.
0004313164 00000 п.
0004313366 00000 п.
0004314074 00000 п.
0004314441 00000 п.
0004315149 00000 п.
0004315351 00000 п.
0004315536 00000 п.
0004316248 00000 п.
0004316467 00000 п.
0004317173 00000 п.
0004317391 00000 п.
0004317603 00000 п.
0004318309 00000 п.
0004319022 00000 п.
0004319224 00000 п.
0004319418 00000 п.
0004319628 00000 п.
0004320336 00000 п.
0004320546 00000 п.
0004320731 00000 п. 0004321439 00000 п.
0004321625 00000 н.
0004322341 00000 п.
0004322527 00000 н.
0004322713 00000 н.
0004322916 00000 н.
0004323625 00000 п.
0004323835 00000 п.
0004324044 00000 п.
0004324750 00000 п.
0004324959 00000 н.
0004325170 00000 н.
0004325924 00000 н.
0004326630 00000 п.
0004326838 00000 п.
0004327033 00000 п.
0004327739 00000 п.
0004327951 00000 п.
0004328160 00000 п.
0004328869 00000 п.
0004329064 00000 н.
0004329259 00000 п.
0004329968 00000 н.
0004330235 00000 п.
0004332266 00000 п.
0004332452 00000 п.
0004332661 00000 п.
0004333370 00000 п.
0004333577 00000 п.
0004333780 00000 п.
0004334484 00000 п.
0004335188 00000 п.
0004335894 00000 п.
0004336598 00000 п.
0004336966 00000 п.
0004337670 00000 п.
0004338374 00000 п.
0004339078 00000 н.
0004339782 00000 п.
0004340487 00000 п.
0004341192 00000 п.
0004341897 00000 п.
0004342638 00000 п.
0004343343 00000 п.
0004344048 00000 п.
0004344752 00000 п.
0004345456 00000 п.
0004346160 00000 п.
0004321439 00000 п.
0004321625 00000 н.
0004322341 00000 п.
0004322527 00000 н.
0004322713 00000 н.
0004322916 00000 н.
0004323625 00000 п.
0004323835 00000 п.
0004324044 00000 п.
0004324750 00000 п.
0004324959 00000 н.
0004325170 00000 н.
0004325924 00000 н.
0004326630 00000 п.
0004326838 00000 п.
0004327033 00000 п.
0004327739 00000 п.
0004327951 00000 п.
0004328160 00000 п.
0004328869 00000 п.
0004329064 00000 н.
0004329259 00000 п.
0004329968 00000 н.
0004330235 00000 п.
0004332266 00000 п.
0004332452 00000 п.
0004332661 00000 п.
0004333370 00000 п.
0004333577 00000 п.
0004333780 00000 п.
0004334484 00000 п.
0004335188 00000 п.
0004335894 00000 п.
0004336598 00000 п.
0004336966 00000 п.
0004337670 00000 п.
0004338374 00000 п.
0004339078 00000 н.
0004339782 00000 п.
0004340487 00000 п.
0004341192 00000 п.
0004341897 00000 п.
0004342638 00000 п.
0004343343 00000 п.
0004344048 00000 п.
0004344752 00000 п.
0004345456 00000 п.
0004346160 00000 п.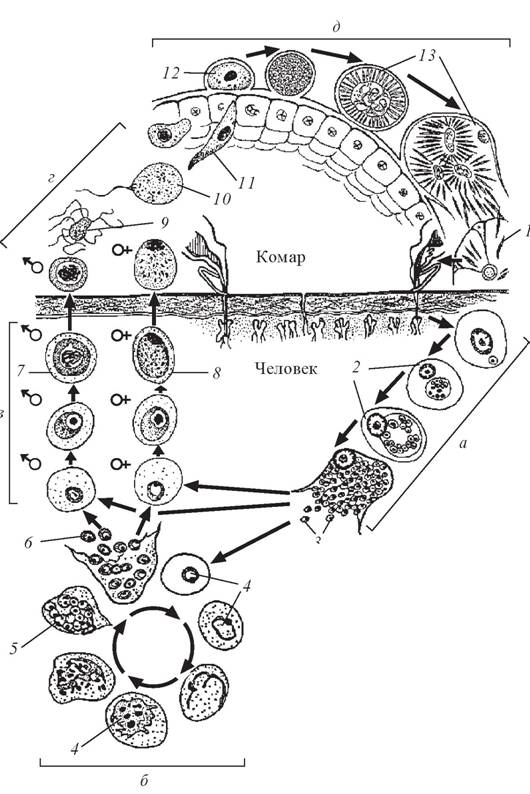 0004346864 00000 п.
0004347568 00000 п.
0004348136 00000 п.
0004348840 00000 п.
0004349544 00000 п.
0004350248 00000 п.
0004350952 00000 п.
0004351656 00000 п.
0004352433 00000 п.
0004353139 00000 п.
0004353845 00000 п.
0004354549 00000 п.
0004355255 00000 п.
0004355967 00000 п.
0004356673 00000 п.
0004357378 00000 п.
0004358083 00000 п.
0004358808 00000 п.
0004359513 00000 п.
0004360218 00000 п.
0004360398 00000 п.
0004361101 00000 п.
0004361805 00000 п.
0004362027 00000 н.
0004362733 00000 п.
0004363437 00000 п.
0004364195 00000 п.
0004364899 00000 н.
0004365603 00000 п.
0004366307 00000 п.
0004367011 00000 п.
0004367715 00000 п.
0004368419 00000 п.
0004369123 00000 п.
0004371215 00000 п.
0004371922 00000 п.
0004372628 00000 п.
0004373332 00000 п.
0004374036 00000 п.
0004374410 00000 п.
0004375115 00000 п.
0004375821 00000 п.
0004376526 00000 п.
0004377231 00000 п.
0004377935 00000 п.
0004378639 00000 п.
0004379343 00000 п.
0004380049 00000 п.
0004346864 00000 п.
0004347568 00000 п.
0004348136 00000 п.
0004348840 00000 п.
0004349544 00000 п.
0004350248 00000 п.
0004350952 00000 п.
0004351656 00000 п.
0004352433 00000 п.
0004353139 00000 п.
0004353845 00000 п.
0004354549 00000 п.
0004355255 00000 п.
0004355967 00000 п.
0004356673 00000 п.
0004357378 00000 п.
0004358083 00000 п.
0004358808 00000 п.
0004359513 00000 п.
0004360218 00000 п.
0004360398 00000 п.
0004361101 00000 п.
0004361805 00000 п.
0004362027 00000 н.
0004362733 00000 п.
0004363437 00000 п.
0004364195 00000 п.
0004364899 00000 н.
0004365603 00000 п.
0004366307 00000 п.
0004367011 00000 п.
0004367715 00000 п.
0004368419 00000 п.
0004369123 00000 п.
0004371215 00000 п.
0004371922 00000 п.
0004372628 00000 п.
0004373332 00000 п.
0004374036 00000 п.
0004374410 00000 п.
0004375115 00000 п.
0004375821 00000 п.
0004376526 00000 п.
0004377231 00000 п.
0004377935 00000 п.
0004378639 00000 п.
0004379343 00000 п.
0004380049 00000 п. 0004380789 00000 п.
0004381493 00000 п.
0004382197 00000 п.
0004382901 00000 п.
0004383605 00000 п.
0004384311 00000 п.
0004385017 00000 п.
0004385721 00000 п.
0004386427 00000 н.
0004387133 00000 п.
0004387837 00000 п.
0004388212 00000 п.
0004388781 00000 п.
0004389485 00000 п.
00043 00000 п.
00043
0004380789 00000 п.
0004381493 00000 п.
0004382197 00000 п.
0004382901 00000 п.
0004383605 00000 п.
0004384311 00000 п.
0004385017 00000 п.
0004385721 00000 п.
0004386427 00000 н.
0004387133 00000 п.
0004387837 00000 п.
0004388212 00000 п.
0004388781 00000 п.
0004389485 00000 п.
00043 00000 п.
00043
00000 п. 00043 00000 п. 00043
00000 п.
0004393050 00000 п.
0004393756 00000 п.
0004394463 00000 н.
0004395169 00000 п.
0004395874 00000 п.
0004396579 00000 п.
0004397283 00000 п.
0004397992 00000 н.
0004398735 00000 п.
0004398949 00000 н.
0004399652 00000 н.
0004400355 00000 п.
0004400550 00000 п.
0004401254 00000 п.
0004401958 00000 п.
0004402664 00000 п.
0004403370 00000 п.
0004404074 00000 п.
0004404778 00000 п.
0004405487 00000 п.
0004406191 00000 п.
0004406404 00000 п.
0004407107 00000 п.
0004407810 00000 п.
0004408513 00000 п.
0004409216 00000 п.
0004409919 00000 п.
0004410105 00000 п.
0004410808 00000 п.
0004411511 00000 п. 0004412711 00000 п.
0004413414 00000 п.
0004414117 00000 п.
0004414820 00000 п.
0004415523 00000 п.
0004416226 00000 п.
0004416930 00000 п.
0004417633 00000 п.
0004418336 00000 п.
0004419039 00000 п.
0004419743 00000 п.
0004420446 00000 н.
0004420649 00000 п.
0004421352 00000 п.
0004422057 00000 п.
0004422762 00000 п.
0004423489 00000 п.
0004424192 00000 п.
0004424895 00000 п.
0004425599 00000 п.
0004426302 00000 п.
0004427005 00000 п.
0004427708 00000 н.
0004428411 00000 п.
0004429141 00000 п.
0004429347 00000 п.
0004430050 00000 п.
0004430754 00000 п.
0004431457 00000 п.
0004431660 00000 п.
0004432363 00000 п.
0004433066 00000 п.
0004433769 00000 п.
0004434495 00000 п.
0004435198 00000 п.
0004435401 00000 п.
0004436170 00000 п.
0004436913 00000 п.
0004437616 00000 п.
0004438319 00000 п.
0004439022 00000 н.
0004439725 00000 п.
0004440428 00000 п.
0004441131 00000 п.
0004441834 00000 п.
0004442566 00000 н.
0004443269 00000 п.
0004443972 00000 н.
0004412711 00000 п.
0004413414 00000 п.
0004414117 00000 п.
0004414820 00000 п.
0004415523 00000 п.
0004416226 00000 п.
0004416930 00000 п.
0004417633 00000 п.
0004418336 00000 п.
0004419039 00000 п.
0004419743 00000 п.
0004420446 00000 н.
0004420649 00000 п.
0004421352 00000 п.
0004422057 00000 п.
0004422762 00000 п.
0004423489 00000 п.
0004424192 00000 п.
0004424895 00000 п.
0004425599 00000 п.
0004426302 00000 п.
0004427005 00000 п.
0004427708 00000 н.
0004428411 00000 п.
0004429141 00000 п.
0004429347 00000 п.
0004430050 00000 п.
0004430754 00000 п.
0004431457 00000 п.
0004431660 00000 п.
0004432363 00000 п.
0004433066 00000 п.
0004433769 00000 п.
0004434495 00000 п.
0004435198 00000 п.
0004435401 00000 п.
0004436170 00000 п.
0004436913 00000 п.
0004437616 00000 п.
0004438319 00000 п.
0004439022 00000 н.
0004439725 00000 п.
0004440428 00000 п.
0004441131 00000 п.
0004441834 00000 п.
0004442566 00000 н.
0004443269 00000 п.
0004443972 00000 н. 0004444675 00000 п.
0004445378 00000 п.
0004446101 00000 п.
0004446804 00000 п.
0004447507 00000 п.
0004447719 00000 п.
0004447930 00000 п.
0004448138 00000 н.
0004448865 00000 н.
0004449568 00000 п.
0004450297 00000 п.
0004450477 00000 п.
0004450690 00000 п.
0004451393 00000 п.
0004452096 00000 п.
0004452822 00000 п.
0004453525 00000 п.
0004454229 00000 п.
0004454934 00000 п.
0004455137 00000 п.
0004456066 00000 п.
0004458691 00000 п.
0004459395 00000 п.
0004460918 00000 п.
0004461891 00000 п.
0004462678 00000 п.
0004463383 00000 п.
0004465909 00000 п.
0004466257 00000 п.
0004467579 00000 н.
0004467945 00000 п.
0004468914 00000 п.
0004469631 00000 п.
0004470360 00000 п.
0004477427 00000 п.
0004478381 00000 п.
0004479102 00000 п.
0004481229 00000 п.
0004481695 00000 п.
0004484056 00000 п.
0004484765 00000 п.
0004485520 00000 п.
0004486019 00000 п.
0004486731 00000 п.
0004487067 00000 п.
0004488152 00000 п.
0004489017 00000 н.
00044
0004444675 00000 п.
0004445378 00000 п.
0004446101 00000 п.
0004446804 00000 п.
0004447507 00000 п.
0004447719 00000 п.
0004447930 00000 п.
0004448138 00000 н.
0004448865 00000 н.
0004449568 00000 п.
0004450297 00000 п.
0004450477 00000 п.
0004450690 00000 п.
0004451393 00000 п.
0004452096 00000 п.
0004452822 00000 п.
0004453525 00000 п.
0004454229 00000 п.
0004454934 00000 п.
0004455137 00000 п.
0004456066 00000 п.
0004458691 00000 п.
0004459395 00000 п.
0004460918 00000 п.
0004461891 00000 п.
0004462678 00000 п.
0004463383 00000 п.
0004465909 00000 п.
0004466257 00000 п.
0004467579 00000 н.
0004467945 00000 п.
0004468914 00000 п.
0004469631 00000 п.
0004470360 00000 п.
0004477427 00000 п.
0004478381 00000 п.
0004479102 00000 п.
0004481229 00000 п.
0004481695 00000 п.
0004484056 00000 п.
0004484765 00000 п.
0004485520 00000 п.
0004486019 00000 п.
0004486731 00000 п.
0004487067 00000 п.
0004488152 00000 п.
0004489017 00000 н.
00044
 00044 00000 п.
00044 00000 н.
00044 00000 п.
00044 00000 п.
0004493019 00000 п.
0004493740 00000 п.
0004494455 00000 п.
0004495163 00000 п.
0004496824 00000 н.
0004497584 00000 п.
0004498339 00000 п.
0004501074 00000 п.
0004503763 00000 п.
0004504987 00000 п.
0004505696 00000 п.
0004506457 00000 п.
0004507212 00000 п.
0004509411 00000 п.
0004511761 00000 п.
0004512036 00000 п.
0004514957 00000 п.
0004515718 00000 п.
0004516475 00000 п.
0004516696 00000 п.
0004517400 00000 п.
0004517621 00000 п.
0004518326 00000 п.
0004520669 00000 п.
0004522912 00000 п.
0004523121 00000 п.
0004523838 00000 п.
0004526445 00000 п.
0004527154 00000 п.
0004527916 00000 n
0004528672 00000 п.
0004528894 00000 п.
0004529110 00000 п.
0004529815 00000 п.
0004530710 00000 п.
0004533037 00000 п.
0004533399 00000 п.
0004534556 00000 п.
0004535262 00000 п.
0004535966 00000 п.
0004536727 00000 п.
0004537483 00000 п.
0004537711 00000 п.
0004537936 00000 п.
0004538639 00000 п.
00044 00000 п.
00044 00000 н.
00044 00000 п.
00044 00000 п.
0004493019 00000 п.
0004493740 00000 п.
0004494455 00000 п.
0004495163 00000 п.
0004496824 00000 н.
0004497584 00000 п.
0004498339 00000 п.
0004501074 00000 п.
0004503763 00000 п.
0004504987 00000 п.
0004505696 00000 п.
0004506457 00000 п.
0004507212 00000 п.
0004509411 00000 п.
0004511761 00000 п.
0004512036 00000 п.
0004514957 00000 п.
0004515718 00000 п.
0004516475 00000 п.
0004516696 00000 п.
0004517400 00000 п.
0004517621 00000 п.
0004518326 00000 п.
0004520669 00000 п.
0004522912 00000 п.
0004523121 00000 п.
0004523838 00000 п.
0004526445 00000 п.
0004527154 00000 п.
0004527916 00000 n
0004528672 00000 п.
0004528894 00000 п.
0004529110 00000 п.
0004529815 00000 п.
0004530710 00000 п.
0004533037 00000 п.
0004533399 00000 п.
0004534556 00000 п.
0004535262 00000 п.
0004535966 00000 п.
0004536727 00000 п.
0004537483 00000 п.
0004537711 00000 п.
0004537936 00000 п.
0004538639 00000 п.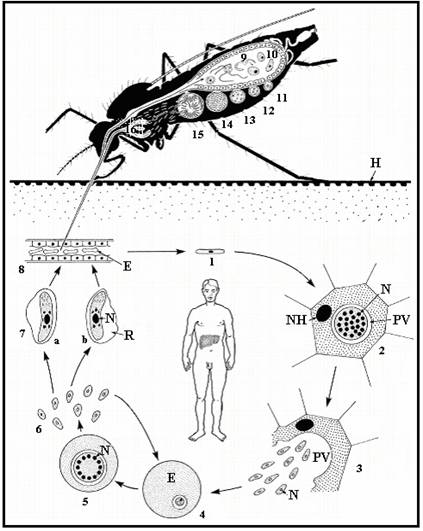 0004539344 00000 п.
0004541535 00000 п.
0004543863 00000 п.
0004544905 00000 п.
0004545665 00000 п.
0004546421 00000 п.
0004546642 00000 п.
0004547345 00000 п.
0004547721 00000 п.
0004550096 00000 п.
0004550857 00000 п.
0004551614 00000 п.
0004552319 00000 п.
0004552543 00000 п.
0004553247 00000 п.
0004553952 00000 п.
0004555838 00000 п.
0004557264 00000 п.
0004560120 00000 п.
0004560855 00000 п.
0004561607 00000 п.
0004561787 00000 п.
0004562010 00000 п.
0004562241 00000 п.
0004562471 00000 п.
0004562674 00000 п.
0004563379 00000 п.
0004563889 00000 п.
0004565837 00000 п.
0004566581 00000 п.
0004567301 00000 п.
0004567614 00000 п.
0004569530 00000 п.
0004569962 00000 н.
0004570878 00000 п.
0004571534 00000 п.
0004571904 00000 п.
0004572625 00000 п.
0004573338 00000 п.
0004574044 00000 п.
0004574770 00000 п.
0004575478 00000 п.
0004576010 00000 п.
0004576747 00000 п.
0004578275 00000 п.
0004579917 00000 п.
0004580203 00000 п.
0004580912 00000 п.
0004539344 00000 п.
0004541535 00000 п.
0004543863 00000 п.
0004544905 00000 п.
0004545665 00000 п.
0004546421 00000 п.
0004546642 00000 п.
0004547345 00000 п.
0004547721 00000 п.
0004550096 00000 п.
0004550857 00000 п.
0004551614 00000 п.
0004552319 00000 п.
0004552543 00000 п.
0004553247 00000 п.
0004553952 00000 п.
0004555838 00000 п.
0004557264 00000 п.
0004560120 00000 п.
0004560855 00000 п.
0004561607 00000 п.
0004561787 00000 п.
0004562010 00000 п.
0004562241 00000 п.
0004562471 00000 п.
0004562674 00000 п.
0004563379 00000 п.
0004563889 00000 п.
0004565837 00000 п.
0004566581 00000 п.
0004567301 00000 п.
0004567614 00000 п.
0004569530 00000 п.
0004569962 00000 н.
0004570878 00000 п.
0004571534 00000 п.
0004571904 00000 п.
0004572625 00000 п.
0004573338 00000 п.
0004574044 00000 п.
0004574770 00000 п.
0004575478 00000 п.
0004576010 00000 п.
0004576747 00000 п.
0004578275 00000 п.
0004579917 00000 п.
0004580203 00000 п.
0004580912 00000 п. 0004581834 00000 п.
0004582625 00000 п.
0004583334 00000 п.
0004584531 00000 п.
0004585198 00000 п.
0004585911 00000 п.
0004586220 00000 п.
0004587223 00000 п.
0004587468 00000 п.
0004589855 00000 п.
00045 00000 п.
00045 00000 п.
0004593068 00000 п.
0004593561 00000 п.
0004593872 00000 п.
0004594098 00000 п.
0004594802 00000 п.
0004595050 00000 п.
0004596738 00000 п.
0004597522 00000 п.
0004599359 00000 н.
0004600203 00000 п.
0004600907 00000 п.
0004603304 00000 п.
0004604019 00000 п.
0004604728 00000 п.
0004605389 00000 п.
0004606093 00000 п.
0004606797 00000 п.
0004607060 00000 п.
0004610274 00000 п.
0004610859 00000 п.
0004611419 00000 п.
0004611633 00000 п.
0004612346 00000 п.
0004612931 00000 п.
0004613516 00000 п.
0004614224 00000 н.
0004614812 00000 п.
0004615202 00000 н.
0004615934 00000 п.
0004616508 00000 п.
0004617217 00000 п.
0004617924 00000 н.
0004618128 00000 п.
0004618331 00000 п.
0004618914 00000 п.
0004619617 00000 п.
0004581834 00000 п.
0004582625 00000 п.
0004583334 00000 п.
0004584531 00000 п.
0004585198 00000 п.
0004585911 00000 п.
0004586220 00000 п.
0004587223 00000 п.
0004587468 00000 п.
0004589855 00000 п.
00045 00000 п.
00045 00000 п.
0004593068 00000 п.
0004593561 00000 п.
0004593872 00000 п.
0004594098 00000 п.
0004594802 00000 п.
0004595050 00000 п.
0004596738 00000 п.
0004597522 00000 п.
0004599359 00000 н.
0004600203 00000 п.
0004600907 00000 п.
0004603304 00000 п.
0004604019 00000 п.
0004604728 00000 п.
0004605389 00000 п.
0004606093 00000 п.
0004606797 00000 п.
0004607060 00000 п.
0004610274 00000 п.
0004610859 00000 п.
0004611419 00000 п.
0004611633 00000 п.
0004612346 00000 п.
0004612931 00000 п.
0004613516 00000 п.
0004614224 00000 н.
0004614812 00000 п.
0004615202 00000 н.
0004615934 00000 п.
0004616508 00000 п.
0004617217 00000 п.
0004617924 00000 н.
0004618128 00000 п.
0004618331 00000 п.
0004618914 00000 п.
0004619617 00000 п.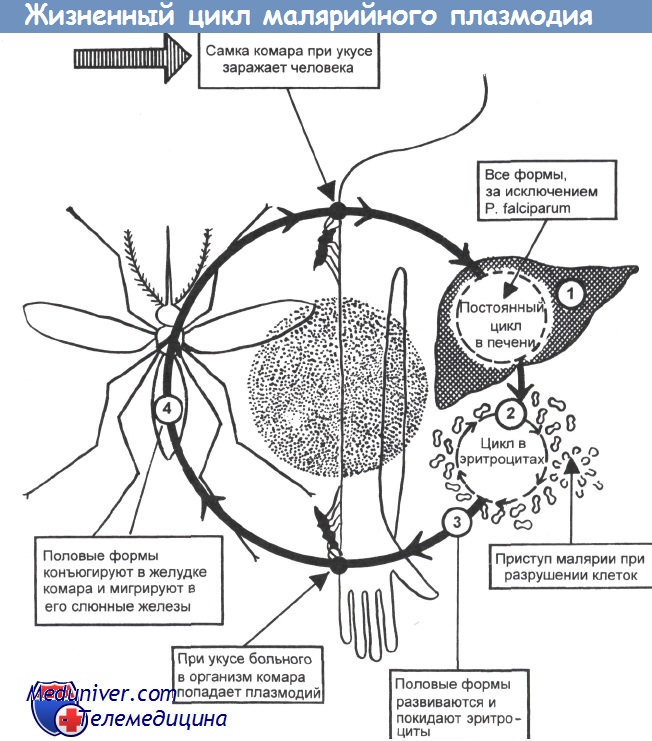 0004620325 00000 н.
0004620511 00000 н.
0004620837 00000 п.
0004621017 00000 п.
0004622407 00000 п.
0004623121 00000 п.
0004623824 00000 п.
0004624552 00000 п.
0004625277 00000 н.
0004625982 00000 п.
0004626687 00000 п.
0004626890 00000 п.
0004627607 00000 п.
0004628316 00000 п.
0004628502 00000 п.
0004628938 00000 п.
0004629644 00000 п.
0004629830 00000 п.
0004630543 00000 п.
0004631249 00000 п.
0004631968 00000 п.
0004632171 00000 п.
0004632876 00000 п.
0004633596 00000 п.
0004633812 00000 п.
0004634536 00000 п.
0004635355 00000 п.
0004636065 00000 п.
0004636278 00000 н.
0004636983 00000 п.
0004637697 00000 п.
0004638404 00000 п.
0004638608 00000 п.
0004638818 00000 п.
0004639004 00000 п.
0004639230 00000 п.
0004641902 00000 н.
0004642105 00000 п.
0004642334 00000 п.
0004642514 00000 п.
0004643251 00000 п.
0004643464 00000 н.
0004643866 00000 п.
0004644255 00000 н.
0004644976 00000 п.
0004645168 00000 п.
0004645889 00000 п.
0004646611 00000 п.
0004620325 00000 н.
0004620511 00000 н.
0004620837 00000 п.
0004621017 00000 п.
0004622407 00000 п.
0004623121 00000 п.
0004623824 00000 п.
0004624552 00000 п.
0004625277 00000 н.
0004625982 00000 п.
0004626687 00000 п.
0004626890 00000 п.
0004627607 00000 п.
0004628316 00000 п.
0004628502 00000 п.
0004628938 00000 п.
0004629644 00000 п.
0004629830 00000 п.
0004630543 00000 п.
0004631249 00000 п.
0004631968 00000 п.
0004632171 00000 п.
0004632876 00000 п.
0004633596 00000 п.
0004633812 00000 п.
0004634536 00000 п.
0004635355 00000 п.
0004636065 00000 п.
0004636278 00000 н.
0004636983 00000 п.
0004637697 00000 п.
0004638404 00000 п.
0004638608 00000 п.
0004638818 00000 п.
0004639004 00000 п.
0004639230 00000 п.
0004641902 00000 н.
0004642105 00000 п.
0004642334 00000 п.
0004642514 00000 п.
0004643251 00000 п.
0004643464 00000 н.
0004643866 00000 п.
0004644255 00000 н.
0004644976 00000 п.
0004645168 00000 п.
0004645889 00000 п.
0004646611 00000 п. 0004646806 00000 п.
0004647510 00000 п.
0004648222 00000 п.
0004648933 00000 п.
0004649119 00000 п.
0004649322 00000 п.
0004649548 00000 н.
0004649740 00000 п.
0004649951 00000 н.
0004650666 00000 п.
0004650894 00000 п.
0004651935 00000 п.
0004652712 00000 н.
0004653479 00000 п.
0004653677 00000 п.
0004654381 00000 п.
0004654567 00000 п.
0004654753 00000 п.
0004655461 00000 п.
0004656187 00000 п.
0004656367 00000 п.
0004656893 00000 п.
0004657605 00000 п.
0004658329 00000 н.
0004659048 00000 н.
0004659794 00000 п.
0004660010 00000 п.
0004660221 00000 п.
0004660924 00000 п.
0004661635 00000 п.
0004662341 00000 п.
0004663050 00000 п.
0004663242 00000 н.
0004663422 00000 п.
0004664131 00000 п.
0004664840 00000 п.
0004665361 00000 п.
0004665547 00000 п.
0004665775 00000 п.
0004665967 00000 п.
0004666159 00000 п.
0004666895 00000 п.
0004667600 00000 п.
0004668313 00000 п.
0004669017 00000 н.
0004669730 00000 п.
0004670456 00000 п.
0004670665 00000 п.
0004646806 00000 п.
0004647510 00000 п.
0004648222 00000 п.
0004648933 00000 п.
0004649119 00000 п.
0004649322 00000 п.
0004649548 00000 н.
0004649740 00000 п.
0004649951 00000 н.
0004650666 00000 п.
0004650894 00000 п.
0004651935 00000 п.
0004652712 00000 н.
0004653479 00000 п.
0004653677 00000 п.
0004654381 00000 п.
0004654567 00000 п.
0004654753 00000 п.
0004655461 00000 п.
0004656187 00000 п.
0004656367 00000 п.
0004656893 00000 п.
0004657605 00000 п.
0004658329 00000 н.
0004659048 00000 н.
0004659794 00000 п.
0004660010 00000 п.
0004660221 00000 п.
0004660924 00000 п.
0004661635 00000 п.
0004662341 00000 п.
0004663050 00000 п.
0004663242 00000 н.
0004663422 00000 п.
0004664131 00000 п.
0004664840 00000 п.
0004665361 00000 п.
0004665547 00000 п.
0004665775 00000 п.
0004665967 00000 п.
0004666159 00000 п.
0004666895 00000 п.
0004667600 00000 п.
0004668313 00000 п.
0004669017 00000 н.
0004669730 00000 п.
0004670456 00000 п.
0004670665 00000 п. 0004670851 00000 п.
0004671569 00000 п.
0004672277 00000 п.
0004672486 00000 п.
0004672942 00000 п.
0004673648 00000 н.
0004674375 00000 п.
0004675098 00000 п.
0004675284 00000 п.
0004675488 00000 п.
0004675691 00000 п.
0004675920 00000 н.
0004676628 00000 п.
0004677341 00000 п.
0004678055 00000 п.
0004678241 00000 п.
0004678427 00000 н.
0004678641 00000 п.
0004678845 00000 п.
0004679048 00000 н.
0004679753 00000 п.
0004679975 00000 н.
0004680155 00000 п.
0004680861 00000 п.
0004681059 00000 п.
0004681631 00000 п.
0004682150 00000 п.
0004682336 00000 п.
0004683041 00000 п.
0004683753 00000 п.
0004684465 00000 п.
0004685171 00000 п.
0004685880 00000 п.
0004686422 00000 п.
0004686889 00000 п.
0004687597 00000 п.
0004688315 00000 п.
0004689175 00000 п.
0004689885 00000 п.
00046 00000 п.
00046
0004670851 00000 п.
0004671569 00000 п.
0004672277 00000 п.
0004672486 00000 п.
0004672942 00000 п.
0004673648 00000 н.
0004674375 00000 п.
0004675098 00000 п.
0004675284 00000 п.
0004675488 00000 п.
0004675691 00000 п.
0004675920 00000 н.
0004676628 00000 п.
0004677341 00000 п.
0004678055 00000 п.
0004678241 00000 п.
0004678427 00000 н.
0004678641 00000 п.
0004678845 00000 п.
0004679048 00000 н.
0004679753 00000 п.
0004679975 00000 н.
0004680155 00000 п.
0004680861 00000 п.
0004681059 00000 п.
0004681631 00000 п.
0004682150 00000 п.
0004682336 00000 п.
0004683041 00000 п.
0004683753 00000 п.
0004684465 00000 п.
0004685171 00000 п.
0004685880 00000 п.
0004686422 00000 п.
0004686889 00000 п.
0004687597 00000 п.
0004688315 00000 п.
0004689175 00000 п.
0004689885 00000 п.
00046 00000 п.
00046 00000 п.
00046 00000 n
0004692710 00000 п.
0004692913 00000 н.
0004693617 00000 п.
0004694320 00000 п.
0004694524 00000 п.
0004695228 00000 п. 0004697943 00000 п.
0004698151 00000 п.
0004698866 00000 п.
0004699594 00000 н.
0004700308 00000 п.
0004700536 00000 п.
0004701251 00000 п.
0004701969 00000 п.
0004702687 00000 п.
0004703408 00000 п.
0004703636 00000 п.
0004704342 00000 п.
0004704570 00000 п.
0004704852 00000 п.
0004706633 00000 п.
0004708469 00000 п.
0004709204 00000 п.
0004709880 00000 п.
0004710268 00000 п.
0004710622 00000 п.
0004711326 00000 п.
0004711674 00000 п.
0004712381 00000 п.
0004712739 00000 п.
0004713450 00000 п.
0004713778 00000 п.
0004714225 00000 п.
0004714929 00000 п.
0004715516 00000 п.
0004716224 00000 п.
0004716927 00000 n
0004717648 00000 н.
0004718353 00000 п.
0004719058 00000 н.
0004719508 00000 п.
0004720212 00000 п.
0004720920 00000 н.
0004721545 00000 п.
0004722252 00000 п.
0004722956 00000 п.
0004723660 00000 п.
0004724370 00000 п.
0004725078 00000 п.
0004725805 00000 п.
0004726256 00000 п.
0004730432 00000 п.
0004730618 00000 п.
0004731081 00000 п.
0004697943 00000 п.
0004698151 00000 п.
0004698866 00000 п.
0004699594 00000 н.
0004700308 00000 п.
0004700536 00000 п.
0004701251 00000 п.
0004701969 00000 п.
0004702687 00000 п.
0004703408 00000 п.
0004703636 00000 п.
0004704342 00000 п.
0004704570 00000 п.
0004704852 00000 п.
0004706633 00000 п.
0004708469 00000 п.
0004709204 00000 п.
0004709880 00000 п.
0004710268 00000 п.
0004710622 00000 п.
0004711326 00000 п.
0004711674 00000 п.
0004712381 00000 п.
0004712739 00000 п.
0004713450 00000 п.
0004713778 00000 п.
0004714225 00000 п.
0004714929 00000 п.
0004715516 00000 п.
0004716224 00000 п.
0004716927 00000 n
0004717648 00000 н.
0004718353 00000 п.
0004719058 00000 н.
0004719508 00000 п.
0004720212 00000 п.
0004720920 00000 н.
0004721545 00000 п.
0004722252 00000 п.
0004722956 00000 п.
0004723660 00000 п.
0004724370 00000 п.
0004725078 00000 п.
0004725805 00000 п.
0004726256 00000 п.
0004730432 00000 п.
0004730618 00000 п.
0004731081 00000 п. 0004731788 00000 п.
0004732494 00000 п.
0004733200 00000 н.
0004733386 00000 п.
0004733912 00000 п.
0004734196 00000 п.
0004734904 00000 н.
0004735623 00000 п.
0004736332 00000 п.
0004736608 00000 п.
0004737061 00000 п.
0004737766 00000 н.
0004737952 00000 п.
0004738393 00000 п.
0004738579 00000 п.
0004739305 00000 п.
0004740009 00000 п.
0004740213 00000 п.
0004740927 00000 п.
0004741180 00000 п.
0004741891 00000 п.
0004742604 00000 п.
0004742808 00000 п.
0004743019 00000 п.
0004743732 00000 п.
0004744441 00000 п.
0004744621 00000 н.
0004744835 00000 н.
0004745021 00000 п.
0004745725 00000 п.
0004746492 00000 п.
0004747203 00000 п.
0004747908 00000 н.
0004748614 00000 н.
0004748818 00000 н.
0004749524 00000 н.
0004749752 00000 п.
0004750460 00000 п.
0004751167 00000 п.
0004751878 00000 п.
0004752073 00000 п.
0004752770 00000 н.
0004752987 00000 п.
0004753701 00000 п.
0004753917 00000 п.
0004754145 00000 п.
0004754850 00000 п.
0004755554 00000 п.
0004731788 00000 п.
0004732494 00000 п.
0004733200 00000 н.
0004733386 00000 п.
0004733912 00000 п.
0004734196 00000 п.
0004734904 00000 н.
0004735623 00000 п.
0004736332 00000 п.
0004736608 00000 п.
0004737061 00000 п.
0004737766 00000 н.
0004737952 00000 п.
0004738393 00000 п.
0004738579 00000 п.
0004739305 00000 п.
0004740009 00000 п.
0004740213 00000 п.
0004740927 00000 п.
0004741180 00000 п.
0004741891 00000 п.
0004742604 00000 п.
0004742808 00000 п.
0004743019 00000 п.
0004743732 00000 п.
0004744441 00000 п.
0004744621 00000 н.
0004744835 00000 н.
0004745021 00000 п.
0004745725 00000 п.
0004746492 00000 п.
0004747203 00000 п.
0004747908 00000 н.
0004748614 00000 н.
0004748818 00000 н.
0004749524 00000 н.
0004749752 00000 п.
0004750460 00000 п.
0004751167 00000 п.
0004751878 00000 п.
0004752073 00000 п.
0004752770 00000 н.
0004752987 00000 п.
0004753701 00000 п.
0004753917 00000 п.
0004754145 00000 п.
0004754850 00000 п.
0004755554 00000 п.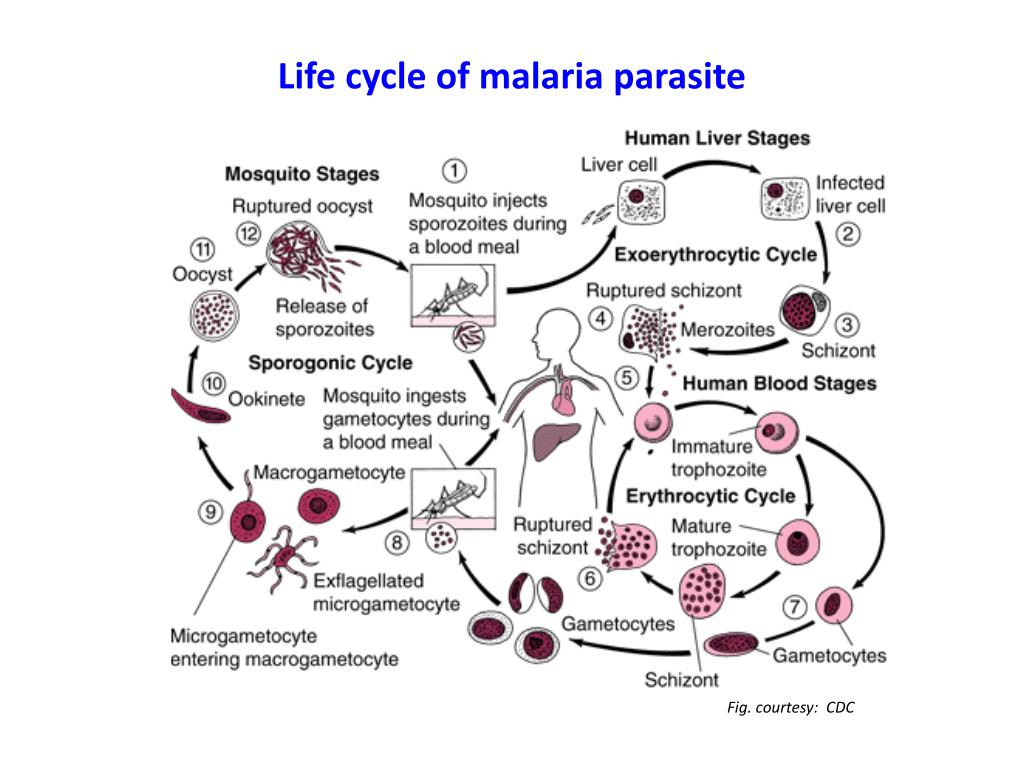 0004755740 00000 п.
0004755920 00000 н.
0004756630 00000 п.
0004756810 00000 п.
0004759928 00000 п.
0004760114 00000 п.
0004760318 00000 п.
0004760534 00000 п.
0004761240 00000 п.
0004761946 00000 н.
0004762159 00000 п.
0004762345 00000 п.
0004762531 00000 н.
0004762735 00000 п.
0004762939 00000 п.
0004763649 00000 п.
0004763869 00000 п.
0004764073 00000 п.
0004764779 00000 н.
0004765485 00000 н.
0004766198 00000 п.
0004766393 00000 п.
0004766598 00000 п.
0004767313 00000 п.
0004768034 00000 п.
0004768220 00000 n
0004768930 00000 п.
0004769135 00000 п.
0004769844 00000 н.
0004770048 00000 н.
0004770234 00000 п.
0004770420 00000 n
0004771134 00000 п.
0004771840 00000 п.
0004772554 00000 п.
0004772770 00000 н.
0004772977 00000 п.
0004773184 00000 п.
0004773364 00000 п.
0004774070 00000 п.
0004774777 00000 п.
0004774969 00000 п.
0004775161 00000 п.
0004775353 00000 п.
0004775987 00000 п.
0004776694 00000 п.
0004777417 00000 п.
0004778138 00000 п.
0004755740 00000 п.
0004755920 00000 н.
0004756630 00000 п.
0004756810 00000 п.
0004759928 00000 п.
0004760114 00000 п.
0004760318 00000 п.
0004760534 00000 п.
0004761240 00000 п.
0004761946 00000 н.
0004762159 00000 п.
0004762345 00000 п.
0004762531 00000 н.
0004762735 00000 п.
0004762939 00000 п.
0004763649 00000 п.
0004763869 00000 п.
0004764073 00000 п.
0004764779 00000 н.
0004765485 00000 н.
0004766198 00000 п.
0004766393 00000 п.
0004766598 00000 п.
0004767313 00000 п.
0004768034 00000 п.
0004768220 00000 n
0004768930 00000 п.
0004769135 00000 п.
0004769844 00000 н.
0004770048 00000 н.
0004770234 00000 п.
0004770420 00000 n
0004771134 00000 п.
0004771840 00000 п.
0004772554 00000 п.
0004772770 00000 н.
0004772977 00000 п.
0004773184 00000 п.
0004773364 00000 п.
0004774070 00000 п.
0004774777 00000 п.
0004774969 00000 п.
0004775161 00000 п.
0004775353 00000 п.
0004775987 00000 п.
0004776694 00000 п.
0004777417 00000 п.
0004778138 00000 п.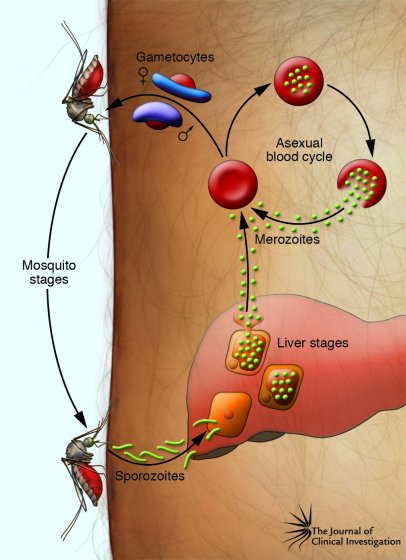 0004778825 00000 п.
0004779302 00000 п.
0004779763 00000 п.
0004779943 00000 н.
0004780687 00000 п.
0004781449 00000 п.
0004781644 00000 п.
0004781858 00000 н.
0004782563 00000 п.
0004782779 00000 н.
0004783483 00000 п.
0004783874 00000 н.
0004787131 00000 п.
0004787838 00000 н.
0004788078 00000 н.
0004788291 00000 п.
0004788507 00000 п.
0004789214 00000 п.
0004789933 00000 н.
00047
0004778825 00000 п.
0004779302 00000 п.
0004779763 00000 п.
0004779943 00000 н.
0004780687 00000 п.
0004781449 00000 п.
0004781644 00000 п.
0004781858 00000 н.
0004782563 00000 п.
0004782779 00000 н.
0004783483 00000 п.
0004783874 00000 н.
0004787131 00000 п.
0004787838 00000 н.
0004788078 00000 н.
0004788291 00000 п.
0004788507 00000 п.
0004789214 00000 п.
0004789933 00000 н.
00047
00000 п. 00047 00000 п. 00047
00000 н. 0004792727 00000 н. 0004793797 00000 п. 0004794501 00000 п. 0004800509 00000 пн 0004807825 00000 п. 0004808530 00000 п. 0004811361 00000 п. 0004812066 00000 н. 0004812771 00000 п. 0004812957 00000 п. 0004813664 00000 п. 0004813867 00000 п. 0004814074 00000 п. 0000034436 00000 п. трейлер ] / Назад 4862651 >> startxref 0 %% EOF 1710 0 объект > поток hV {pTǽ {77ds $ h # rSe Ո PngiЕItFE @ $]! Ҩg5U 芑 RAn` ޙ | = v4? fY8FD_ $ 1A $ b (PI T.pX \ zAux \ .cV)! z \ s> zrb «2 $] $ ** \ BHgF3-15PF 㸭 ޅ * q2D- @ C> fedn4 &» hn2C? @ + ͟aˉ (x4! v ‘& 99uE
Жизненный цикл паразита | Medicines for Malaria Venture
Малярийные паразиты распространяются путем последовательного заражения двух типов хозяев: самок комаров Anopheles и людей. См. Анимированный слайд PowerPoint, показывающий жизненный цикл малярийного паразита.
См. Анимированный слайд PowerPoint, показывающий жизненный цикл малярийного паразита.
- В то же время, когда она принимает кровяную муку для питания своих яиц, самка Anopheles москита вводит спорозоиты в кровоток следующей жертвы малярии.
- Спорозоиты быстро поглощаются клетками печени.
- У всех видов Plasmodium эти паразиты развиваются с образованием шизонтов (многоядерная стадия клетки во время бесполого размножения), из которых развиваются несколько тысяч мерозоитов.
- Только в Plasmodium vivax и Plasmodium ovale часть паразитов на стадии печени (известных как гипнозоиты) остается в состоянии покоя в гепатоцитах. На этой стадии паразит может оставаться в спящем состоянии в течение нескольких месяцев или нескольких лет.Таким образом, эти два вида паразитов могут инициировать цикл бесполого размножения, вызывая клинические симптомы в отсутствие нового укуса комара, в результате чего инфекция, вызванная P. vivax , получила название рецидивирующей малярии.
- Когда клетки печени разрываются, мерозоиты попадают в кровоток, где они быстро проникают в эритроциты. Эти паразиты на стадии крови размножаются бесполым путем, быстро достигая высокой концентрации паразитов и разрушая все инфицированные ими эритроциты, что приводит к клиническим симптомам малярии.
- Триггер пока неизвестен, но небольшой процент мерозоитов дифференцируется на мужские и женские гаметоциты, которые попадают в кровь комара. Именно эти гаметоциты вызывают продолжение цикла передачи вируса комару.
- Мужские и женские гаметоциты сливаются внутри комара, образуя диплоидные зиготы, которые, в свою очередь, становятся оокинетами.
- Эти оокинеты мигрируют в среднюю кишку насекомого, проходят через стенку кишечника и образуют ооцисты.
- Происходит мейотическое деление ооцист и образуются спорозоиты, которые затем мигрируют в слюнные железы самок Anopheles комара, готовые продолжить цикл передачи обратно человеку.
 vivax , получила название рецидивирующей малярии.
vivax , получила название рецидивирующей малярии.
Frontiers | Изучение жизненного цикла малярийных паразитов: биолюминесцентный плазмодий в фундаментальных и прикладных исследованиях
Введение
Половина населения мира подвержена риску малярии (Всемирная организация здравоохранения, 2013 г.), наиболее распространенного и тяжелого паразитарного заболевания, передаваемого комарами (White et al., 2014). Пять видов простейших из рода Plasmodium инфицируют людей, причем Plasmodium falciparum и P. vivax вызывают более 200 миллионов случаев в год, а P. falciparum вызывают практически все 6–700 000 смертей в год (2013 г.), зарегистрированных в основном у детей в странах Африки к югу от Сахары.
Малярийный паразит проявляет сложный жизненный цикл с участием комара Anopheles и позвоночного хозяина (рис. 1). Когда инфицированная самка комара кусает человека, спорозоиты Plasmodium перемещаются в печень и проникают в гепатоциты, где паразиты размножаются как печеночные шизонты, пока несколько тысяч мерозоитов не будут продуцированы и выпущены в кровоток. У P. vivax , но не у P. falciparum , некоторые печеночные паразиты остаются неподвижными (гипнозоиты), возобновляя размножение и заражение через несколько недель или месяцев. При инвазии эритроцитов в кровоток Plasmodium паразиты подвергаются бесполой репликации, образуя зрелых шизонтов, разрыв которых высвобождает мерозоиты, которые вторгаются в новые эритроциты. Некоторые паразиты на стадии крови вместо этого дифференцируются на мужские и женские гаметоциты, которые при попадании в кровь комара активируются с образованием гамет.В результате слияния гамет в средней кишке насекомых образуется зигота, которая превращается в подвижную оокинету, пересекает стенку кишечника и превращается в ооцисту, в которой образуются тысячи спорозоитов. Жизненный цикл замыкается, когда спорозоиты, мигрировавшие из разорванной ооцисты в слюнные железы комара, вводятся новому человеку-хозяину через укус насекомого.
У P. vivax , но не у P. falciparum , некоторые печеночные паразиты остаются неподвижными (гипнозоиты), возобновляя размножение и заражение через несколько недель или месяцев. При инвазии эритроцитов в кровоток Plasmodium паразиты подвергаются бесполой репликации, образуя зрелых шизонтов, разрыв которых высвобождает мерозоиты, которые вторгаются в новые эритроциты. Некоторые паразиты на стадии крови вместо этого дифференцируются на мужские и женские гаметоциты, которые при попадании в кровь комара активируются с образованием гамет.В результате слияния гамет в средней кишке насекомых образуется зигота, которая превращается в подвижную оокинету, пересекает стенку кишечника и превращается в ооцисту, в которой образуются тысячи спорозоитов. Жизненный цикл замыкается, когда спорозоиты, мигрировавшие из разорванной ооцисты в слюнные железы комара, вводятся новому человеку-хозяину через укус насекомого.
РИСУНОК 1. Трансгенные биолюминесцентные малярийные паразиты на разных стадиях своего развития у позвоночных и комаров-хозяев . Центральная диаграмма представляет жизненный цикл Plasmodium , показывая прогрессию через стадии развития паразитов в комарах-переносчиках и в позвоночных-хозяевах. (A) Биолюминесцентная визуализация (BLI) индивидуальных гаметоцитов Plasmodium falciparum , экспрессирующих люциферазу жуков-щелкунов под промотором, специфичным для половой стадии. Изображение в ярком поле показывает иммобилизованные гаметоциты, выделенные зеленым цветом, среди неинфицированных эритроцитов; темное поле показывает сигнал биолюминесценции гаметоцитов, инкубированных с D-люциферином.Полоса увеличения: 15 мкм. (По материалам Cevenini et al., 2014). (B) BLI мыши, инфицированной бесполыми паразитами P. berghei , экспрессирующими слияние люциферазы светлячка-зеленый флуоресцентный белок (GFP). Тепловая карта биолюминесцентного сигнала идентифицирует места скопления паразитов (Воспроизведено с разрешения Claser et al., 2011). (C) Флуоресценция слитого белка люциферазы светлячка-GFP, экспрессируемого в спорозоитах P.
Центральная диаграмма представляет жизненный цикл Plasmodium , показывая прогрессию через стадии развития паразитов в комарах-переносчиках и в позвоночных-хозяевах. (A) Биолюминесцентная визуализация (BLI) индивидуальных гаметоцитов Plasmodium falciparum , экспрессирующих люциферазу жуков-щелкунов под промотором, специфичным для половой стадии. Изображение в ярком поле показывает иммобилизованные гаметоциты, выделенные зеленым цветом, среди неинфицированных эритроцитов; темное поле показывает сигнал биолюминесценции гаметоцитов, инкубированных с D-люциферином.Полоса увеличения: 15 мкм. (По материалам Cevenini et al., 2014). (B) BLI мыши, инфицированной бесполыми паразитами P. berghei , экспрессирующими слияние люциферазы светлячка-зеленый флуоресцентный белок (GFP). Тепловая карта биолюминесцентного сигнала идентифицирует места скопления паразитов (Воспроизведено с разрешения Claser et al., 2011). (C) Флуоресценция слитого белка люциферазы светлячка-GFP, экспрессируемого в спорозоитах P.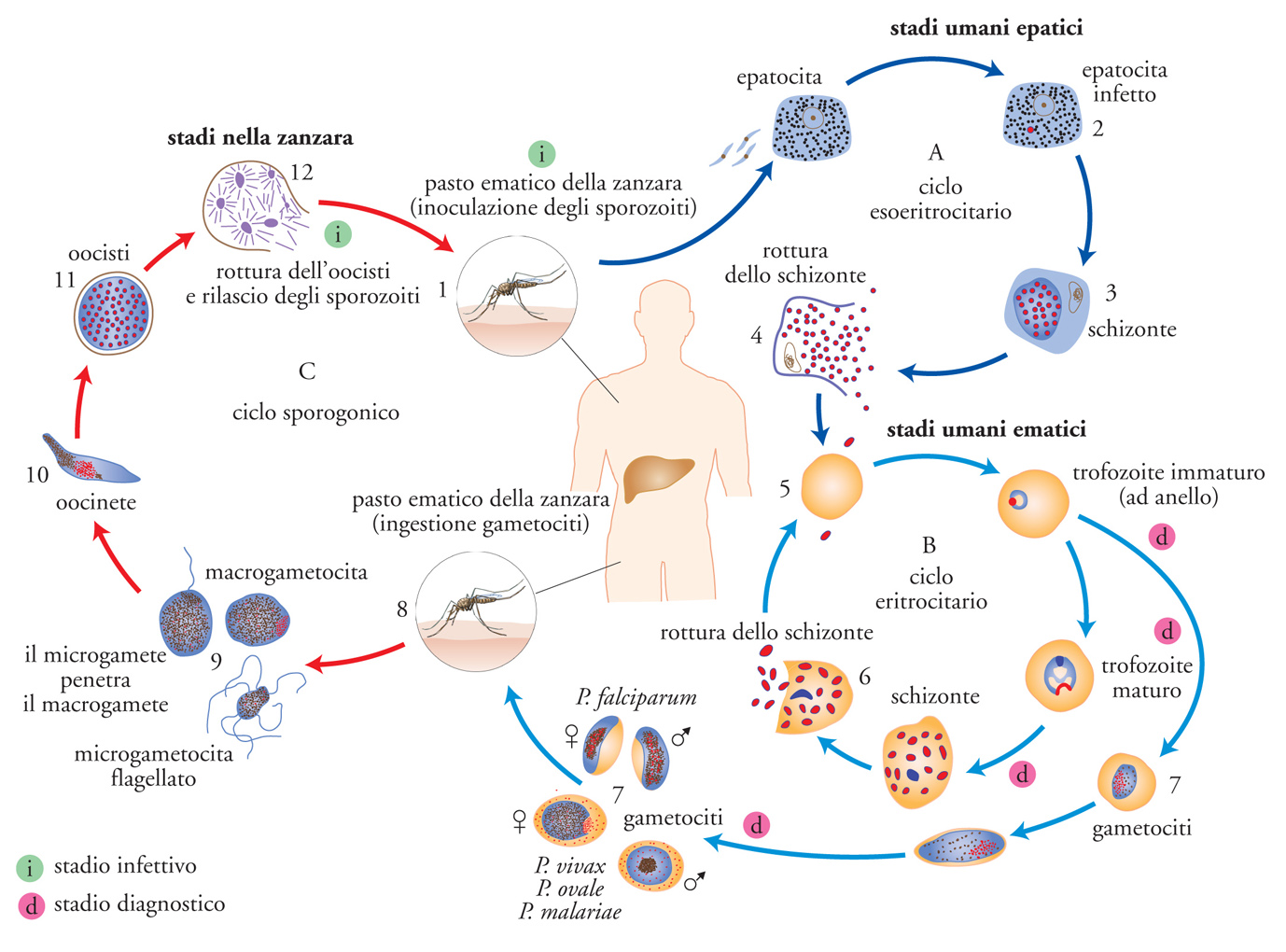 falciparum, , содержащихся в ооцисте, и (D), , полученных в результате вскрытия инфицированных слюнных желез комаров.Полоса увеличения: 5 мкм. (E) Биолюминесцентный сигнал in vivo, полученный трансгенными паразитами на стадии печени P. falciparum , развивающимися в химерной печени гуманизированной мыши. (C – E) воспроизведены с разрешения Vaughan et al. (2012).
falciparum, , содержащихся в ооцисте, и (D), , полученных в результате вскрытия инфицированных слюнных желез комаров.Полоса увеличения: 5 мкм. (E) Биолюминесцентный сигнал in vivo, полученный трансгенными паразитами на стадии печени P. falciparum , развивающимися в химерной печени гуманизированной мыши. (C – E) воспроизведены с разрешения Vaughan et al. (2012).
Патогенез малярии обусловлен стадиями бесполой крови. При клинических проявлениях малярии, вызванной P. falciparum , способность паразитов изолироваться в микроциркуляторном русле некоторых органов, включая мозг, является основной причиной тяжести заболевания и летального исхода (Miller et al., 2002; Milner et al., 2014; Sack et al., 2014). Следовательно, необходимость вылечить пациентов с симптомами традиционно побуждала к поиску лекарств, нацеленных на паразитов бесполой стадии крови, часто недооценивая важность устранения также стадий передачи спорозоитов и гаметоцитов или, в случае P. vivax , гипнозоитов. Недавние тревожные сообщения из Юго-Восточной Азии о снижении чувствительности примерно инфекций P. falciparum к комбинированной терапии на основе производных артемизинина теперь требуют возобновления усилий по решению этой чрезвычайной ситуации в рамках глобальной стратегии борьбы с малярией и, в конечном итоге, искоренить этого смертоносного паразита.
vivax , гипнозоитов. Недавние тревожные сообщения из Юго-Восточной Азии о снижении чувствительности примерно инфекций P. falciparum к комбинированной терапии на основе производных артемизинина теперь требуют возобновления усилий по решению этой чрезвычайной ситуации в рамках глобальной стратегии борьбы с малярией и, в конечном итоге, искоренить этого смертоносного паразита.
Можно культивировать все стадии бесполой и половой крови P. falciparum in vitro , в отличие от P. vivax . Plasmodium видов, инфицирующих грызунов, также интенсивно изучались в качестве мышиных моделей аспектов малярии, причем P. berghei особенно использовались из-за его способности к генетическим манипуляциям. Напротив, технология трансгенеза была сравнительно более сложной для P. falciparum . Этот обзор направлен на то, чтобы подчеркнуть важность трансгенных паразитов Plasmodium , особенно тех, которые были созданы с использованием биолюминесцентных репортеров, как в изучении фундаментальной биологии плазмодия , так и в разработке эффективных противомалярийных препаратов.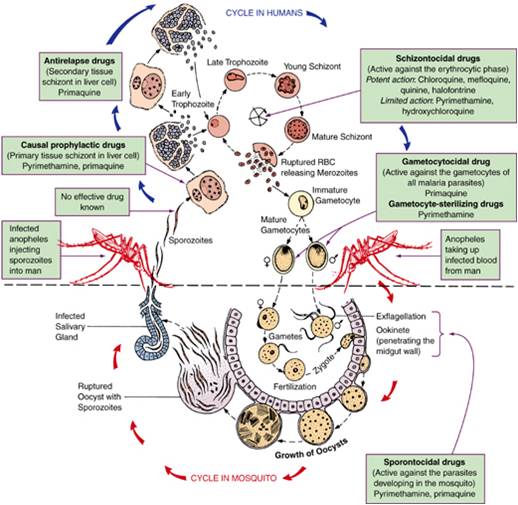
Ферменты люциферазы катализируют светообразующие химические реакции биолюминесцентных организмов, в которых люминогенный субстрат (например, D-люциферин) окисляется в присутствии АТФ с образованием фотонов. Их можно точно измерить люминометром с чувствительностью и фактическим отсутствием фона, что сделало биолюминесцентные репортеры мощным и универсальным инструментом в биологии (Smale, 2010). Люциферазы занимают особое место в истории трансгенеза Plasmodium : первая плазмидная конструкция, успешно трансфицированная малярийными паразитами, содержала ген люциферазы светлячка ( Photynus pyralis ), экспрессия которого обусловлена промотором паразита, специфичного для половой стадии. ген, измерен в оокинетах паразита птиц P.gallinaceum (Goonewardene et al., 1993). Впоследствии репортеры люциферазы были использованы для оптимизации методов трансфекции у паразитов Plasmodium (Epp et al., 2008; Hasenkamp et al., 2012), включая введение люциферазы из морских анютиных глазок Renilla reniformis , где использовали разные субстраты (D-люциферин и целентеразин) позволяли одновременно обнаруживать два репортера, продуцируемых паразитами (Militello and Wirth, 2003; Helm et al.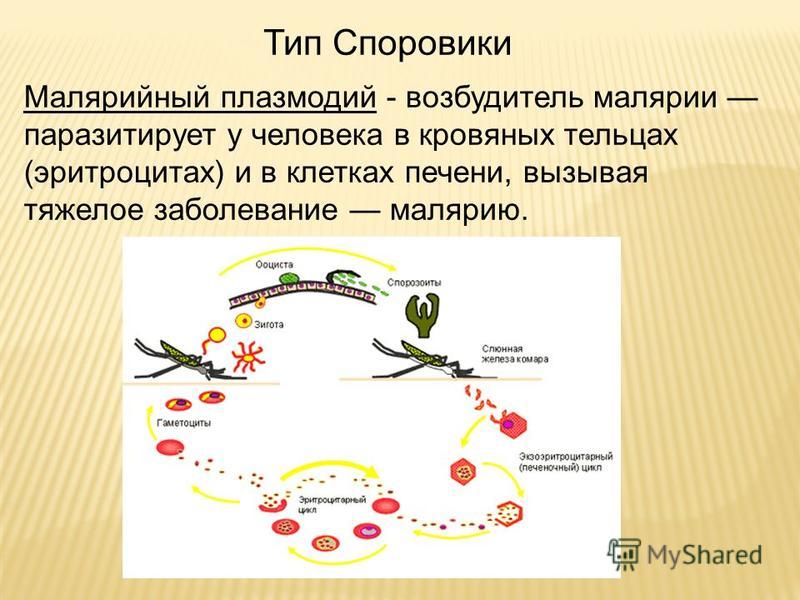 , 2010). С 1990-х годов при стабильной генетической трансформации разных видов Plasmodium (Waters et al., 1997), гены-репортеры люциферазы внесли большой вклад в выяснение ключевых аспектов малярийной инфекции, начиная с клеточной биологии паразитов, трафика белков, функций генов и устойчивости к лекарствам, на нескольких стадиях развития на протяжении жизненного цикла Plasmodium (рис.
, 2010). С 1990-х годов при стабильной генетической трансформации разных видов Plasmodium (Waters et al., 1997), гены-репортеры люциферазы внесли большой вклад в выяснение ключевых аспектов малярийной инфекции, начиная с клеточной биологии паразитов, трафика белков, функций генов и устойчивости к лекарствам, на нескольких стадиях развития на протяжении жизненного цикла Plasmodium (рис.
Жизненный цикл плазмодия
, отмеченный стадиями развития биолюминесцентных паразитов Плазмодий Стадии комаров Развитие половой стадии паразита у комара-переносчика имеет решающее значение для передачи Plasmodium , и поэтому выяснение биологии этого процесса может привести к разработке новых стратегий блокирования передачи малярии.Некоторые исследования биолюминесцентных паразитов подчеркнули важность посттранскрипционной регуляции, действующей на стабильность и трансляцию нескольких мРНК, включая те, которые кодируют основные белки поверхности гамет и оокинет (Mair et al.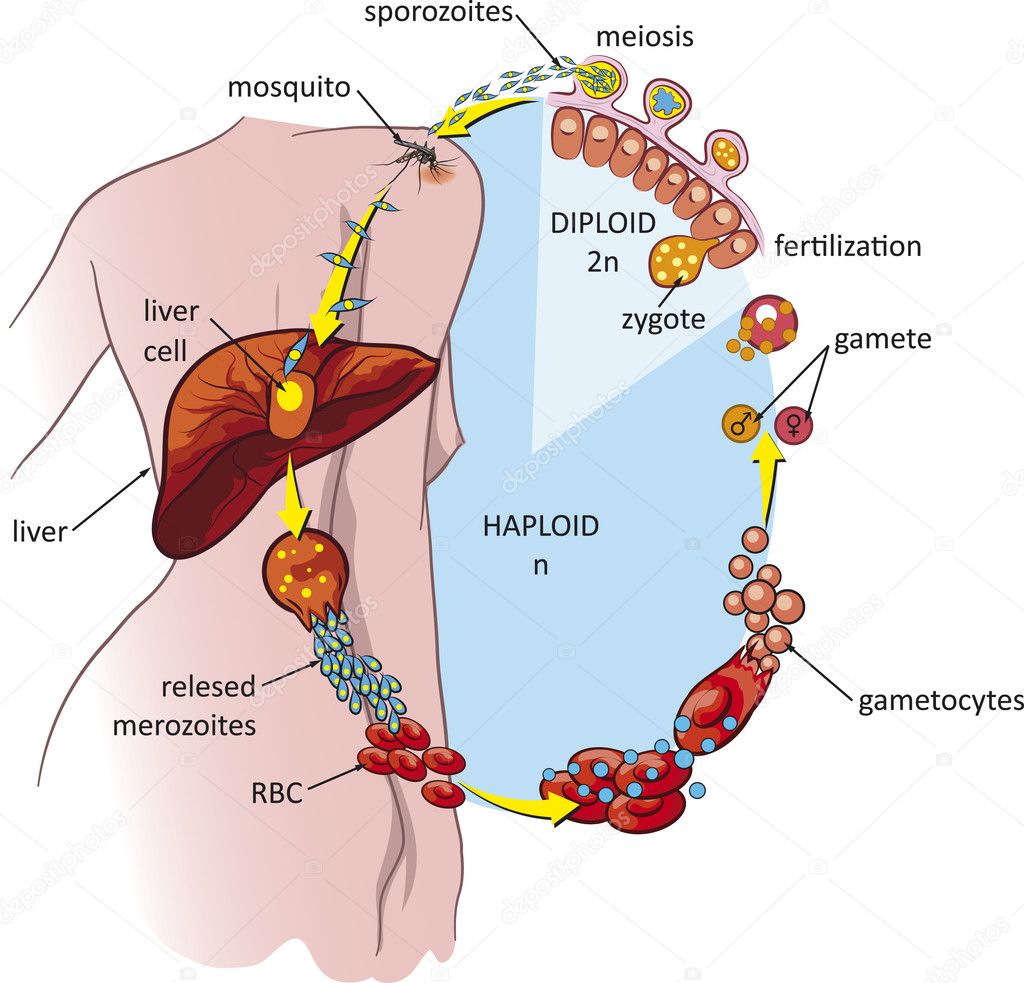 , 2006). Например, анализы с люциферазными репортерами были основополагающими для идентификации регуляторных элементов в транскриптах поверхностных белков P25 и P28 P. gallinaceum и P. falciparum (Golightly et al., 2000; Oguariri et al., 2006).
, 2006). Например, анализы с люциферазными репортерами были основополагающими для идентификации регуляторных элементов в транскриптах поверхностных белков P25 и P28 P. gallinaceum и P. falciparum (Golightly et al., 2000; Oguariri et al., 2006).
Plasmodium , паразиты, экспрессирующие люциферазы, также улучшили разработку инструментов для прикладных исследований. Мощный биоанализ для определения способности паразитов инфицировать комаров основан на кормлении москитами культивированных гаметоцитов Plasmodium и используется для измерения эффекта от лекарств или антител, блокирующих передачу. Однако этот анализ технически сложен и требует много времени, поскольку полученные ооцисты необходимо индивидуально подсчитывать у препарированных насекомых.После улучшений с использованием паразитов P. berghei , экспрессирующих зеленый флуоресцентный белок (GFP) на стадиях комара (Delves and Sinden, 2010), была разработана трансгенная линия паразита человека линия P.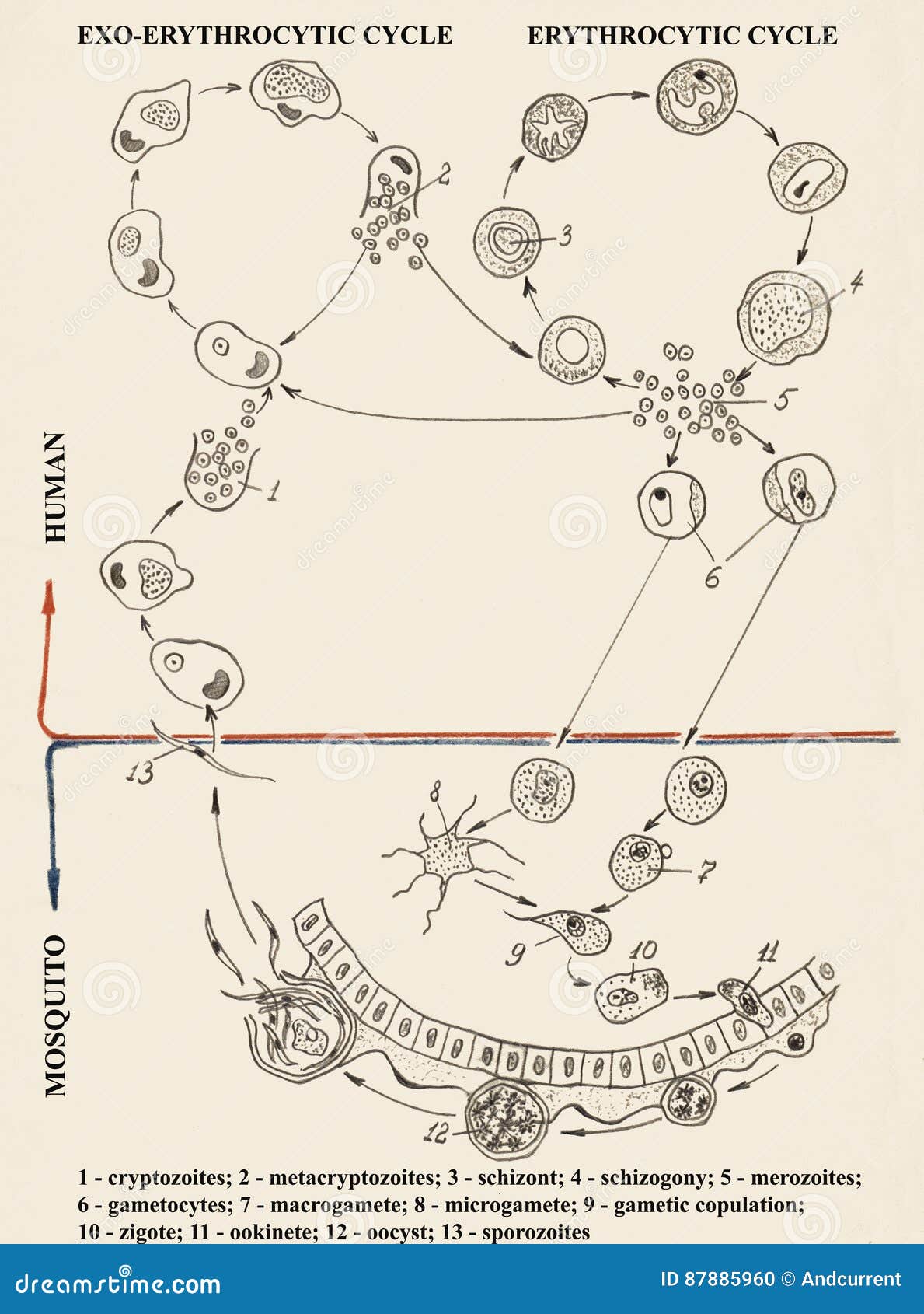 falciparum , экспрессирующая люциферазу светлячка в ооцистах. . В результирующем стандартном анализе мембранного питания (SMFA), основанном на люминесценции, средняя интенсивность люминесценции отдельных и объединенных комаров точно определяла среднюю интенсивность ооцист, устраняя необходимость в вскрытии комаров и создавая основу для значительной масштабируемости SMFA (Stone et al., 2014).
falciparum , экспрессирующая люциферазу светлячка в ооцистах. . В результирующем стандартном анализе мембранного питания (SMFA), основанном на люминесценции, средняя интенсивность люминесценции отдельных и объединенных комаров точно определяла среднюю интенсивность ооцист, устраняя необходимость в вскрытии комаров и создавая основу для значительной масштабируемости SMFA (Stone et al., 2014).
Ближе к концу развития паразита у комара спорозоиты, образующиеся в ооцисте, мигрируют в слюнные железы насекомых. Количество спорозоитов слюнных желез, единственных комаров, заразных для млекопитающих-хозяев, является важным показателем развития комаров Plasmodium . Конструирование линии P. berghei , в которой слияние GFP-люциферазы специфически экспрессируется в спорозоитах, позволило провести простой и быстрый анализ нагрузки спорозоитом от целых комаров (Ramakrishnan et al., 2012).
Передача от комаров: спорозоиты и стадии печени
Plasmodium спорозоиты, введенные инфицированным комаром человеку или грызуну-хозяину, начинают свое внутриклеточное развитие в гепатоцитах печени. Эта клинически бессимптомная стадия является мишенью для профилактических или вакцинных стратегий, особенно против P. vivax длительно действующих гипнозоитов.
Эта клинически бессимптомная стадия является мишенью для профилактических или вакцинных стратегий, особенно против P. vivax длительно действующих гипнозоитов.
Plasmodium стадия развития печени мало изучена по сравнению с стадией развития крови отчасти потому, что анализы in vivo, и in vitro, , соответственно, на мышиных моделях и в культивируемых клетках печени ограничены необходимостью жертвовать высокими показателями. численности мышей или неэффективностью заражения спорозоитами культивируемых клеток печени.Трансгенные спорозоиты, экспрессирующие люциферазу, улучшили стратегии обнаружения, введя биолюминесцентную визуализацию (BLI) и систему визуализации in vivo (IVIS) при анализе стадии развития паразитов печени у живых мышей и в культивируемых гепатоцитах. BLI в реальном времени требует инъекции субстрата люциферина в мышь или в рассеченный орган и усиленной видеокамеры с зарядовой связью для подсчета фотонов для измерения испускания фотонов (Franke-Fayard et al.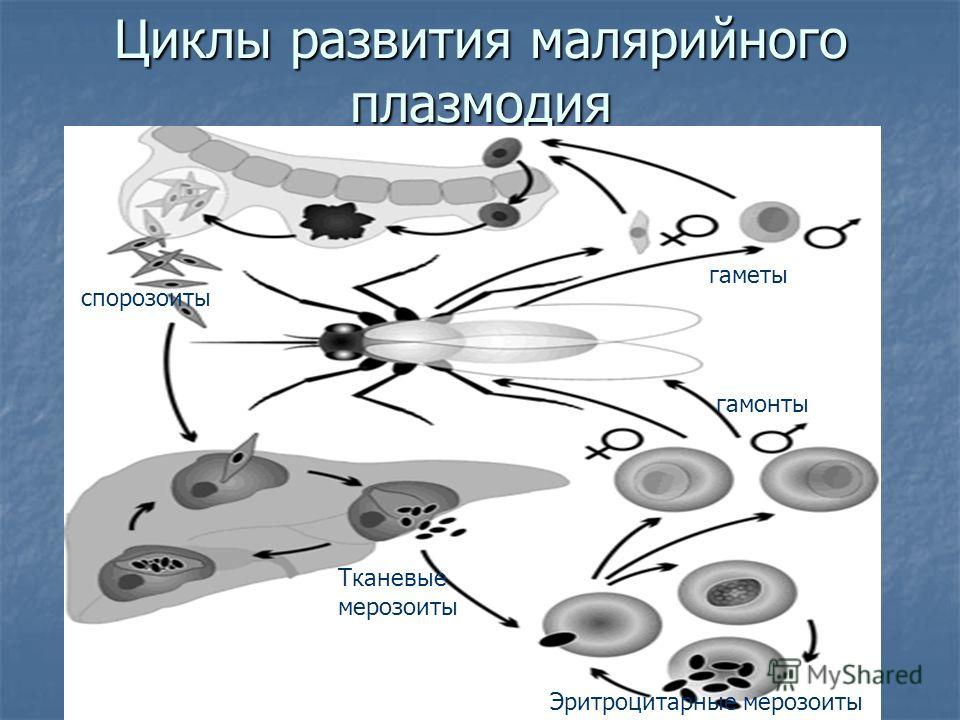 , 2006; Braks et al., 2013). BLI и IVIS с использованием люцифераз светлячков или морских анютиных глазок использовались для мониторинга в реальном времени прогрессирования паразитарной инфекции грызунов у всего животного или в конкретных органах (Ploemen et al., 2009; Annoura et al., 2013; Manzoni et al., 2014), а также для тестирования активности лекарств, направленных на инфекцию на стадии печени, с использованием трансгенных спорозоитов P. yoelii и P. berghei в клетках печени человека HepG2 или Huh-7 и у целых мышей (Mwakingwe et al. , 2009; Ramalhete et al., 2011, 2014; Derbyshire et al., 2012; Lacrue et al., 2013; Li et al., 2014; Marcsisin et al., 2014; Zuzarte-Luis et al., 2014). Для улучшения этих подходов идентификация промоторов паразитов, специфически активируемых при развитии печени, была достигнута у P.berghei , также в этом случае полагаясь на использование трансгенных слияний люциферазы-промотора (Helm et al., 2010).
, 2006; Braks et al., 2013). BLI и IVIS с использованием люцифераз светлячков или морских анютиных глазок использовались для мониторинга в реальном времени прогрессирования паразитарной инфекции грызунов у всего животного или в конкретных органах (Ploemen et al., 2009; Annoura et al., 2013; Manzoni et al., 2014), а также для тестирования активности лекарств, направленных на инфекцию на стадии печени, с использованием трансгенных спорозоитов P. yoelii и P. berghei в клетках печени человека HepG2 или Huh-7 и у целых мышей (Mwakingwe et al. , 2009; Ramalhete et al., 2011, 2014; Derbyshire et al., 2012; Lacrue et al., 2013; Li et al., 2014; Marcsisin et al., 2014; Zuzarte-Luis et al., 2014). Для улучшения этих подходов идентификация промоторов паразитов, специфически активируемых при развитии печени, была достигнута у P.berghei , также в этом случае полагаясь на использование трансгенных слияний люциферазы-промотора (Helm et al., 2010).
Возможность надежного количественного определения паразитарной инфекции в гепатоцитах имеет важное значение при разработке вакцин против малярии.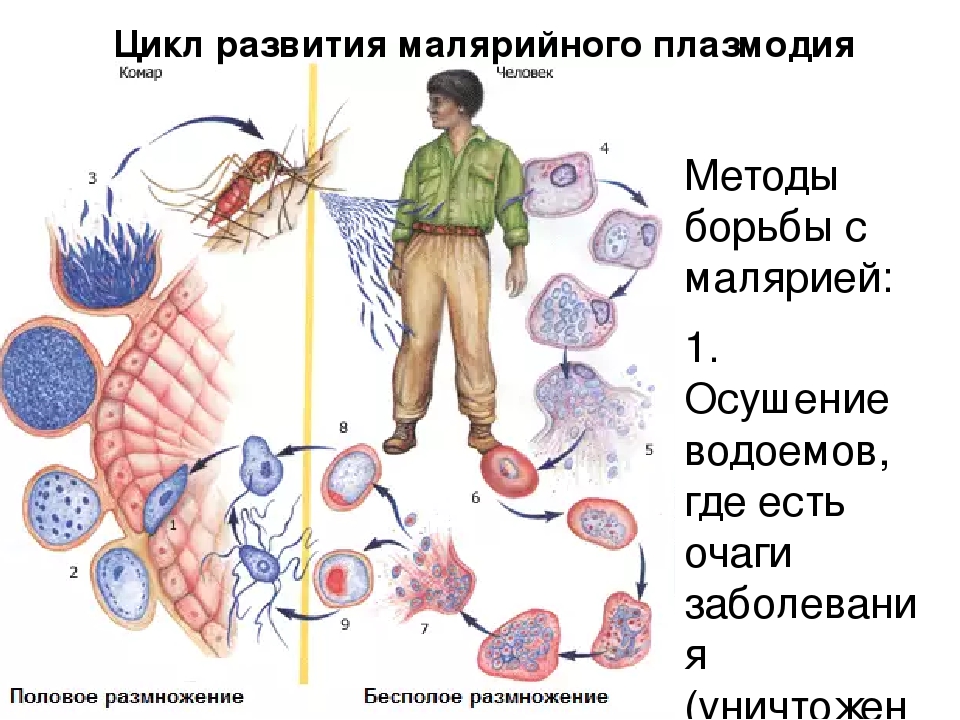 Чтобы преодолеть ограничения количественной оценки на основе qRT-PCR, паразитов P. berghei , экспрессирующих слияние GFP-люциферазы, были введены для оценки противомалярийного иммунитета как in vivo, так и у мышей, у которых он был индуцирован спорозоитами, неспособными размножаться после облучения или хлорохина. профилактика и in vitro в клетках гепатомы печени человека Huh-7 (Ploemen et al., 2011; Миллер и др., 2013). Люцифераза, экспрессирующая спорозоитов P. berghei и P. falciparum также использовалась для оценки адекватности ослабления спорозоитов, полученного на этот раз путем генетической мутации, соответственно, в модели in vivo мышиной малярии и в первичных гепатоцитах человека (Annoura et al. ., 2012; van Schaijk et al., 2014). Эти исследования подчеркнули роль клеточного роста иммунитета против спорозоитов, дефицитных по размножению. Роль опосредованного антителами иммунитета вместо этого была показана BLI экспрессирующих люциферазу спорозоитов паразита человека P.
Чтобы преодолеть ограничения количественной оценки на основе qRT-PCR, паразитов P. berghei , экспрессирующих слияние GFP-люциферазы, были введены для оценки противомалярийного иммунитета как in vivo, так и у мышей, у которых он был индуцирован спорозоитами, неспособными размножаться после облучения или хлорохина. профилактика и in vitro в клетках гепатомы печени человека Huh-7 (Ploemen et al., 2011; Миллер и др., 2013). Люцифераза, экспрессирующая спорозоитов P. berghei и P. falciparum также использовалась для оценки адекватности ослабления спорозоитов, полученного на этот раз путем генетической мутации, соответственно, в модели in vivo мышиной малярии и в первичных гепатоцитах человека (Annoura et al. ., 2012; van Schaijk et al., 2014). Эти исследования подчеркнули роль клеточного роста иммунитета против спорозоитов, дефицитных по размножению. Роль опосредованного антителами иммунитета вместо этого была показана BLI экспрессирующих люциферазу спорозоитов паразита человека P. falciparum у мышей с гуманизированной печенью, показывая, что инфекция в этом органе снижается за счет пассивного переноса моноклональных антител, нацеленных на поверхностный белок спорозоитов CSP (Sack et al., 2014). Наконец, трансгенные паразиты люциферазы P. berghei и P. yoelii сыграли важную роль в оценке способов введения спорозоитов, критического узкого места при иммунизации и протоколов заражения (Ploemen et al., 2013).
falciparum у мышей с гуманизированной печенью, показывая, что инфекция в этом органе снижается за счет пассивного переноса моноклональных антител, нацеленных на поверхностный белок спорозоитов CSP (Sack et al., 2014). Наконец, трансгенные паразиты люциферазы P. berghei и P. yoelii сыграли важную роль в оценке способов введения спорозоитов, критического узкого места при иммунизации и протоколов заражения (Ploemen et al., 2013).
От печени к крови: стадии бесполого эритроцита
Созревание печеночного шизонта высвобождает тысячи мерозоитов, которые проникают в эритроциты кровотока и запускают бесполую симптоматическую инфекцию в крови.В P. falciparum шизонты стадии крови исчезают из кровотока, поскольку они прикрепляются к лигандам хозяина на эндотелиальных клетках микрососудов в нескольких органах, особенно в головном мозге и плаценте, через белки паразита, экспрессируемые на поверхности инфицированных эритроцитов, что приводит к тяжелый патогенез, такой как церебральная малярия или побочные эффекты во время беременности. Поскольку наблюдается накопление паразитов в нескольких органах, включая мозг, а также на модели малярии мышей, BLI в реальном времени у целых мышей или в разрезанных органах проводился с P.berghei , экспрессирующие люциферазу под конститутивным или шизонт-специфическим промотором для идентификации задействованных компонентов иммунной системы (Franke-Fayard et al., 2005; Amante et al., 2007; Spaccapelo et al., 2010; Claser et al., 2010; Claser et al., 2005; Amante et al., 2007; Spaccapelo et al., 2010; Claser et al. al., 2011; Pasini et al., 2013; Imai et al., 2014).
Необходимость выяснения механизмов патогенеза малярии направлена на изучение фундаментальной биологии бесполого развития паразита, одним из важных аспектов которого является то, как паразит регулирует экспрессию своих генов.Однако чрезвычайно высокое содержание A + T в геномах Plasmodium препятствовало идентификации промоторов, регуляторных элементов и факторов транскрипции паразитов на основе гомологии, тогда как репортеры люциферазы оказались чрезвычайно важными для функциональной идентификации промоторов генов и регуляторных областей (Horrocks и Kilbey, 1996; Porter, 2002; Militello et al., 2004; Hasenkamp et al., 2013a). Эта работа идентифицировала последовательности, функционирующие как двунаправленные промоторы, такие как межгенная область P.berghei гена фактора элонгации-1α ( ef-1α ) (de Koning-Ward et al., 1999; Fernandez-Becerra et al., 2003) или интрона генов P. falciparum var (Epp et al. ., 2008), или оценивали, способны ли специфические промоторы одного вида Plasmodium (Fernandez-Becerra et al., 2003; Ozwara et al., 2003) или неспособны (Azevedo and del Portillo, 2007) рекрутировать транскрипционный механизмы малярии другого вида. Важно отметить, что паразиты, экспрессирующие люциферазу, были использованы для идентификации регуляторных областей, управляющих экспрессией P.falciparum , полиморфные гены var , кодирующие лиганды секвестрации паразита, переключатель экспрессии которых отвечает за антигенную изменчивость паразита и уклонение от иммунитета (Deitsch et al., 1999; Calderwood et al., 2003; Frank et al., 2006; Muhle et al., 2006; Muhle et al. др., 2009). Таким образом, репортеры люциферазы не только способствовали идентификации функциональных элементов, участвующих в регуляции генов паразитов (Bischoff et al., 2000; Militello et al., 2004; López-Estraño et al., 2007; Gopalakrishnan and López-Estraño, 2010; Zhang и другие., 2011; Patakottu et al., 2012), но также были необходимы для выбора специфических промоторов при разработке индуцируемых экспрессионных систем Plasmodium (de Azevedo et al., 2012; Kolevzon et al., 2014) и для тестирования новых регуляторных областей в хромосомах. интегрированные кассеты люциферазы (Ekland et al., 2011; Weiwer et al., 2011; Che et al., 2012; Khan et al., 2012; Hasenkamp et al., 2013b).
Основным направлением борьбы с малярией, в частности с малярией P. falciparum , был скрининг новых противомалярийных препаратов — мероприятие, которое в связи с появлением резистентности к артемизинину в Юго-Восточной Азии делает крайне актуальным.В последние десятилетия in vitro, методов измерения включения [ 3 H] -меченного гипоксантина и этаноламина или активности паразитарной лактатдегидрогеназы были стандартом для анализов на основе клеток P. falciparum и использовались в крупных лекарственных препаратах. просмотры (Fidock, 2010). Потребность в высокопроизводительных нерадиоактивных анализах побудила также использовать в Plasmodium высокую чувствительность и фактическое отсутствие фона репортеров люциферазы, которые до недавнего времени использовались в этой области только для изучения экспрессии P.falciparum , ген множественной лекарственной устойчивости pfmdr1 у паразитов, леченных лекарствами (Myrick et al., 2003; Waller et al., 2003). С этой целью линия P. falciparum , экспрессирующая люциферазу светлячка под промотором гена белка теплового шока 86 ( pfhsp86 ) на бесполых стадиях, позволила создать клеточный скрининговый анализ лекарственного средства люциферазы в 96w планшетах (Cui et al ., 2008), впоследствии адаптированные к планшету 384w с использованием 10 5 –10 6 паразитов на лунку (Lucumi et al., 2010). Также паразитов P. berghei , экспрессирующих слияние люциферазы светлячка-GFP, были использованы для in vitro и in vivo биолюминесцентного анализа лекарственных средств, что позволило использовать модели на животных для тестирования новых лекарств in vivo (Franke-Fayard et al. , 2008; Lin et al., 2013).
Подготовка к исходу из крови: гаметоциты
Гаметоциты Plasmodium представляют собой половые стадии паразита, ответственные за передачу от позвоночного хозяина к комару.Мужские и женские гаметоциты образуются в кровотоке, и у P. falciparum они созревают за 10 дней, проходя пять стадий развития. При попадании в кишечник комара зрелые гаметоциты быстро дифференцируются в гаметы, а оплодотворение обеспечивает заражение паразитами насекомых-переносчиков. Ключевым приоритетом в нынешней цели по глобальной ликвидации малярии является выявление новых лекарств, нацеленных на кровоток как на бесполой, так и на половой стадиях паразита. Однако нерепликативная природа гаметоцитов необходима для разработки специфических клеточных скрининговых анализов, отличных от тех, которые используются для бесполых стадий.Одна из проблем — это, например, ложноотрицательные сигналы из-за сохранения флуоресцентного репортера или активности паразитарных ферментов в нездоровых или умирающих гаметоцитах. P. falciparum Линии , экспрессирующие ген люциферазы GFP-светлячка под промоторами, специфичными для гаметоцитов, были созданы (Adjalley et al., 2011) и использованы в высокопроизводительных скрининговых анализах соединений с активностью на анти-асексуальной стадии (Lucantoni et al., 2013). ). В последнее время методы анализа гаметоцитов на основе люциферазы были усовершенствованы путем замены обычно используемых коммерческих субстратов люциферазы на не содержащий АТФ и не лизирующий состав D-люциферина, в результате чего были получены результаты анализа, позволяющие более надежно контролировать жизнеспособность и чувствительность обработанных гаметоцитов к соединениям (Cevenini). и другие., 2014). В данной работе отсутствие лизиса клеток паразита и введение в P. falciparum использования сильнодействующей люциферазы из Pyrophorus plagiophthalamus под промотором гаметоцитов позволило впервые провести BLI на уровне одиночных клеток паразита. индивидуально различая живые и мертвые гаметоцитов P. falciparum (Cevenini et al., 2014).
Мультиплексирование, субклеточная локализация, визуализация: будущее использования биолюминесцентных малярийных паразитов
Практически все стадии сложного жизненного цикла малярийных паразитов были освещены путем использования в нескольких исследованиях репортеров люциферазы.Эти искусственно созданные паразиты дали ключевые ответы на фундаментальные биологические вопросы и теперь представляют собой важные инструменты для скрининга лекарств. Новые мощные репортеры уже расширили репертуар люциферазы, используемой в P. falciparum , за пределы ферментов P. pyralis и Renilla (Azevedo et al., 2014; Cevenini et al., 2014), повысив чувствительность и позволив еще больше снизить количество паразитов. числа в высокопроизводительных скрининговых анализах, нетривиальное улучшение при использовании определенных этапов (например,g., гаметоциты), культивирование которых технически сложно. Тем не менее использование всего потенциала биолюминесцентных репортеров в исследованиях малярии — это только первые шаги.
Возможность настройки свойств излучения люциферазы, таких как длина волны излучения, кинетика или термо- и рН-стабильность, посредством случайного или сайт-направленного мутагенеза или использования природных вариантов ферментов, привела к введению многоцветной биолюминесценции при скрининге противомалярийных препаратов. Люцифераза, излучающая зеленый и красный свет из P.plagiophthalamus были экспрессированы в незрелых и зрелых гаметоцитах P. falciparum , что впервые дает возможность одновременно измерять дифференциальные, стадийно-специфические эффекты лекарств в двухцветном анализе люциферазы и открывает возможность применять многоцветную биолюминесценцию к любым паразитарный этап в фундаментальных и прикладных исследованиях. Система двойной экспрессии с отдельными люциферазами могла бы, например, быть полезной при высокопроизводительном скрининге на основе клеток, чтобы легко идентифицировать и отбрасывать соединения, активные против репортера, а не клетки-мишени (Thorne et al., 2012), так как они, скорее всего, будут влиять только на один тип люциферазы.
Еще одно многообещающее применение люциферазных репортеров заключается в их слиянии со специфическими сигналами, используемыми паразитом для передачи белков в различные внеклеточные компартменты инфицированного эритроцита. Поскольку экспорт белка однозначно регулируется у паразита и необходим для его выживания, использование таких слияний может быть неоценимым для скрининга соединений, нацеленных на этот процесс. Предварительные исследования были проведены с P.pyralis (Burghaus and Lingelbach, 2001) и более поздняя работа установила линий P. falciparum , которые экспрессируют более яркую люциферазу глубоководных креветок, снабженную последовательностями, управляющими репортером в цитоплазме паразита или в компартментах эритроцитов (Azevedo et al., 2014).
В другой области применения достижение BLI отдельных клеток паразита и доступность люцифераз, чье излучение света с красным смещением более эффективно обнаруживается в крови и тканях, прокладывают путь к значительному прогрессу в анализе взаимодействия паразита и хозяина.Совместное культивирование различных стадий P. falciparum и типов клеток человека in vitro может дать новое понимание физиологии секвестрации паразитов на бесполой и половой стадиях. Повышенная чувствительность, достигнутая при визуализации in vivo мышей с люциферазой с красным смещением, экспрессируемой одноклеточным простейшим паразитом Trypanosoma brucei (Van Reet et al., 2014), является многообещающей с точки зрения использования также на мышах, инфицированных Plasmodium . Важно отметить растущую доступность гуманизированных моделей мышей для P.falciparum и P. vivax , поддерживающие развитие бесполой и половой стадий крови и стадии печени (Kaushansky et al., 2014), а также использование трансгенных линий P. falciparum с конститутивно экспрессируемой люциферазой (Vaughan et al. al., 2012) или промоутеры для конкретных стадий, как ожидается, ответят на многие нерешенные вопросы.
Богатство биологической информации, полученной с помощью искусственно созданных биолюминесцентных малярийных паразитов, не говоря уже о тех, которые не рассматриваются здесь, которые экспрессируют множество флуоресцентных репортеров, было и, скорее всего, будет огромным.Ограниченное использование этих цельноклеточных биосенсоров в лабораторных условиях не вызывает озабоченности по поводу выбросов в окружающую среду. Несмотря на асептические условия использования, эти генетически модифицированные паразиты, тем не менее, будут иметь самое значительное влияние в реальном мире, по сравнению с невыносимым бременем разрушительной болезни во всем мире.
Вклад автора
GS подготовил рукопись, а PA отредактировал.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы выражают признательность докторам Луке Севенини, Элизе Мичелини и Альдо Рода, Университет Болоньи, Болонья, Италия, и доктору Брюсу Б. Бранчини, Колледж Коннектикута, Нью-Лондон, Коннектикут, США, за несколько содержательных обсуждений во время нашего сотрудничества и поблагодарить г-на Козимо М. Куриано за рисунок. Исследования в лаборатории авторов поддерживаются грантами Фонда Билла и Мелинды Гейтс и Европейской сети передового опыта EVIMalaR.
Список литературы
Аджаллея, С.Х., Джонстон, Г. Л., Ли, Т., Истман, Р. Т., Экланд, Э. Х., Эппен, А. Г. и др. (2011). Количественная оценка полового развития Plasmodium falciparum выявила сильную блокирующую способность метиленового синего. Proc. Natl. Акад. Sci. США 108, E1214 – E1223. DOI: 10.1073 / pnas.1112037108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Аманте, Ф. Х., Стэнли, А. К., Рэндалл, Л. М., Чжоу, Ю., Хак, А., МакСвини, К. и др.(2007). Роль естественных регуляторных Т-клеток в патогенезе экспериментальной церебральной малярии. Am. J. Pathol. 171, 548–559. DOI: 10.2353 / ajpath.2007.061033
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Annoura, T., Chevalley, S., Janse, C.J., Franke-Fayard, B., and Khan, S.M. (2013). Количественный анализ стадий печени Plasmodium berghei методом биолюминесцентной визуализации. Methods Mol. Биол. 923, 429–443. DOI: 10.1007 / 978-1-62703-026-7_30
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Анноура, Т., Ploemen, I.H., van Schaijk, B.C., Sajid, M., Vos, M.W., van Gemert, G.J., et al. (2012). Оценка адекватности аттенуации кандидатных вакцин против генетически модифицированных паразитов от малярии. Вакцина. 30, 2662–2670. DOI: 10.1016 / j.vaccine.2012.02.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Азеведо, М. Ф., и дель Портильо, Х. А. (2007). Промоторные области Plasmodium vivax плохо или не распознаются Plasmodium falciparum . Малар. J. 6, 20. DOI: 10.1186 / 1475-2875-6-20
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Азеведо, М. Ф., Ни, К. К., Элсворт, Б., Шарно, С. К., Сандерс, П. Р., Крабб, Б. С. и др. (2014). Plasmodium falciparum , трансфицированный сверхъяркой люциферазой NanoLuc, обеспечивает высокую чувствительность обнаружения для скрининга ингибиторов роста и клеточного транспорта. PLoS ONE 9: e112571. DOI: 10.1371 / journal.pone.0112571
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бишофф, Э., Гильотт, М., Мерсеро-Пуйялон, О., и Боннефой, С. (2000). Член мультигенного семейства Plasmodium falciparum Pf60 кодирует ядерный белок, экспрессируемый путем считывания внутреннего стоп-кодона. Мол. Microbiol. 35, 1005–1016. DOI: 10.1046 / j.1365-2958.2000.01788.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бракс, Дж., Эйме, Э., Спаккапело, Р., Клоп, О., Янсе, К. Дж., И Франке-Файяр, Б. (2013). Биолюминесцентное изображение P.berghei Секвестрация шизонтов у грызунов. Methods Mol. Биол . 923, 353–368. DOI: 10.1007 / 978-1-62703-026-7_25
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Burghaus, P. A., and Lingelbach, K. (2001). Люцифераза при слиянии с N-концевым сигнальным пептидом секретируется из трансфицированного Plasmodium falciparum и транспортируется в цитозоль инфицированных эритроцитов. J. Biol. Chem. 276, 26838–26845. DOI: 10.1074 / JBC.M100111200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кальдервуд, М. С., Ганнун-Заки, Л., Веллемс, Т. Э., и Дейч, К. В. (2003). Plasmodium falciparum Гены var регулируются двумя областями с отдельными промоторами, один перед кодирующей областью, а второй — внутри интрона. J. Biol. Chem. 278, 34125–34132. DOI: 10.1074 / jbc.M213065200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чевенини, Л., Camarda, G., Michelini, E., Siciliano, G., Calabretta, M. M., Bona, R., et al. (2014). Многоцветная биолюминесценция способствует исследованиям малярии: количественный двухцветный анализ и визуализация отдельных клеток у паразитов Plasmodium falciparum . Анал. Chem. 86, 8814–8821. DOI: 10.1021 / ac502098w
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Che, P., Cui, L., Kutsch, O., Cui, L., and Li, Q. (2012). Валидация высокопроизводительного скринингового анализа на основе люциферазы светлячков на предмет обнаружения противомалярийных препаратов. Assay Drug Dev. Technol. 10, 61–68. DOI: 10.1089 / adt.2011.0378
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Claser, C., Malleret, B., Gun, S.Y., Wong, A.Y., Chang, Z. W., Teo, P., et al. (2011). CD8 + Т-клетки и IFN-γ опосредуют зависящее от времени накопление инфицированных эритроцитов в глубоких органах во время экспериментальной церебральной малярии. PLoS ONE 6: e18720. DOI: 10.1371 / journal.pone.0018720
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Куй, Л., Мяо, Дж., Ван, Дж., Ли, К., и Цуй, Л. (2008). Plasmodium falciparum : разработка трансгенной линии для скрининга противомалярийных средств с использованием люциферазы светлячков в качестве репортера. Exp. Паразитол. 120, 80–87. DOI: 10.1016 / j.exppara.2008.05.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
de Azevedo, M. F., Gilson, P. R., Gabriel, H. B., Simões, R.F., Angrisano, F., Baum, J., et al. (2012). Систематический анализ стратегий мечения домена индуцируемой деградации FKBP для малярийного паразита человека Plasmodium falciparum . PLoS ONE 7: e40981. DOI: 10.1371 / journal.pone.0040981
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Deitsch, K. W., del Pinal, A., and Wellems, T. E. (1999). Внутрикластерная рекомбинация и переключатели транскрипции var в антигенной вариации Plasmodium falciparum . Мол. Biochem. Паразитол. 101, 107–116. DOI: 10.1016 / S0166-6851 (99) 00062-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
де Конинг-Уорд, Т.Ф., Сперанса, М. А., Уотерс, А. П., и Джансе, К. Дж. (1999). Анализ стадийной специфичности промоторов Plasmodium berghei с использованием люциферазы в качестве репортера. Мол. Biochem. Паразитол. 100, 141–146. DOI: 10.1016 / S0166-6851 (99) 00042-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Делвес, М. Дж., И Синден, Р. Э. (2010). Полуавтоматический метод подсчета флуоресцентных ооцист малярии увеличивает производительность исследований по блокированию передачи. Малар. J. 9, 35. DOI: 10.1186 / 1475-2875-9-35
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дербишир, Э. Р., Пруденсио, М., Мота, М. М., и Кларди, Дж. (2012). Паразиты малярии на стадии печени уязвимы для различных химических структур. Proc. Natl. Акад. Sci. США 109, 8511–8516. DOI: 10.1073 / pnas.1118370109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Экланд Э. Х., Шнайдер Дж. И Фидок Д.А. (2011). Выявление противомалярийных средств, нацеленных на апикопласты, с использованием подходов, совместимых с высокой пропускной способностью. FASEB J. 25, 3583–3593. DOI: 10.1096 / fj.11-187401
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эпп К., Раскольников Д. и Дейч К. В. (2008). Регулируемая система экспрессии трансгена для культивируемых паразитов Plasmodium falciparum . Малар. J. 7, 86. DOI: 10.1186 / 1475-2875-7-86
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фернандес-Бесерра, К., де Азеведо, М. Ф., Ямамото, М. М., и дель Портильо, Х. А. (2003). Plasmodium falciparum : новый вектор с двунаправленной промоторной активностью для стабильной экспрессии трансгенов. Exp. Паразитол. 103, 88–91. DOI: 10.1016 / S0014-4894 (03) 00065-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Франк, М., Дзиковски, Р., Константини, Д., Амулич, Б., Бердуго, Э. и Дейч, К. (2006). Строгое спаривание промоторов и интронов var необходимо для подавления гена var у малярийного паразита Plasmodium falciparum . J. Biol. Chem. 281, 9942–9952 DOI: 10.1074 / jbc.M513067200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Franke-Fayard, B., Djokovic, D., Dooren, M. W., Ramesar, J., Waters, A. P., Falade, M. O., et al. (2008). Простой и чувствительный скрининг противомалярийных препаратов in vitro и in vivo с использованием трансгенной люциферазы, экспрессирующей Plasmodium berghei паразитов. Внутр. J. Parasitol. 38, 1651–1662. DOI: 10.1016 / j.ijpara.2008.05.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Franke-Fayard, B., Janse, C.J., Cunha-Rodrigues, M., Ramesar, J., Büscher, P., Que, I., et al. (2005). Секвестрация малярийных паразитов у мышей: CD36 является основным рецептором, но церебральная патология не связана с секвестрацией. Proc. Natl. Акад. Sci. США 102, 11468–11473. DOI: 10.1073 / pnas.0503386102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Franke-Fayard, B., Waters, A.P., and Janse, C.J. (2006). Визуализация in vivo трансгенных биолюминесцентных стадий в крови паразитов малярии грызунов у мышей. Nat. Protoc. 1, 476–485. DOI: 10.1038 / nprot.2006.69
CrossRef Полный текст | Google Scholar
Голайтли, Л. М., Мбахам, В., Дейли, Дж. И Вирт, Д. Ф. (2000). Элементы 3 ’UTR усиливают экспрессию Pgs28, белка оокинета Plasmodium gallinaceum . Мол. Biochem. Паразитол. 105, 61–70. DOI: 10.1016 / S0166-6851 (99) 00165-6
CrossRef Полный текст | Google Scholar
Goonewardene, R., Daily, J., Kaslow, D., Sullivan, T.J., Duffy, P., Carter, R., et al. (1993). Трансфекция малярийного паразита и экспрессия люциферазы светлячков. Proc. Natl. Акад. Sci. США 90, 5234–5236. DOI: 10.1073 / pnas.90.11.5234
CrossRef Полный текст | Google Scholar
Гопалакришнан, А. М., и Лопес-Эстраньо, К. (2010). Роль цис-регуляторных элементов в кольцевом промоторе hrp3 у паразита человека Plasmodium falciparum . Parasitol Res. 106, 833–845. DOI: 10.1007 / s00436-010-1738-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хазенкамп, С., Рассел, К. Т., и Хоррокс, П. (2012). Сравнение абсолютной и относительной эффективности протоколов трансфекции на основе электропорации для Plasmodium falciparum . Малар. J. 11, 210. DOI: 10.1186 / 1475-2875-11-210
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hasenkamp, S., Russell, K. T., Ullah, I., and Horrocks, P.(2013a). Функциональный анализ 5’-нетранслируемой области транскрипта фосфоглутамазы 2 в Plasmodium falciparum . Acta Trop. 127, 69–74. DOI: 10.1016 / j.actatropica.2013.03.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хасенкамп, С., Сидавей, А., Дивайн, О., Рой, Р., и Хоррокс, П. (2013b). Оценка антималярийной активности препаратов на основе биолюминесценции. Малар. J. 12, 58. DOI: 10.1186 / 1475-2875-12-58
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хельм, С., Lehmann, C., Nagel, A., Stanway, R.R., Horstmann, S., Llinas, M., et al. (2010). Идентификация и характеристика промоторной области, специфичной для стадии печени, малярийного паразита Plasmodium . PLoS ONE 5: e13653. DOI: 10.1371 / journal.pone.0013653
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хоррокс П. и Килби Б. Дж. (1996). Физическое и функциональное картирование сайтов начала транскрипции Plasmodium falciparum ядерного антигена пролиферирующих клеток. Мол. Biochem. Паразитол. 82, 207–215. DOI: 10.1016 / 0166-6851 (96) 02737-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Имаи, Т., Иваваки, Т., Акаи, Р., Сузуе, К., Хираи, М., Танигучи, Т. и др. (2014). Оценка экспериментальной церебральной малярии с использованием индикатора окислительного стресса OKD48 мышей. Внутр. J. Parasitol. 44, 681–685. DOI: 10.1016 / j.ijpara.2014.06.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Каушанский, А., Миколайчак, С.А., Виньяли, М., и Каппе, С.Х. (2014). Человека на мышах: успех и перспективы гуманизированных моделей мышей для паразитарных инфекций малярии человека. Cell Microbiol. 16, 602–611. DOI: 10.1111 / cmi.12277
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хан Т., ван Браммелен А. К., Паркинсон К. Дж. И Хоппе Х. С. (2012). Анализы АТФ и люциферазы для определения скорости действия лекарственного средства в культурах in vitro Plasmodium falciparum . Малар. J. 11, 369. DOI: 10.1186 / 1475-2875-11-369
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Колевзон, Н., Насереддин, А., Наик, С., Явин, Э., Дзиковски, Р. (2014). Использование пептидных нуклеиновых кислот для управления экспрессией генов у малярийного паразита Plasmodium falciparum . PLoS ONE 9: e86802. DOI: 10.1371 / journal.pone.0086802
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лакрю, А.Н., Саенс, Ф. Э., Кросс, Р. М., Удензе, К. О., Монастырский, А., Штейн, С. и др. (2013). 4 (1H) -хинолоны с активностью на стадии печени против Plasmodium berghei . Антимикробный. Агенты. Chemother. 57, 417–424. DOI: 10.1128 / AAC.00793-12
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, К., О’Нил, М., Се, Л., Карида, Д., Цзэн, К., Чжан, Дж. И др. (2014). Оценка профилактической активности и фармакокинетического профиля перорального тафенохина по сравнению с примахином для подавления инфекций, вызванных малярией на стадии печени. Малар. J. 13, 141. DOI: 10.1186 / 1475-2875-13141
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лин, Дж. У., Саджид, М., Рамесар, Дж., Хан, С. М., Джансе, К. Дж., И Франке-Файард, Б. (2013). Скрининг ингибиторов стадий крови P. berghei с использованием биолюминесцентных репортерных паразитов. Methods Mol. Биол. 923, 507–522. DOI: 10.1007 / 978-1-62703-026-7_35
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лопес-Эстраньо, C., Гопалакришнан, А.М., Семблат, Дж. П., Фергус, М. Р., Мазье, Д., и Халдар, К. (2007). Энхансер-подобная область регулирует специфическую для стадии промотора экспрессию гена hrp3 у малярийного паразита человека Plasmodium falciparum . Biochim. Биофиз. Acta 1769, 506–513. DOI: 10.1016 / j.bbaexp.2007.04.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лукантони Л., Даффи С., Аджалли С. Х., Фидок Д. А. и Эйвери В. М. (2013). Идентификация ингибиторов MMV-малярии гаметоцитов Plasmodium falciparum на ранних стадиях с использованием высокопроизводительного анализа на основе люциферазы. Антимикробный. Агенты Chemother. 57, 6050–6062. DOI: 10.1128 / AAC.00870-13
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Люкуми, Э., Дарлинг, К., Джо, Х., Нэппер, А. Д., Чандрамоханадас, Р., Фишер, Н. и др. (2010). Открытие мощных низкомолекулярных ингибиторов мультирезистентности Plasmodium falciparum с использованием нового миниатюрного высокопроизводительного анализа на основе люциферазы. Антимикробный. Агенты Chemother. 54, 3597–3604. DOI: 10.1128 / AAC.00431-10
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Майр, Г. Р., Бракс, Дж. А., Гарвер, Л. С., Вигант, Дж. К., Холл, Н., Диркс, Р. В. и др. (2006). Регуляция полового развития Plasmodium путем репрессии трансляции. Наука 313, 667–669 DOI: 10.1126 / science.1125129
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Манцони, Г., Брике, С., Риско-Кастильо, В., Готье, К., Топчу, С., Иванеску, М. Л. и др. (2014). Быстрая и надежная процедура отбора для создания рекомбинантных малярийных паразитов, не содержащих маркеров. Sci. Реп. 4, 4760. doi: 10.1038 / srep04760
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Марксисин, С. Р., Соуза, Дж. К., Райхард, Г. А., Карида, Д., Зенг, К., Ронкал, Н., и др. (2014). Тафенохин и NPC-1161B требуют метаболизма CYP 2D для противомалярийной активности: последствия для класса 8-аминохинолина противомалярийных соединений. Малар. J. 13, 2. DOI: 10.1186 / 1475-2875-13-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Милителло, К. Т., Додж, М., Бетке, Л., и Вирт, Д. Ф. (2004). Идентификация регуляторных элементов в геноме Plasmodium falciparum . Мол. Biochem. Паразитол. 134, 75–88. DOI: 10.1016 / j.molbiopara.2003.11.004
CrossRef Полный текст | Google Scholar
Миллер, Дж. Л., Мюррей, С., Воган, А. М., Харупа, А., Sack, B., Baldwin, M., et al. (2013). Количественная биолюминесцентная визуализация преэритроцитарной малярийной паразитарной инфекции с использованием экспрессии люциферазы Plasmodium yoelii . PLoS ONE 8: e60820. DOI: 10.1371 / journal.pone.0060820
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Милнер, Д. А. младший, Уиттен, Р. О., Камиза, С., Карр, Р., Лиомба, Г., Дзамалала, К., и др. (2014). Системная патология церебральной малярии у детей Африки. Фронт.Cell Infect Microbiol. 4: 104. DOI: 10.3389 / fcimb.2014.00104
CrossRef Полный текст | Google Scholar
Myrick, A., Munasinghe, A., Patankar, S., and Wirth, D. F. (2003). Картирование гена множественной лекарственной устойчивости Plasmodium falciparum в 5’-верхнем направлении и доказательства индукции уровней транскрипта противомалярийными препаратами у чувствительных к хлорохину паразитов. Мол. Microbiol. 49, 671–683. DOI: 10.1046 / j.1365-2958.2003.03597.x
CrossRef Полный текст | Google Scholar
Мюле, Р.А., Аджаллей С., Фалкард Б., Нкрума Л. Дж., Мюле М. Э. и Фидок Д. А. (2009). Промотор гена var, вовлеченный в тяжелую форму малярии, вызывает молчание ядра и регулируется 3’-нетранслируемой областью и интронными цис-элементами. Внутр. J. Parasitol. 39, 1425–1439. DOI: 10.1016 / j.ijpara.2009.05.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мвакингве А., Тинг Л. М., Хохман С., Чен Дж., Синнис П. и Ким К. (2009). Неинвазивный мониторинг в реальном времени биолюминесцентного развития на стадии печени Plasmodium parasites . J. Infect. Дис. 200, 1470–1478. DOI: 10.1086 / 606115
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Огуарири Р. М., Данн Дж. М. и Голайтли Л. М. (2006). Регуляторные элементы 3 ’гена, необходимые для экспрессии онтогенетического белка Plasmodium falciparum , Pfs25. Мол. Biochem. Паразитол. 146, 163–172. DOI: 10.1016 / j.molbiopara.2005.12.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Озвара, Х., ван дер Вел, А., Кокен, К. Х., и Томас, А. В. (2003). Активность гетерологичного промотора в стабильных и временных трансгенах Plasmodium knowlesi. Мол. Biochem. Паразитол. 130, 61–64. DOI: 10.1016 / S0166-6851 (03) 00141-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Pasini, E.M., Braks, J.A., Fonager, J., Klop, O., Aime, E., Spaccapelo, R., et al. (2013). Протеомные и генетические анализы показывают, что Plasmodium berghei стадий крови экспортируют большой и разнообразный репертуар белков. Мол. Клетка. Протеомика 12, 426–448. DOI: 10.1074 / mcp.M112.021238
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Патакотту Б. Р., Сингх П. К., Малхотра П., Чаухан В. С. и Патанкар С. (2012). Анализ in vivo сайтов инициации трансляции в Plasmodium falciparum . Мол. Биол. Репутация . 39, 2225–2232. DOI: 10.1007 / s11033-011-0971-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Плоемен, И., Behet, M., Nganou-Makamdop, K., van Gemert, G.J., Bijker, E., Hermsen, C., et al. (2011). Оценка иммунитета против малярии с использованием люцифераз-экспрессирующих Plasmodium berghei паразитов. Малар. J. 10, 350. DOI: 10.1186 / 1475-2875-10-350
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ploemen, I.H., Chakravarty, S., van Gemert, G.J., Annoura, T., Khan, S.M., Janse, C.J., et al. (2013). Plasmodium Нагрузка на печень после парентерального введения спорозоитов грызунам. Вакцина. 31, 3410–3416. DOI: 10.1016 / j.vaccine.2012.09.080
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ploemen, I.H., Prudêncio, M., Douradinha, B.G., Ramesar, J., Fonager, J., van Gemert, G.J., et al. (2009). Визуализация и количественный анализ стадии малярии грызунов с помощью визуализации в реальном времени. PLoS ONE 4: e7881. DOI: 10.1371 / journal.pone.0007881
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Портер, М.Э. (2002). Положительные и отрицательные эффекты делеций и мутаций в 5’-фланкирующих последовательностях ДНК-полимеразы Plasmodium falciparum дельта. Мол. Biochem. Паразитол. 122, 9–19. DOI: 10.1016 / S0166-6851 (02) 00064-6
CrossRef Полный текст | Google Scholar
Рамакришнан, К., Радемахер, А., Соичот, Дж., Коста, Г., Уотерс, А. П., Джансе, К. Дж. И др. (2012). Репортерные линии P. berghei , специфичные для слюнных желез, позволяют быстро оценить тканеспецифическую нагрузку спорозоитов у комаров. PLoS ONE. 7: e36376. DOI: 10.1371 / journal.pone.0036376
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ramalhete, C., da Cruz, F. P., Lopes, D., Mulhovo, S., Rosário, V. E., Prudêncio, M., et al. (2011). Тритерпеноиды как ингибиторы эритроцитарной и печеночной стадий инфекций Plasmodium . Bioorg. Med. Chem. 19, 7474–7481. DOI: 10.1016 / j.bmc.2011.10.044.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рамалете, К., да Круз, Ф. П., Мулхово, С., Соуза, И. Дж., Фернандес, М. Х., Пруденсио, М. и др. (2014). Двухстадийные тритерпеноиды африканского лекарственного растения против малярийных паразитов. Bioorg. Med. Chem. 22, 3887–3890. DOI: 10.1016 / j.bmc.2014.06.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сак, Б. К., Миллер, Дж. Л., Воган, А. М., Дуглас, А., Каушанский, А., Миколайчак, С. и др. (2014). Модель для оценки гуморальной защиты от заражения спорозоитом малярии in vivo путем пассивного переноса моноклональных антител и иммунной сыворотки. Заражение. Иммун. 82, 808–817. DOI: 10.1128 / IAI.01249-13
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Спаккапело, Р., Янсе, К. Дж., Катерби, С., Франке-Файяр, Б., Бонилья, Дж. А., Сифард, Л. М. и др. (2010). Plasmepsin 4-дефицитный Plasmodium berghei обладает ослабленной вирулентностью и индуцирует защитный иммунитет против экспериментальной малярии. Am. J. Pathol. 176, 205–217. DOI: 10.2353 / ajpath.2010.0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стоун, W.J., Churcher, T. S., Graumans, W., van Gemert, G.J., Vos, M. W., Lanke, K. H., et al. (2014). Масштабируемая оценка передачи Plasmodium falciparum в стандартном анализе мембранного питания с использованием трансгенных паразитов, экспрессирующих зеленую флуоресцентную протеин-люциферазу. J. Infect. Дис. 210, 1456–1463. DOI: 10.1093 / infdis / jiu271
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Торн Н., Шен М., Леа В. А., Симеонов А., Ловелл С., Auld, D. S., et al. (2012). Люцифераза светлячков в химической биологии: сборник ингибиторов, механистическая оценка хемотипов и предлагаемое использование в качестве репортера. Chem. Биол. 19, 1060–1072. DOI: 10.1016 / j.chembiol.2012.07.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван Рит, Н., Ван де Вайвер, Х., Пьяна, П. П., Ван дер Линден, А. М., и Бюшер, П. (2014). Панель штаммов Trypanosoma brucei , меченных люциферазами с синим и красным смещением, для биолюминесцентной визуализации в моделях инфицирования мышей. PLoS Negl. Троп. Дис. 8: e3054. DOI: 10.1371 / journal.pntd.0003054
PubMed Аннотация | CrossRef Полный текст | Google Scholar
van Schaijk, B.C., Ploemen, I.H., Annoura, T., Vos, M.W., Lander, F., van Gemert, G.J., et al. (2014). Генетически аттенуированная кандидатная вакцина против малярии на основе спорозоитов, дефицитных по гену P. falciparum b 9 / slarp. Элиф 19: 3. DOI: 10.7554 / eLife.03582
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Воан, А.М., Миколайчак, С. А., Камарго, Н., Лакшманан, В., Кеннеди, М., Линднер, С. Е. и др. (2012). Трансгенный штамм Plasmodium falciparum NF54, который экспрессирует GFP-люциферазу на протяжении всего жизненного цикла паразита. Мол. Biochem. Паразитол. 186, 143–147. DOI: 10.1016 / j.molbiopara.2012.10.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Валлер, К. Л., Мюле, Р. А., Урсос, Л. М., Хоррокс, П., Вердье-Пинар, Д., Сидху, А. Б. и др. (2003).Устойчивость к хлорохину модулируется in vitro уровнями экспрессии переносчика устойчивости к хлорохину Plasmodium falciparum . J. Biol. Chem. 278, 33593–33601. DOI: 10.1074 / jbc.M302215200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Weiwer, M., Mulrooney, C., Massi, D., Heidebrecht, R., Wiegand, R., Lukens, A.K, et al. (2011). ML238: Малая противомалярийная молекула уникального структурного класса . Отчеты о зондировании. Bethesda, MD: Программа молекулярных библиотек NIH.
Google Scholar
Уайт, Н. Дж., Пукриттаяками, С., Хиен, Т. Т., Фаиз, М. А., Мокуолу, О. А., и Дондорп, А. М. (2014). Малярия. Ланцет 383, 723–735. DOI: 10.1016 / S0140-6736 (13) 60024-0
CrossRef Полный текст
Zhang, X., Tolzmann, C.A., Melcher, M., Haas, B.J., Gardner, M.J., Smith, J.D., et al. (2011). Идентификация точки ветвления и требования к последовательности для сплайсинга интронов в Plasmodium falciparum . Eukaryot Cell. 10, 1422–1428. DOI: 10.1128 / EC.05193-11
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зузарте-Луис, В., Салес-Диас, Дж., И Мота, М. М. (2014). Простой, чувствительный и количественный анализ биолюминесценции для определения предпатентного периода малярии. Малар. J. 13, 15. DOI: 10.1186 / 1475-2875-13-15
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Полный жизненный цикл малярийного паразита у гуманизированных мышей DRAG
Первая мышь, обеспечивающая размножение паразита и иммунные ответы человека
Малярия — это печально известное смертельное инфекционное заболевание, поражающее миллионы людей в тропических и субтропических регионах мира.О новом важном прогрессе в развитии терапии малярии сообщается в журнале Malaria Journal (Wijayalath et al., 2014). Авторы трансплантировали гемопоэтические стволовые клетки (HSC) пуповинной крови человека иммунодефицитным мышам, экспрессирующим HLA класса II. Помимо установления человеческого иммунитета у мышей, у мышей также развились гепатоциты человека, эндотелиальные клетки, клетки Купфера, ретикулоциты и эритроциты. Все эти популяции клеток человека необходимы для завершения жизненного цикла Plasmodium falciparum , наиболее опасного вида, вызывающего малярию.Мыши с малярийными паразитами в крови сделали возможным передачу паразита комарам и вызвали иммунный ответ, который очистил организм за 45 дней.
Гуманизированные мыши DRAG, которым привиты несколько популяций клеток человека
Мышь DRAG, NOD.Cg- Rag1 tm1Mom Il2rg tm1Wjl Tg (HLA-DRA, HLA-DRB1 * 0401) 39-2Kito / ScasJ (017914), является высоко иммунодефицитным штаммом мышей человека. Аллель HLA класса II, обычно называемый HLA-DR4.Wijayalath et al. трансплантировали мышам DRAG, подвергнутым сублетальной миелоабляции, человеческие HSC CD34 + из пуповинной крови, соответствующие гаплотипу HLA-DR4. Экспрессия HLA-DR4 у мышей вместе с HSC, подобранными по донору, была выбрана для улучшения развития зрелых человеческих Т-клеток и В-клеток, способных к переключению класса иммуноглобулинов, что необходимо для функционального иммунитета человека.
Жизненный цикл P. falciparum требует человеческих красных кровяных телец (эритроцитов). Используя проточную цитометрию, исследователи обнаружили, что гуманизированные мыши DRAG образуют редкие популяции ретикулоцитов и эритроцитов (~ 0.2-1% периферической крови) не менее 4 месяцев.
Другими ключевыми популяциями клеток человека, важными для жизненного цикла паразитов, являются гепатоциты, эндотелиальные клетки и клетки Купфера. Клетки Купфера происходят из миелоидной линии костного мозга и меняются в результате миелоабляции. У трансплантированных мышей человеческие CD45 + CD68 + клетки Купфера составляли ~ 11% от выделенных мононуклеарных клеток печени. Гепатоциты и эндотелиальные клетки не происходят из костного мозга, однако во многих публикациях сообщалось об их присутствии после трансплантации HSC.Доказательства наличия гепатоцитов человека у химерных мышей получены из:
.- Обнаружение трансферрина человека в плазме
- Положительные клетки на трансферрин человека и Hep Par-1 человека (маркер митохондрий гепатоцитов человека) в гистологических срезах печени
- Альбумин-положительные клетки человека, обнаруживаемые с помощью проточной цитометрии
Эндотелиальные клетки человека были также обнаружены у химерных мышей путем мечения срезов печени анти-человеческим CD36. Клетки CD36 + наблюдались в венах печени и синусоидах.
Эти результаты продемонстрировали, что гуманизированные мыши DRAG содержат все популяции клеток, необходимые для жизненного цикла малярийных паразитов (см. Рисунок).
Жизненный цикл малярийных паразитов. Комар вызывает инфекцию при укусе. Сначала спорозоиты попадают в кровоток, затем мигрируют в печень. Они заражают клетки печени, где они размножаются в мерозоиты, разрывают клетки печени и возвращаются в кровоток. Затем мерозоиты заражают красные кровяные тельца, где они развиваются в кольцевые формы, трофозоиты и шизонты, которые, в свою очередь, производят новые мерозоиты.Также образуются половые формы, которые, если их захватит комар, заразят насекомое и продолжат жизненный цикл. (Изображение предоставлено NIAID)
Малярийная паразитемия у гуманизированных мышей
Гуманизированным мышам DRAG внутривенно вводили спорозоитов P. falciparum , а через 5 дней исследовали печень на наличие шизонтов. При гистохимическом окрашивании срезов печени обнаружены шизонты PfHSP70 + . Кроме того, ПЦР для паразитоспецифической 18s рРНК в крови мышей продемонстрировала продолжающуюся паразитемию.Все инфицированные мыши (n = 49) были положительными на инфекцию печени и паразитемию крови. Прогрессирование стадии в крови паразита подтверждено обнаружением колец P. falciparum и трофозоитов в мазках периферической крови, окрашенной по Геймсе. Кроме того, при совместном культивировании крови, инфицированной паразитами, с эритроцитами, авторы обнаружили расширение колец, трофозоитов, шизонтов и гаметоцитов. Эти результаты свидетельствуют о том, что гуманизированные мыши DRAG воспроизводят все in vivo человеческих частей жизненного цикла паразитов.
Для завершения жизненного цикла паразита требуется комар. An. Комарам Stephensi позволяли питаться содержащей гаметоциты кровью in vitro , а в отдельном эксперименте позволяли питаться непосредственно инфицированными мышами. Оба метода кормления привели к инфицированию ооцист и слюнных желез у комаров, а также к обнаружению ооцитов в средней кишке.
В целом, эти результаты ясно демонстрируют полный жизненный цикл паразита с использованием модели комара и мелких животных, полезной в лабораторных условиях.
Иммунные реакции против паразитов
Чтобы платформа гуманизированных мышей была полезной для терапевтических разработок, она должна быть способна к паразитоспецифическим иммунным ответам. Интересно, что инфицированные мыши были способны избавиться от паразита в течение 45 дней. Спленоциты инфицированных мышей DRAG стимулировали in vitro экстрактами эритроцитов, инфицированных паразитами. Проточная цитометрия использовалась для обнаружения активации Т-клеток. Стимулированные человеческие CD4 + и CD8 + Т-клетки продуцировали TNFα, но не INFγ.Гуморальный ответ является ключевым компонентом иммунного очищения паразита, поскольку эритроциты не экспрессируют антиген, необходимый для опосредованного Т-клетками удаления. Поэтому кровь инфицированных мышей исследовали на предмет продукции антител, специфичных для паразитов. Мыши продуцировали антитела IgM и IgG, специфичные к эритроцитам на стадии кольца, трофозоита и шизонта, у 14 из 18 мышей.
Предыдущие платформы мышей для заражения P. falciparum позволяли изучать только печень или эритроцитарный компонент жизненного цикла паразитов.Работа, представленная Wijayalath, et. др., впервые демонстрирует модель мыши, которая включает все компоненты жизненного цикла. Кроме того, эта новая модель поддерживает иммунные функции человека, что позволит лучше понять иммунобиологию малярии, что, в свою очередь, может позволить разработать новые стратегии лечения.
.