Pityrosporum orbiculare — это… Что такое Pityrosporum orbiculare?
Pityrosporum — A genus of fungi of disputed pathogenicity found in dandruff and seborrheic dermatitis. [G. pityron, bran, + sporos, seed] P. orbiculare SYN: Malassezia furfur. P. ovale SYN: Malassezia furfur. * * * Pit·y·ros·po·rum .pit ə räs pə rəm; .pit ər ō… … Medical dictionary
Pityrosporum — n. a genus of yeasts, now renamed Malassezia, producing superficial infections of the skin. The species P. orbiculare is a normal inhabitant of the scalp but it can become pathogenic in susceptible individuals (see pityriasis) … The new mediacal dictionary
Malassezia — A genus of fungi (family Cryptococcaceae) of low pathogenicity that lack the ability to synthesize medium chain and long chain fatty acid s and require an exogenous supply of these lipids for growth as can be found in the skin. [L. C. Malassez] M … Medical dictionary
Залаин — Действующее вещество ›› Сертаконазол* (Sertaconazole*) Латинское название Zalain АТХ: ›› D01AC14 Сертаконазол Фармакологическая группа: Противогрибковые средства Нозологическая классификация (МКБ 10) ›› B35.
Тербикс — Действующее вещество ›› Тербинафин* (Terbinafine*) Латинское название Terbix АТХ: ›› D01AE15 Тербинафин Фармакологическая группа: Противогрибковые средства Нозологическая классификация (МКБ 10) ›› B35 Дерматофития ›› B35.3 Микоз стоп ›› B35.6… … Словарь медицинских препаратов
Себорейный дерматит — В этой статье не хватает ссылок на источники информации. Информация должна быть проверяема, иначе она может быть поставлена под сомнение и удалена. Вы можете … Википедия
Фунготербин — Действующее вещество ›› Тербинафин* (Terbinafine*) Латинское название Fungoterbine АТХ: ›› D01AE15 Тербинафин Фармакологическая группа: Противогрибковые средства Нозологическая классификация (МКБ 10) ›› B35 Дерматофития ›› B35.0 Микоз бороды и… … Словарь медицинских препаратов
Кето плюс — Действующее вещество ›› Кетоконазол* + Пиритион цинк* (Ketoconazole* + Pyrithione zinc*) Латинское название Keto plus АТХ: ›› D01AE Прочие противогрибковые препараты для местного применения Фармакологическая группа: Противогрибковые средства в… … Словарь медицинских препаратов
Отрубевидный лишай — Отрубевидный (разноцветный) лишай … Википедия
Лишай отрубевидный — I Лишай отрубевидный (pityriasis furfuracea; синоним лишай разноцветный) грибковое заболевание кожи, характеризующееся поражением рогового слоя эпидермиса.
Лечение за рубежом
Частный кабинет доктора Херинга находится с самом центре Мюнхена. Дерматологический кабинет оснащен всем необходимым для амбулаторного приема пациентов. Основная задача кабинета – диагностика, лечение и профилактика кожных заболеваний.
<p><b>В кабинете доктора Херинга профессионально диагностируют и лечат следующие заболевания:</b></p>
</br>
<ul>
<li>атопический дерматит аллергическое заболевание кожи</li>
<li>аллергологическая диагностика</li>
<li>бородавки — доброкачественное новообразование (наросты) на коже</li>
<li>витилиго — нарушение пигментации кожи</li>
<li>экзема — воспалительное заболевание кожи, сопровождающееся сыпью и склонностью к рецидивам</li>
<li>микозы — заболевание кожи и ногтей, вызванное паразитарными грибами</li>
<li>псориаз — хроническое неинфекционное заболевание, поражающее кожу</li>
<li>рак кожи и меланомы — злокачественные опухли</li>
<li>разноцветный лишай — грибковое заболевание кожи, вызванное грибом pityrosporum orbiculare</li>
<li>розовые угри — патология сальных желез</li>
<li>склеродермия — заболевание, связанное с уплотнением кожи</li>
<li>cеборейный дерматит — воспалительное заболевание, которое поражает участки кожи, где наиболее сосредоточены сальные железы</li>
</ul>
</br>
<p><b>Членство в:</b></p>
</br>
<ul>
<li>Немецком дерматологическом обществе</li>
</ul>
Как произносится pityrosporum orbiculare | HowToPronounce.
 com
com0 рейтинг рейтинг рейтинги
Запишите и прослушайте произношение
Вы можете произносить это слово лучше
или произносить в разных акцента ?
Добавить детали pityrosporum orbiculare
Значения для pityrosporum orbiculareДобавить определение
Отмена showDefinitionsForm»>Спасибо за помощь
showDefinitionsForm»>Спасибо за помощь вы не авторизованы..
Пожалуйста Войти в или регистр или пост в качестве гостя
Разместить
Фонетическое написание pityrosporum orbiculareДобавить фонетическое написание
Отмена
вы не авторизованы..
Пожалуйста Войти в или регистр или пост в качестве гостя
Разместить
Синонимы для pityrosporum orbiculare Добавить синонимы Отмена showSynonymsForm»>Спасибо за помощь
showSynonymsForm»>Спасибо за помощьвы не авторизованы..
Пожалуйста Войти в или регистр или пост в качестве гостя
Разместить
Антонимы к pityrosporum orbiculareДобавить антонимы
Отмена showAntonymsForm»>Спасибо за помощь
showAntonymsForm»>Спасибо за помощьвы не авторизованы..
Пожалуйста Войти в или регистр или пост в качестве гостя
Разместить
Примеры pityrosporum orbiculare в предложенииДобавить предложение
Отмена sentenceForm»>Спасибо за помощь
sentenceForm»>Спасибо за помощьpityrosporum orbiculare должен быть в предложении
вы не авторизованы..
Пожалуйста Войти в или регистр или пост в качестве гостя
Разместить
Переводы pityrosporum orbiculareДобавить перевод
Отмена tranlsationForm»>Спасибо за помощь
tranlsationForm»>Спасибо за помощьВыбрать языкАфрикаансАлбанскийАмхарскийАрабский языкАрменииazerbaijanбашкирскийБаскскийБелорусскийБенгальскийБосниецbretonБолгарскийбирманскийКаталанскийКитайскийХорватскийЧешскаяДатскийГолландскийАнглийскийЭсперантоЭстонскийfilipinoФинскийФранцузскийГалицкаягрузинскийНемецкийГреческийГуджаратиhaitian HawaiianИвритХиндиВенгерскийИсландскийИндонезийскийИрландскийИтальянскийЯпонскийЯванскийКаннадаКазахКхмерскийКорейскийКиргизскийЛаоскаЛатинскийЛатвииЛитовскийЛюксембургскийМакедонскийМалагасиецMалайскийMalayalamМальтийскийmandarin-chineseМаориМаратхиМонгольскийНепальскийНорвежскийПерсидскийПольскийПортугальскийпанджабиРумынскийРусскийГэльскийСербскийSinhalaСловацкийCловенскийИспанскийСуданскийСуахилиШведскийтагальскогоТаджикскийТамилататаринТелугутайскийТурецкийУкраинецУрдуузбекВьетнамскийВаллийскийXhosaИдиш
вы не авторизованы.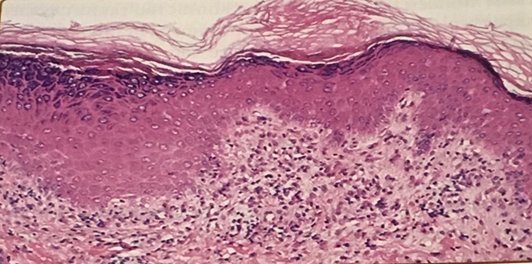 .
.
Пожалуйста Войти в или регистр или пост в качестве гостя
Разместить
Лечение отрубевидного (разноцветного) лишая в Житомире
Отрубевидный (разноцветный) лишай это заболевание грибковой породы, которое поражает роговой слой эпидермиса. Возбудителем лишая является липофильный дрожжевой грибок Питироспорум Орбикуларе (Pityrosporum Orbiculare), который размножается в роговом слое эпидермиса и может передаваться людям со сниженным местным иммунитетом и специфическим метаболизмом кожи.
ПРИЧИНЫ РАЗНОЦВЕТНОГО ЛИШАЯСледующие условия способствуют развитию заболевания:
параллельные заболевания иммунной, пищеварительной или эндокринной системы (сахарного диабета, бронхиальной астмы, пневмонии, ожирения, и т.
 д.)
д.)ношение тесной синтетической одежды;
генетическая предрасположенность к дерматомикозам;
период менопаузы;
чрезмерная потливость, которая приводит к изменениям химического состава пота и усиленному размножению грибка;
длительный прием медикаментов.
 д.)
д.)Отрубевидный лишай появляется на теле человека, если его кожа становится благоприятной средой для грибка, наступает активное его размножение, которое нарушает работу клеток, вырабатывающих пигмент меланин. Характерная симптоматика для этого заболевания:
появление небольших пятен белого, розового и светло-коричневого цвета, которые постепенно меняют свой окрас и становятся красными, темно-бурыми, а потом коричневыми (отсюда и произошло второе название лишая «разноцветный»), которые имеют четкие края, могут занимать значительные участки кожи;
очень часто сопровождается отрубевидным шелушением;
иногда возможно появление зуда;
в некоторых случаях возникает чрезмерная потливость.

Отрубевидный лишай появляется преимущественно на верхней части тела — грудь, спина, живот, шея, волосистая часть головы, чаще всего к заболеванию склонны молодые мужчины и подростки.
ДИАГНОСТИКА И ЛЕЧЕНИЕ ОТРУБЕВИДНОГО ЛИШАЯДиагностировать заболевание без консультации врача практически невозможно, поскольку симптоматика дерматологических заболеваний зачастую очень схожа.
Квалифицированный врач дерматолог клиники «Оксфорд Медикал» проведет осмотр кожных покровов, назначит соскоб кожи на выявления грибка-возбудителя. И только на основании полной клинической картины и при наличии классических симптомов отрубевидного лишая поставит правильный диагноз и назначит лечение.
Лечение этого заболевания включает в себя:
наружную терапию, это обработка кожи противогрибковыми препаратами системного или местного действия: мазями, кремами и растворами;
дезинфекция вещей пациента (одежды, нательного белья, головных уборов и предметов личной гигиены), эта процедура является обязательной.

Терапию отрубевидного лишая в обязательном порядке должен контролировать дерматолог, так как самостоятельное лечение, к которому прибегают пациенты, зачастую неэффективно и может привести к рецидиву.
В клинике «Оксфорд Медикал Житомир» вам помогут быстро и эффективно избавиться от отрубевидного лишая.
ЗАПИСАТЬСЯ НА ПРИЕМ
Дерматиты и роль бактерии Лептотрихии в их возникновении
Себорейный и пероральный дерматиты — одна из актуальных проблем дерматокосметологии.
Врачи отмечают, что количество пациентов с этими заболеваниями, которые которые проявляются значительным косметическим дефектом кожи в виде жирного блеска, шелушения и высыпаний, сопровождаемых зудом, постоянно растет.
Почему возникает дерматит? Как он дает о себе знать? И что делать, чтобы справиться с этим состоянием? На вопросы отвечает врач-дерматовенеролог (взрослый), дерматовенеролог (детский), косметолог СЦНТ «НОВОСТОМ» Мусаелова Наталья Владимировна.
Что, когда, зачем и почему
В возникновении себорейного дерматита в последние годы была доказана роль бактерии Leptotrichia bucalis.
В основе себорейного дерматита лежит хроническое воспаление кожи, связанное повышением количества и изменением качества кожного сала с локализацией на тех участках кожи головы и туловища, где развиты сальные железы -волосистая часть головы, бровей, ресниц, носощечных складок, ушей, заушных пространств, передней области груди , межлопаточной и паховой областей.
Поражении кожи волосистой части головы наблюдается в виде появления перхоти, чешуек .корочек , истончением и поредением волос, очень часто сопровождаемые зудом и раздражением Нередко можно видеть желтоватые чешуйко-корки и геморрагические корки, образовавшиеся в результате расчесов. Часто в процесс вовлекается кожа лба, заушных областей, области ушных проходов.
На гладкой коже лица, спины, груди появляются очаги покраснения, папуло- бляшки. Очень часто в возникновении этих дерматитов можно проследить генетическую предрасположенность, стрессы, заболевания ЖКТ, гормональные сбои и влияние внешних и климатических факторов.
Ведущую роль в патогенезе себорейного дерматита волосистой части головы и перхоти отводят возбудителям-дрожжеподобным грибам.
Эти грибы являются постоянным компонентом микрофлоры здоровой кожи у более чем 90% населения. Они используют секрет сальных желёз для своего роста и развития .Разновидность Malassezia futur чаще встречается на волосистой части головы. Pityrosporum orbiculare — на коже туловища и лица. Стрессовые ситуации ухудшают течение себорейного и перорального дерматитов, вследствие изменения гормонального фона. У мужчин 40-60 лет себорейный дерматит наблюдается чаще, чем у женщин, возможно это связано со снижением мужского гормона- тестостерона.
Таким образом, вышеназванные причины приводят к распространению и увеличению количества ранее сапрофитных микроорганизмов и развитию очагов воспаления с нарушением иммунного кожного ответа.
В последние годы усиливается интерес к анаэробным микроорганизмам, а именно к бактериям рода Leptothrichia u Leptothrix.
Одна из разновидностей -Leptotrichia bucalis, обитатель ротовой полости человека. При микроскопии Лептотрихии имеют вид прямых или слегка изогнутых палочек диаметром 1,0 – 1,5 мкм с заострёнными или закруглёнными концами. При длительном паразитировании лептотрихий на слизистых оболочках полости рта и половых органов лептотрихозная инфекция приводит к общему снижению резистентности макроорганизма, что является пусковым фактором в развитии воспалительных реакций на коже и слизистых
Для диагностики изучают соскобы из очагов поражения. Основным критерием диагностики лептотрихозного поражения является обнаружение возбудителя (10-15 бактерий во всех полях зрения) с наличием элементов воспаления (лейкоциты, гистиоциты). Без отсутствия элементов воспаления, следует рассматривать бактерионосительство.
Поражению подвергаются преимущественно миндалины, боковая и задняя часть глотки, язык. Лептотрихоз – псевдомикотическое поражение слизистой оболочки рта. Объективно: на слизистой ротовой полости, языка, дужек мягкого неба появляются серовато-белесоватый налет с переходом в светло-коричневый, коричневый цвет, который не снимается свободно шпателем.
Диагноз заболевания выставляется на основании характерных клинических данных и микроскопической картины препарата.
При микроскопии в соскобах со слизистой оболочки твердого неба обнаруживаются споры и псевдомицелий рода Candida с языка лептотрихии с элементами воспаления. Поэтому следует проводить правильную интерпретацию бактериоскопических препаратов, учитывая, что лечение кандидоза и лептотрихоза — неодинаковое.
Лечение дерматита
Исходя из результатов проведенных микроскопических исследований и исследовании иммунного статуса, клинической картины больных себорейным, пероральным дерматитом и лептотрихозом, необходимо использовать комплексную терапию.
Для лечения слизистых используют бактериофаги (стафилококковый, комплексный) и растворы антисептиков (Бетадин и Мирамистин) путем аппликаций на язык или для полоскания полости рта, а также противомикотическую (Орунгал, Ламизил) и противопаразитарную (Трихопол, Декарис) терапию внутрь.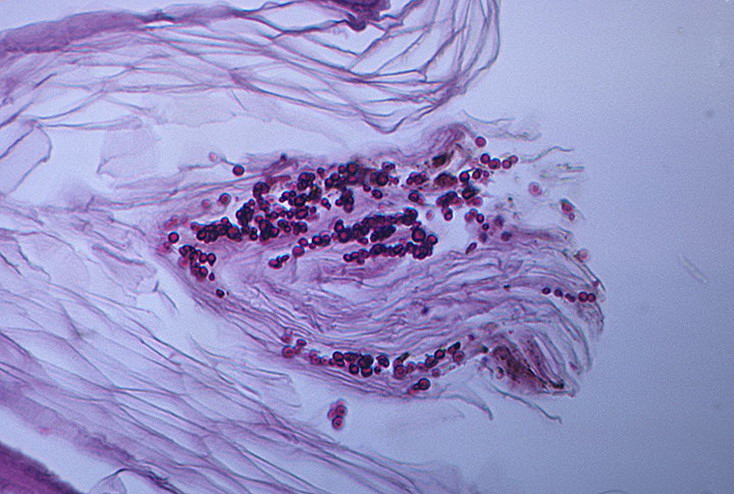
Для лечения воспаления кожи используются в сочетании антибактериальные (Метрогил) и противогрибковые (Низорал, Тридерм, Акридерм ГК и др.) препараты.
Источник
LOINC 10949-6 — Malassezia furfur IgE Ab [Units/volume] in Serum
Term Description
Pityrosporum orbiculare is an obsolete synonym of Malassezia furfur
Source: Regenstrief LOINC
Part Description
LP14960-6 Malassezia furfur
Malassezia (formerly known as Pityrosporum) is a genus of related fungi, classified as yeasts, naturally found on the skin surfaces of many animals including humans. It can cause hypopigmentation on the trunk and other locations in humans if it becomes an opportunistic infection. Copyright Text is available under the Creative Commons Attribution/Share-Alike License. See http://creativecommons.org/licenses/by-sa/3.0/ for details. Source: Wikipedia, Wikipedia
Source: Wikipedia, Wikipedia
Fully-Specified Name
- Component
- Malassezia furfur Ab.IgE
- Property
- ACnc
- Time
- Pt
- System
- Ser
- Scale
- Qn
- Method
Additional Names
- Short Name
- M furfur IgE Qn
- Display Name
- Malassezia furfur IgE Qn (S)
- Consumer Name Alpha
- Malassezia furfur allergy test
Basic Attributes
- Class
- ALLERGY
- Type
- Laboratory
- First Released
- Version 1.0j-a
- Last Updated
- Version 2.42
- Order vs. Observation
- Both
Language Variants
Get Info- zh-CNChinese (China)
- 糠秕马拉色菌 抗体.IgE:
任意型浓度: 时间点: 血清: 定量型: - nl-NLDutch (Netherlands)
- Malassezia furfur As.IgE:
eenheid/volume: moment: serum: kwantitatief: - et-EEEstonian (Estonia)
- Malassezia furfur antikehad.IgE:
Acnc: Pt: S: Qn: - fr-BEFrench (Belgium)
- Malassezia furfur Anticorps.IgE:
Concentration arbitraire: Temps ponctuel: Sérum: Quantitatif: - fr-CAFrench (Canada)
- Malassezia furfur , IgE:
Concentration arbitraire: Temps ponctuel: Sérum: Quantitatif: - fr-FRFrench (France)
- Malassezia furfur Ac IgE:
Arbitraire/Volume: Ponctuel: Sérum: Numérique: - it-ITItalian (Italy)
- Malassezia furfur , Ab.IgE:
ACnc: Pt: Siero: Qn: - pt-BRPortuguese (Brazil)
- Malassezia furfur Ac.IgE:
ACnc: Pt: Soro: Qn: - ru-RURussian (Russian Federation)
- Malassezia furfur Ат.IgE:
ПроизвКонц: ТчкВрм: Сыв: Колич: - es-ARSpanish (Argentina)
- anticuerp contra Malassezia furfur.inmunoglobulina E:
concentración arbitraria: punto en el tiempo: suero: cuantitativo: - es-ESSpanish (Spain)
- Malassezia furfur IgE:
Concentración arbitraria: Punto temporal: Suero: Qn: - tr-TRTurkish (Turkey)
- Malassezia furfur Ab.IgE:
SçKons: Zmlı: Ser: Kant:
Related Names
- ABS
- Aby
- Allergen
- Allergens
- ALLERGY TESTING
- Antby
- Anti
- Antibodies
- Antibody
- Arbitrary concentration
- Autoantibodies
- Autoantibody
- Immune globulin E
- Immunoglobulin E
- M furfur
- m70
- Pityrosporum orbiculare
- Point in time
- QNT
- Quan
- Quant
- Quantitative
- Random
- Serum
- SR
Example Units
| Unit | Source |
|---|---|
| k[IU]/L | Example UCUM Units |
LOINC FHIR
® API Example — CodeSystem Request Get Infohttps://fhir.loinc.org/CodeSystem/$lookup?system=http://loinc.org&code=10949-6LOINC Copyright
Copyright © 2021 Regenstrief Institute, Inc. All Rights Reserved. To the extent included herein, the LOINC table and LOINC codes are copyright © 1995-2021, Regenstrief Institute, Inc. and the Logical Observation Identifiers Names and Codes (LOINC) Committee. See https://
Разноцветный лишай и Pityrosporum orbiculare: микологическое исследование | Медицинская микология
Аннотация
Принято считать, что причиной разноцветного лишая является диморфный липофильный гриб Pityrosporum orbiculare . В этом исследовании грибок культивировали от всех пациентов с разноцветным лишаем. Он также был изолирован от нормально выглядящей кожи и, по-видимому, излечивал поражения у значительной части пациентов с разноцветным лишаем; от пациентов с себорейным дерматитом и от здоровых добровольцев.
Выделение P. orbiculare наиболее удовлетворительно было достигнуто на среде пептон-глюкоза-дрожжевой экстракт, содержащей моностеарат глицерина, покрытой оливковым маслом. Пробирки для зародышей были получены при инкубации P. orbiculare в атмосфере, содержащей воздух с 7% CO 2 . Культуры P. orbiculare и P. ovale не проявляли флуоресценции в свете Вуда.
Резюме
II является общим элементом, содержащим агент разноцветного шампиньона и липофильных шампиньонов Pityrosporum orbiculare .
Dans lá présente étude le шампиньонов и été изоляты tous les malades atteints de tinea versicolor. II également été изолирующий де-ла-peau à внешний вид нормальный и d’affections, одежда guéries d’un grand nombre de malades atteints de tinea versicolor; de plus il a été изол де malades суфрант дерматит séborrhéique et des volontaires sains.
L’isolement de P. orbiculare se fait aisément sur Solide de Sabouraud contenant du monostérate de glycérol et recouvert d’huile d’olive.Рудиментарные волокна производятся в твердой среде глюкозы Сабуро в атмосфере 7% CO 2 . Культуры P. orbiculare и P. ovale , экспозиции в люмьер-де-Вуд, без флуоресценции.
Этот контент доступен только в формате PDF.
© 1979 Международное общество микологии человека и животных
кожных заболеваний, связанных с дрожжами Malassezia
Введение
Malassezia — распространенные липидозависимые грибы, которые растут на сальных участках кожи человека, включая лицо, кожу головы и верхнюю часть туловища.Хотя Malassezia являются частью нормальной кожной флоры человека, они также могут вызывать или обострять некоторые кожные заболевания, включая разноцветный лишай, фолликулит Pityrosporum и себорейный дерматит. Противогрибковые препараты местного действия являются основой лечения заболеваний, связанных с Malassezia . Для предотвращения рецидивов часто требуется хроническая профилактика.
Malassezia — это липофильные (липидозависимые) грибы, которые составляют часть нормальной кожной флоры человека, но теперь также признано, что они играют роль в кожных заболеваниях.Было ясно показано, что Malassezia является возбудителем опоясывающего лишая и Pityrosporum фолликулита и, вероятно, играет роль в себорейном дерматите (Gupta, Batra, Bluhm, Boekhout, & Dawson, 2004). Доказательства их участия в других кожных заболеваниях, включая атопический дерматит, псориаз и головной пустулез новорожденных, слабее.
Дрожжи Malassezia были впервые описаны в XIX веке как почкующиеся дрожжи, обнаруженные на коже пациентов с перхотью.Они названы в честь Луи-Шарля Малассеса, французского ученого, который идентифицировал дрожжи во внешнем слое эпидермиса пациентов с себорейным дерматитом. Таксономия Malassezia была несколько запутанной, потому что название рода Pityrosporum исторически использовалось для дрожжевых форм, тогда как название Malassezia было зарезервировано для гифальных (нитчатых) форм, и только после того, как стали применяться молекулярные методы определения видов. В 1980-х годах было обнаружено, что Malassezia и Pityrosporum идентичны.
В настоящее время идентифицировано 11 видов из Malassezia (Gueho, Midgley, & Guillot, 1996; Sugita et al., 2002; Sugita et al., 2004). Наиболее важными с клинической точки зрения являются M. globosa, M. sympodialis, M. restricta, M. obtusa, и M. furfur . Различные виды Malassezia различаются на основе их морфологии, характеристик роста, активности ферментов, а также молекулярных методов (Gemmer, DeAngelis, Theelen, Boekhout, & Dawson, 2002). Malassezia — диморфные грибы, иногда принимающие дрожжевые формы, а иногда гифальные, в зависимости от условий культивирования. В своей дрожжевой форме они могут быть сферическими, овальными или удлиненными, и они размножаются за счет однополярного почкования. Названия Pityrosporum ovale и Pityrosporum orbiculare все еще иногда используются для обозначения дрожжевых форм, но правильнее обозначать как дрожжевые, так и гифальные формы, используя названия видов Malassezia .
Pityrosporum orbiculare — Phadia — Устанавливая стандарты
Код: m70
Латинское название: Pityrosporum orbiculare
Исходный материал: Дрожжевые клетки
Инфекции, вызываемые Pityrosporum , возникают в волосяных фолликулах
Воздействие аллергенов
Pityrosporum представляет собой липофильную дрожжевую форму Malassezia furfur, который обычно считается непатогенным или более сапрофитным чем у 80% здоровых взрослых, но редко встречается у маленьких детей.Инфекции, вызываемые Pityrosporum , возникают в волосяных фолликулах.
Клинический опыт
По-видимому, недавно была описана тесная связь между сенсибилизацией IgE к P. orbiculare и тяжелой атопической экземой (1). Эти авторы обнаружили тесную связь между атопической экземой и появлением антител IgE к плесени и пришли к выводу, что эти антитела могут иметь патогенетическое значение, по крайней мере, у некоторых пациентов с атопической экземой.
Kieffer et al.(2) обнаружили IgE-антитела к Pityrosporum у пациентов с атопическим дерматитом (AD), но не обнаружили доказательств немедленной или замедленной гиперчувствительности при себорейном дерматите. Rokugo et al. (3) также обнаружили положительные пробы на экстракт Porbiculare у 75% пациентов с БА.
Review
Pityrosporum представляет собой липофильную дрожжевую форму Malassezia furfur, обычно считающуюся непатогенным сапрофитом и обнаруживаемую в коже более 80% здоровых взрослых, но редко встречающуюся у молодых. дети.Инфекции, вызываемые Pityrosporum , возникают в волосяных фолликулах.
По-видимому, недавно была описана тесная связь между сенсибилизацией IgE к P. orbiculare и тяжелой атопической экземой (1). Эти авторы обнаружили тесную связь между атопической экземой и появлением антител IgE к плесени и пришли к выводу, что эти антитела могут иметь патогенетическое значение, по крайней мере, у некоторых пациентов с атопической экземой. Киффер и др. (2) обнаружили IgE-антитела к Pityrosporum у пациентов с атопическим дерматитом (AD), но не обнаружили доказательств немедленной или замедленной гиперчувствительности при себорейном дерматите.Rokugo et al. (3) также обнаружили положительные пробы на экстракт Porbiculare у 75% пациентов с БА. Они пришли к выводу, что плесень играет роль аллергена, происходящего из окружающей среды хозяина, в обострении кожных повреждений AD.
Список литературы
- Nordvall LS, Johansson S. IgE-антитела к Pityrosporum orbiculate у детей с атопическими заболеваниями. Acta Paediatr Scand 1990; 70 (3): 343-8.
- Киффер М., Бергбрант И.М., Фаергеманн Дж., Джемек Г.Б., Оттевангер В., Шталь Сков Б., Свейгаард Э.Иммунные реакции на Pityrosporum ovale у взрослых пациентов с атопическим и себорейным дерматитом. J Am Acad Dermatol 1990; 22 (5, Pt 1): 739-42.
- Рокуго М., Тагами Н., Усуба Ю., Томита Ю. Контактная чувствительность к Pityrosporum ovale у пациентов с атопическим дерматитом. Arch Dermatol 1990; 126 (5): 627-32.
Положительные реакции на атопический пластырь на Pityrosporum orbiculare у пациентов с атопическим дерматитом
Hefen der Gattung Malassezia (M. furfur, M. sympodialis, M.globosa, M. obtusa, M. restricta, M. slooffiae, M. pachydermatis) sind ubiquitär auftretende Hefen. Mit Ausnahme von M. pachydermatis besteht eine Abhängigkeit von der Versorgung mit längerkettigen Fettsäuren, so dass Malassezia-Hefen zu den wenigen Obligat lipophilen Mikroorganismen gehören. Aufgrund ihrer Lipophilie lassen sie sich besonders in den talgdrüsenreichen Körperarealen nachweisen. Da sich im Bereich der Glans penis freie Talgdrüsen befinden, sogenannte Tyson`sche Drüsen, stellte sich die Frage, ob auch hier Malassezia-Hefen regelhaft anzutreffen sind.Zu Erkrankungen der Vulva durch Malassezia spp. gibt es Einzelberichte, ebenso hinsichtlich des Auftretens der Pityriasis versicolor, der häufigsten durch Malassezia verursachten Erkrankung, im Bereich der Glans penis oder des Penisschaftes. Über eine Mitbeteiligung der Malassezia-Hefen an der residencenten Flora der Glans penis bzw. des Präputialraumes wurden bisher in der Literatur keine Angaben gemacht, ebenso fehlte eine Aussage bezüglich des Vorkommens im Bereich des hinteren Vaginalgewölbes.In dieser Arbeit wurden Vorkommen, Häufigkeit und Artenspektrum von lipophilen Malassezia-Hefen bei gesunden, in der Mehrzahl nicht zirkumzidierten Männern untersucht. Dabei sollten auch die Standardisolationsmedien zur Anzucht von Malassezia-Hefen, mDixon und Leeming- Notman-Agar, hinsichtlich Anzuchthäufigkeit und isoliertem Erregerspektrum verglichen werden. Ebenso sollte auf die mögliche Beteiligung der Hefen auf genitale Erkrankungen eingegangen werden. Nach 10 tägiger Inkubation bei 32 ° C wurden die Malassezia- und Candida-Hefen nach mikromorphologischen und biochemischen Kriterien Differenziert.Nahezu jeder zweite Proband der untersuchten 130 Männer wies eine Besiedlung des Präputialraumes mit Malassezia spp. auf, wobei am häufigsten die Spezies M. sympodialis und M. globosa nachgewiesen werden konnten, die auch zu den häufigsten Malassezia-Hefen auf der übrigen menschlichen Haut gehören. M. pachydermatis konnte nur в 2 Fällen nachgewiesen werden. M. furfur, der möglicherweise bedeutsam in der Pathogenese der Pityriasis versicolor ist, wurde nur bei einem Probanden isoliert. Aufgrund der Untersuchung ist anzunehmen, dass Malassezia-Hefen möglicherweise Bestandteil der residencenten Mikroflora der Haut im Bereich der Glans penis sind.Der Nachweis der Malassezia-Hefen im Bereich der Glans penis könnte die Bedeutung an der Mitbeteiligung zahlreicher Erkrankungen des äußeren Genitale vermuten lassen, so etwa bei der Balanoposthitis, Dem seborrhoischen der Balanoposthitis, Dem seborrhoischemischen der Balanoposthitis, Dem seborrhoischen der Genitale der Follikulitis, der Tysonitis und dem atopischen Ekzem mit Beteiligung des äußeren Genitale. Bezüglich der Anzüchtung auf den beiden zur Anzüchtung von Malassezia – Hefen empfohlenen Standardmedien konnten keine auffälligen Unterschiede in der Differenzierung nachgewiesen werden.Die erhaltenen Werte für die Kolonisation mit Candida spp. sind mit denen der Literatur vergleichbar, ebenso das Spektrum der Differenzierten Candida-Hefen. Um die Rolle von M. spp. in der Physiologie und Pathophysiologie der Genitalregion zu bestimmen, ist die Anwendung von lipidhaltigem Medium bei Männern mit rezidivierenden und Potentiell Infektiösen Erkrankungen des äußeren Genitale zu empfehlen. Wird wie in dieser Untersuchung auf Cycloheximid verzichtet, so kann auch die Candida-Flora miterfasst werden.Eine Diagnosestellung durch Pilznachweis ist unerlässlich, wenn nicht unnötige Therapieversager in Kauf genommen werden sollen, da alline schon die Standortflora des Präputialraumes eine hohe Komplexität und Inkonstanz aufweist. Die exakte Diagnose ist также die Voraussetzung für eine gezielte Behandlung. Bei 100 Frauen konnten mittels Vaginalabstrich aus dem hinteren Scheidengewölbe keine Malassezia – Hefen nachgewiesen werden, wobei wohl auch der vaginale pH (~ 4,5) eine Rolle spielt, da er unterhalb des Wachstumf? eine Rolle spielen.28% der Frauen wiesen, wie schon aus früheren Studien zu erwarten war, eine Candida- Besiedlung auf, wobei der größte Anteil (> 2/3) wie ebenfalls zu erwarten auf C. albicans fiel. Bei Frauen mit klinischer Symptomatik im Bereich der Vulva, beispielsweise bei vulvären Dystrophien oder einer unklaren Schuppung im Bereich der Vulva ist aber zumindest an die Möglichkeit einer Malassezia Besiediner Oversiedlung im Pesisiedlungenor de Pésisiedlungiseneris de la de la Pésisiedlungeneris coli de la de la de la de la Möglichkeit einer Malassezia Besiediner Oversiedlung. Дрожжи рода Malassezia (M.furfur, M. sympodialis, M. globosa, M. obtusa, M. restricta, M. slooffiae, M. pachydermatis) распространены повсеместно. За исключением M. pachydermatis, они зависят от поступления длинноцепочечных жирных кислот и, таким образом, относятся к немногим липофильным микроорганизмам. Дрожжи Malassezia в основном встречаются в себорейных участках тела. В связи с этим возник вопрос, можно ли их продемонстрировать в области головки полового члена, богатой свободными сальными железами (железы Тайсона). Только единичные наблюдения заболеваний вульвы, вызванных Malassezia spp.Сообщалось также о разноцветном лишайе — наиболее ассоциированном с малассезией заболевании — на головке полового члена и влагалище полового члена. В литературе пока нет доказательств того, что дрожжи Malassezia являются частью резидентной флоры головки полового члена и препуциального пространства, а также в области заднего свода влагалища. Настоящее исследование было направлено на оценку этих вопросов и изучило наличие, частоту и спектр липофильных дрожжей Malassezia в области головки полового члена и препуциального пространства у здоровых мужчин, большинство из которых не были обрезаны.В то же время стандартные среды для выделения культуры Malassezia, модифицированный агар Диксона (mDixon) и агар Лиминга-Нотмана, сравнивали с точки зрения частоты культивирования и патогенного спектра. Кроме того, обсуждалось возможное участие этих дрожжей в заболеваниях половых органов. После 10 дней инкубации при 32 ° C дрожжи Malassezia и Candida были дифференцированы по микроморфологическим и биохимическим критериям. Колонизация Malassezia spp. был обнаружен почти у 50% из 130 исследованных мужчин, M.sympodialis и M. globosa, которые также наиболее распространены в других областях жизни человека. M. pachydermatis обнаружен в двух случаях. M. furfur, который может играть важную роль в патогенезе разноцветного лишая, был выделен только у одного человека. Из исследования можно предположить, что дрожжи Malassezia являются частью резидентной микрофлоры головки полового члена. Их демонстрация в области головки полового члена может свидетельствовать о том, что они также вовлечены в многочисленные заболевания половых органов, например баланопостит, себорейная экзема, псориаз головки полового члена, красный плоский лишай, фолликулит, тизонит и атопическая экзема.Не было обнаружено значительных различий между двумя стандартными средами, используемыми для культивирования дрожжей Malassezia. Результаты колонизации Candida в препуциальной области здоровых мужчин были сопоставимы с данными, опубликованными в литературе; то же самое и со спектром. Чтобы определить роль Malassezia spp. в физиологии и патофизиологии области гениталий липидсодержащие среды рекомендуются мужчинам с рецидивирующими и потенциально инфекционными заболеваниями наружных половых органов. Исключение циклогексимида, как в настоящем исследовании, также позволит обнаружить виды Candida.Поскольку резидентная флора препуциального пространства очень сложна и непостоянна, диагноз должен быть подтвержден демонстрацией грибка, чтобы избежать ненужных неудач лечения. Точный диагноз — основа объективной терапии. В исследовании с участием 100 женщин не удалось обнаружить дрожжевые грибки Malassezia в мазках из заднего свода влагалища. Вагинальный pH (~ 4,5) ниже, чем требуется для оптимального роста Malassezia, и, возможно, другие местные факторы ответственны за отрицательный результат посева.Как и следовало ожидать из предыдущих исследований, колонизация Candida была обнаружена у 28% женщин, причем Candida albicans преобладала более чем у 2/3. Однако у женщин с клиническими симптомами в области вульвы, например всегда следует помнить о дистрофии вульвы или неустановленном шелушении, о возможности колонизации Malassezia, ведущей к разноцветному лишайнику или фолликулиту Malassezia.
Идентификация компонентов аллергена условно-патогенных дрожжей Pityrosporum orbiculare с помощью моноклональных антител
Ссылки
1 января 1976 г. · Клиническая аллергия · JL LongbottomJ Pepys
1 апреля 1977 г. · Canadian Journal of Microbiology, MA Gord Salkin
1 апреля 1992 г. · Клиническая и экспериментальная аллергия: журнал Британского общества аллергии и клинической иммунологии · Дж. Саволайнен, А. Броберг
1 мая 1990 г. · Журнал Американской академии дерматологии · М. Киффер Свейгаард
1 января , 1992 · Журнал аллергии и клинической иммунологии · E Jensen-JarolimP Stahl Skov
1 сентября 1991 · Клиническая и экспериментальная дерматология · IM BergbrantT Söderström
1 сентября 1991 · Британский журнал дерматологии · MW WesselsE Young
1 января 1990 г. · Журнал медицинской и ветеринарной микологии: двухмесячная публикация Международного общества микологии человека и животных · С. Йоханссон, Дж. Фэргеман n1 марта 1990 · Acta paediatrica Scandinavica · SL Nordvall, S Johansson
1 мая 1990 · Архив дерматологии · M RokugoY Tomita
1 января 1988 · Международный архив аллергии и прикладной иммунологии · E JohanssonS G Johansson
1 марта 1986 · Журнал Американской академии дерматологии · J FaergemannA Scheynius
1 января 1985 · Журнал Американской академии дерматологии · O BäckR Hörnqvist
1 июля 1969 · Архив дерматологии · L JuhlinN Thyresson
1 июня 1968 · Журнал аллергии · J PepysF E Hargreave
1 марта 1973 · Архив дерматологии · BS PotterW C Johnson
1 апреля 1966 · Журнал аллергии · IH Itkin, M Dennis
1 апреля 1969 г. · Британский журнал дерматологии · С.О. Робертс
1 августа 1984 г. · Британский журнал дерматологии · С. Шустер
1 января 1983 г. · Британский журнал дерматологии · PG SohnleK E Huhta
1 января, 1980 · Международный архив аллергии и прикладная иммунология · GJ DaMertP G Sohnle
1 января 1951 г. · Annals of Internal Medicine · EL KEENEY
1 ноября 1951 г. · Журнал исследовательской дерматологии · MA GORDON
Цитирования
22 февраля 2001 · Бюллетень экспериментальной биологии и медицины · В.Г. Арзуманян Н. Н. Козлова
1 августа 1995 г. · Иммунология сегодня · GC MuddeC A. Bruijnzeel-Koomen
18 мая 1999 г. · Журнал аллергии и клинической иммунологии · A ZargariP Whitley 25 0007
июня 1999 · Аллергия · J Faergemann
9 апреля 1999 г. · Аллергия · MK KägiB Wüthrich
15 мая 1998 г. · Британский журнал дерматологии · A WolkerstorferA P Oranje
13 марта 1998 г. · Скандинавский журнал иммунологии · M Tengvall LinderA Scheynius
2 апреля 1999 г. · Скандинавский журнал иммунологии · C JohanssonA Scheynius
21 июня 2002 г. · Ветеринарная дерматология · Tai-An ChenPeter B Hill
2 апреля 1999 г. · Европейский журнал биохимии · Y OnishiK Takesako
6 июля 2000 г. · Европейский журнал биохимии · O RasoolA Scheynius
28 февраля 2002 г. · Acta Dermato-venereologica · A ZargariA Scheynius
1 ноября 1996 г. · Клиническая и экспериментальная аллергия: журнал Британского общества аллергии и аллергии. Клиническая иммунология · M Tengvall LinderA Scheynius
1 марта 1994 г. · Аллергия · G Doekes
1 августа 1997 г. · Аллергия · R van Ree
5 августа 1998 г. · Аллергия · B BarlettaG Di Felice
15 мая 1997 г. · Европейский журнал биохимии · М. Шмидта Шейниус
11 января 2005 г. · Прикладная и экологическая микробиология · Эдвард ДолкТео Веррипс
10 января 2002 г. · Обзоры клинической микробиологии · Х. Рут Эшби, Э. Глин В. Эванс
5 октября 2002 г. · Клинические Обзоры микробиологии · Ян Фэргеманн
25 декабря 2008 г. · PloS One · Анника М. Сяаф Мария Брэдли
21 декабря 2000 г. · Клиническая и экспериментальная аллергия: Журнал Британского общества аллергии и клинической иммунологии · E BuentkeA Scheynius
22 декабря 1999 г. · Клиническая и экспериментальная аллергия: журнал Британского общества аллергии и клинической иммунологии · М. Тенгвалл Линдер К. Уолгрен
23 августа 2002 г. · Клиническая и экспериментальная аллергия: журнал Британского общества аллергии и клинической иммунологии · N KandaS Watanabe
1 декабря 1995 г. · Клиническая и экспериментальная аллергия: журнал Британского общества аллергии и клинической иммунологии · A ZargariA Scheynius
1 июля 2005 г. · Обзор клинической иммунологии · SGO JohanssonHywel Williams
1 сентября 1999 г. · Журнал следственной дерматологии · М. Линдборг П. Уитли
13 июня 2002 г. · Журнал следственной дерматологии · Катарина ЙоханссонАнника Шейниус
17 июня 2006 г. · Журнал следственной дерматологии · Барбра Фишер Касагранде Петер Шмид-Гренделмайер 9000 июн. · Письма неврологов · Аббас ХагпарастМохаммад Камалинеджад
8 мая 2007 г. · Журнал молекулярной биологии · Моника Вильхельмссон Б. Мар tin Hallberg
24 февраля 2005 г. · Ветеринарная дерматология · Тай-Ань Чен, Питер Б. Хилл
14 мая 2004 г. · Скандинавский журнал иммунологии · E BuentkeA Scheynius
26 марта 2003 г. · Британский журнал дерматологии · C JohanssonM Tengvall Linder
24 апреля 2003 г. · Аллергия · A ZargariA Scheynius
20 июня 2007 г. · Микозы · AR KhosraviM Moazzeni
27 июля 2006 г. · Аллергия · C SelanderA Scheynius
23 февраля 2007 г. · Аллергия · A Scheynius
30 января 2014 г. · Журнал дерматологии · Carren S HauShinichi Watanabe
4 ноября 2004 г. · Журнал Американской академии дерматологии · Адитья К. Гупта Томас Л. Доусон
24 июля 1998 г. · Сообщения о биохимических и биофизических исследованиях · H Ясуедак Акияма
28 октября 2008 г. · Письма неврологов · Али ХатибиМохаммад Камалинеджад
21 января 2020 г. · Ветеринарная дерматология · Росс БондПитер Б. Хилл
21 марта 2009 г. · Журнал иммунологии: Официальный журнал Америки n Ассоциация иммунологов · Кристин Селандер · Каролина Лундериус Андерссон
22 декабря 2006 · Журнал иммунологии: Официальный журнал Американской ассоциации иммунологов · Андреас Лимахер Рето Крамери
1 апреля 1995 · Обзоры клинической микробиологии · WE HornerS B Lehrer
Роль L-DOPA в меланизации и производстве мицелия в Malassezia Furfur
Abstract
Меланины синтезируются организмами всех биологических царств и составляют гетерогенный класс природных пигментов.Некоторые из этих полимеров участвуют в патогенезе нескольких важных грибковых патогенов человека. В этом исследовании изучали, продуцирует ли грибковый кожный патоген Malassezia furfur меланин или меланин-подобные соединения. Меланин-связывающее моноклональное антитело (MAb), меченное in vitro , культивировало дрожжевые клетки M. furfur . Кроме того, меланизация дрожжей и гиф Malassezia была обнаружена антителами против меланина в соскобах от пациентов с разноцветным лишаем.Обработка дрожжей Malassezia протеолитическими ферментами, денатурирующим агентом и концентрированной горячей кислотой дала темные частицы, а спектроскопия электронного спинового резонанса показала, что эти частицы содержат стабильное соединение со свободными радикалами, что соответствует их идентификации как меланины. Дрожжи Malassezia требовали фенольных соединений, таких как L-DOPA, для синтеза меланина. L-ДОФА также запускал образование гиф in vitro в сочетании с койевой кислотой, ингибитором тирозиназы, в зависимости от дозы.В этом отношении L-ДОФА считается важным веществом, которое связано как с меланизацией, так и с дрожжевой мицелиальной трансформацией в M. furfur . Таким образом, M. furfur может продуцировать меланин или меланин-подобные соединения in vitro и in vivo , а путь меланина DOPA участвует в меланизации клеточной стенки.
Образец цитирования: Youngchim S, Nosanchuk JD, Pornsuwan S, Kajiwara S, Vanittanakom N (2013) Роль L-DOPA в меланизации и производстве мицелия в Malassezia Furfur .PLoS ONE 8 (6): e63764. https://doi.org/10.1371/journal.pone.0063764
Редактор: Флойд Л. Уормли, Техасский университет в Сан-Антонио, Соединенные Штаты Америки
Поступила: 27 декабря 2012 г .; Дата принятия: 5 апреля 2013 г .; Опубликовано: 7 июня 2013 г.
Авторские права: © 2013 Youngchim et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была поддержана Исследовательским фондом медицинского факультета Университета Чиангмая, Чиангмай, 50200, Таиланд. JDN частично поддерживается NIH AI52733. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Введение
Виды Malassezia представляют собой липофильные дрожжи, которые являются частым компонентом микрофлоры кожи человека и большинства теплокровных животных [1], [2].На основании молекулярных данных и потребностей в липидах в настоящее время распознаются семь видов Malassezia [3], [4]. У людей разноцветный лишай, известный также как разноцветный лишай, представляет собой обычную поверхностную инфекцию кожи, ограниченную роговым слоем, которая возникает, когда дрожжевых клеток Malassezia превращаются в патогенную мицелиальную форму. M. globosa и M. furfur являются значительно более частыми причинами PV, чем другие виды [5]. Заболевание характеризуется наличием мелких чешуйчатых пятен или пятен, которые могут быть гипо- или гиперпигментированными и обычно располагаются на верхних частях туловища, шеи и рук [5], [6].PV является одним из наиболее распространенных пигментных расстройств во всем мире, но чаще встречается в тропическом климате, достигая 50% в некоторых тропических регионах, что объясняется положительным эффектом для роста, достигаемым при относительно высокой температуре и влажности в этих регионах [ 5].
Поскольку ПВ является одной из наиболее распространенных кожных инфекций человека, изучение патофизиологических механизмов, лежащих в основе заболевания, может привести к более глубокому пониманию причин, развития и исходов инфекций человека Malassezia .Однако на сегодняшний день имеется относительно мало информации о факторах вирулентности Malassezia spp. Признаки дрожжей Malassezia , вовлеченные до сих пор в патогенез, включают липолитические ферменты, которые повреждают ткани хозяина и обеспечивают питательными веществами гриб [7] — [9], высокое содержание липидов в клеточных стенках, которое защищает дрожжи Malassezia от фагоцитоза [10] и подавляет воспалительный иммунный ответ [11], [12], образование гиф и выработку триптофан-зависимого пигмента, который действует как защитный барьер против UVA и UVB спектра [13], [14].
Меланины — это биологически важные макромолекулы, которые представляют собой темно-коричневые и черные пигменты, образованные в результате окислительной полимеризации фенольных соединений. Меланины можно разделить на 3 типичных типа: эумеланин; образуется в результате сложного процесса полимеризации с участием хинонов и свободных радикалов, феомеланин; производные тирозина, цистеина и алломеланинов; образованы из предшественников, не содержащих азота [15]. Обычно в грибах обнаруживаются два типа меланинов, 1,8-дигидроксинафталин (DHN) и L-3,4-дигидроксифенилаланин (DOPA), но большинство из них синтезируется из DHN-меланинов.Меланины, полученные из ацетата посредством поликетидсинтазного пути, обычно имеют черный или коричневый цвет и называются меланинами DHN. Меланины ДОФА катализируются фенолоксидазами (такими как тирозиназы, лакказы или катехолазы) и называются эумеланинами [16].
Они были связаны с вирулентностью ряда патогенных грибов человека, таких как Cryptococcus neoformans [17] — [19], Aspergillus fumigatus [20] — [24], Histoplasma capsulatum [25] и ). Penicillium marneffei [26].Было показано, что меланин защищает грибки несколькими способами. Он действует как полезный физиологический окислительно-восстановительный буфер, защищает патогены от стресса окружающей среды, такого как УФ-излучение, экстремальные температуры, гидролитические ферменты, металлы или свободные радикалы, и повышает устойчивость к противогрибковым препаратам [19].
Изучение меланизации у Malassezia spp. ранее был исследован с использованием красителя Массона-Фонтана, который продемонстрировал накопление черного пигмента на клеточной стенке дрожжевых клеток как in vitro, , так и во время инфекции [27].Однако окрашивание серебром по Массон-Фонтана не является специфичным для меланинов, о чем свидетельствует тот факт, что этим методом окрашиваются как меланизированные, так и непигментированные клетки C. neoformans [28]. В этом отчете мы подтвердили присутствие меланинов in vitro -меланизированных дрожжевых клеток M. furfur , используя методы, разработанные для изучения и выделения меланинов от других грибковых патогенов. Кроме того, для подтверждения меланизации как in vitro , так и во время инфицирования человека используется специфическое к меланину моноклональное антитело (MAb).Мы показываем, что M. furfur продуцирует ДОФА-меланин. Мы также показываем, что койевая кислота, ингибитор тирозиназы, может запускать трансформацию мицелия в M. furfur , что считается важным в патогенезе Malassezia .
Материалы и методы
Штамм грибов и среды
M. furfur NBRC 0656 поддерживали ежемесячным пересевом на модифицированной среде Диксона (mDixon; 1 литр дистиллированной воды, 36 г солодового экстракта, 6 г пептона, 20 г бычьей желчи (все получены от Difco), 10 мл Tween 40 , 2 мл глицерина, 2 мл оливкового масла (все получено от Sigma) 0.05 г хлорамфеникола (Amresco) и 15,0 г агара (Difco) с pH 6,0 (модифицировано из Guillot и соавторов [29]). Если не указано иное, M. furfur культивировали на химически определенном агаре с минимальной средой (MM) (15,0 мМ глюкозы, 10,0 мМ MgSO 4 , 29,4 мМ KH 2 PO 4 , 13,0 мМ глицина, 10,0 мМ NH 4 Cl и 3,0 мкМ тиамина; pH 5,5) с добавлением 0,1% (об. / Об.) Твин 40, 0,1% (об. / Об.) Твин 80 и 1,5% агара. Для исследований меланизации: M.furfur инкубировали при 30 ° C в течение 10–14 дней в MM с или без L-3,4-дигидроксифенилаланина (L-DOPA) (Sigma) в концентрации 1,0 мМ. Клетки дрожжей Malassezia собирали путем добавления 3,0 мл стерильного PBS в культуральные планшеты и удаляли осторожным соскабливанием с помощью петли. Клетки дрожжей собирали центрифугированием при 4500 g в течение 20 мин и осадки трижды промывали стерильным PBS.
Рост
M. furfur в среде с различными аминокислотами в качестве источников азотаДля изучения влияния источников азота, М.Дрожжи furfur культивировали на основной среде (3,0% Твин 80 (Sigma, Сент-Луис, Миссури, США), 50,0 мкг / мл хлорамфеникола (Амреско, Огайо, США) и 2,0% агаре (Дифко) (модифицировано Mayser et al.). [30]) с или без 15 мМ стерильного отфильтрованного L-триптофана (Tryp; Sigma) или аргинина (Arg; Sigma) в качестве источника азота аминогруппы. Для дальнейшего изучения влияния источников азота, трипа или Arg, в концентрации 0,6 % добавляли в среду mDixon вместо пептона в качестве источника азота.В качестве отрицательного контроля использовали mDixon без пептона или аминокислоты.Чашки инкубировали при 30 ° C в течение 10–14 дней.
Выделение и очистка частиц меланина из
M. furfurДрожжи Malassezia были подвергнуты протоколу экстракции меланина [31]. Вкратце, дрожжевые клетки промывали сначала PBS, а затем 1,0 М сорбитолом и 0,1 М цитратом натрия (pH 5,5). Добавляли лизирующий фермент из Trichoderma harzianum (Sigma) в концентрации 10 мг / мл и инкубировали в течение ночи при 30 ° C для образования протопластов.Протопласты собирали центрифугированием, трижды промывали PBS и инкубировали в 4,0 М тиоцианате гуанидина (Sigma) в течение ночи при комнатной температуре. Полученные темные частицы собирали центрифугированием, трижды промывали PBS и обрабатывали 1,0 мг / мл протеиназы K (Roche) в реакционном буфере [10,0 мМ Трис, 1,0 мМ CaCl 2 и 0,5% (мас. / Об.) SDS. , pH 7,8] и инкубировали при 37 ° C. Полученный мусор трижды промывали PBS и кипятили в 6,0 М HCl в течение 1,5 часов.После обработки кипячением в кислоте частицы меланина собирали фильтрованием через ватман № 1 и были тщательно промыты дистиллированной водой. Затем частицы меланина диализовали против дистиллированной воды в течение 10 дней до полного удаления кислоты, после чего их лиофилизировали.
Анализ спектроскопии электронного спинового резонанса (ЭПР)
ЭПР-спектроскопия — это метод, который непосредственно определяет сигнал от свободного спина электрона или свободных радикалов. Из-за обилия стабильных свободных радикалов в пигментах меланина, СОЭ используется в качестве инструмента для определения присутствия меланина в образцах с помощью диода Ганна в качестве источника микроволн [32].Сигнал ЭПР меланина в производном режиме показывает типичный одиночный пик, расположенный в середине спектра приблизительно при 3500 Гаусс. Мы использовали этот метод с общим количеством 2 г лиофилизированного материала в каждом случае (анализ проводился в кюветах с диоксидом кремния). В качестве положительного контроля использовали Aspergillus fumigatus и P. marneffei меланинов [24], [26].
Иммунофлуоресцентный анализ меланина в
M. furfur in vitroСуспензии дрожжевых клеток из М.furfur , выращенный на mDixon или MM с L-DOPA, сушили на воздухе на предметных стеклах с поли-L-лизином, а затем инкубировали с блокирующим буфером Superblock в PBS (Pierce) в течение 2 часов при 37 ° C или в течение ночи при 4 ° C для блокирования. неспецифическое связывание. Срезы трижды промывали PBS, затем инкубировали в течение 1,5 ч при 37 ° C с 10 мг / мл антимеланинового MAb 8D6 (против меланина A. fumigatus ; [24]), приготовленного в блокирующем буфере Superblock в PBS. После трехкратной промывки PBS слайды инкубировали с разведениями 1 × 100 козьих антимышиных IgM, конъюгированных с флуоресцеин-изотиоцианатом (Jackson Immunoresearch Laboratories) на 1.5 часов при 37 ° C. Предметные стекла трижды промывали PBS для удаления несвязавшегося антитела, а затем помещали в глицерин-PBS (1-1 об. / Об.). Накладывали покровные стекла и исследовали предметные стекла с помощью флуоресцентного микроскопа Nikon Eclipse 50i. В качестве отрицательного контроля в эксперименты включали только конъюгированный козий-антимышиный IgM (Jackson) без первичных MAb. Кроме того, Saccharomyces cerevisiae MMCM 5211 использовали в качестве отрицательного контроля. Изображения были сняты на Nikon DS Fi1.
Планшетный анализ фенолоксидазы
Полуколичественный анализ лакказы, оценивающий окисление L-DOPA в M.furfur был модифицирован из Srinivasan et al. [33] и Кроу и Олссон [34]. Для анализа на пластинке с лакказой 15 мл плавящегося стерильного MM-агара с 1,0 мМ L-DOPA помещали в стерильную чашку Петри (90 на 15 мм), содержащую три стерильных чашки (диаметром 6 мм). Чашки удаляли после затвердевания агара. Двести мкл суспензии дрожжевых клеток (доведенной до McFarland № 5) в стерильном PBS инокулировали в каждую лунку, и планшеты инкубировали при 28 ° C в течение 7-10 дней. C. neoformans H99 и S.cerevisiae MMCM 5211 использовали в качестве положительного и отрицательного контролей соответственно. Все планшеты ежедневно проверяли на предмет образования пигмента. Появление темно-черного цвета вокруг лунок считалось положительной реакцией на активность лакказы.
Влияние койевой кислоты на меланизацию
M. furfurКойевая кислота, ингибитор синтеза 3,4-дигидроксифенилаланина [ДОФА] меланина, была использована для определения типа меланина в M. furfur . M. furfur культивировали с койевой кислотой в различных концентрациях, 0, 600, 800, 1000, 1200 и 1500 мкг / мл, в определенной ММ с или без 1,0 мМ L-ДОФА с добавлением 0,1% Твина 40, 0,1% Твина. 80 при 30 ° C в течение 12–14 дней в темноте и исследованы на предмет образования пигмента. После инкубации дрожжи Malassezia окрашивали меланин-связывающим MAb 8D6, затем собирали и подвергали экстракции меланином, как описано выше в 2.3, для дальнейшего анализа извлеченных остатков с помощью ESR.
Индукция мицелиальной фазы
M. furfur in vitro с использованием L-DOPA и койевой кислотыЧтобы исследовать оптимальные условия для индуцирования продукции мицелия, клетки Malassezia инкубировали в полутвердой (0,5% агар) культуральной среде ММ, состоящей из липидных добавок и 1,0 мМ L-DOPA с различными концентрациями койевой кислоты. Чтобы продемонстрировать потребность в L-DOPA, также был протестирован агар, содержащий дофамин или L-тирозин с койевой кислотой.Культуры инкубировали при 30 ° C в течение 5-7 дней в микроаэрофильных условиях, используя анаэробный пластиковый сосуд с системой создания микроаэробной атмосферы (Pack MicroAero, Mitsubishi Gas Chemical Co. Inc), а затем просматривали под световым микроскопом (увеличение × 1000). для определения процента производства гиф. Для каждого условия было подсчитано всего 1000 грибковых клеток, чтобы определить процент филаментации. Кроме того, M. furfur NBRC 10987, CBS 6000, 6001, 6046, 6094 и 7966 (изоляты пациентов, любезно полученные от Dr.Теун Бекхаут, Centraal bureau voor Schimmelcultures (CBS-KNAW), Yeast Research Group, Утрехт, Нидерланды) были исследованы на их способность образовывать филаменты в этих условиях. Кроме того, в это исследование были включены два изолята M. furfur от пациентов без кожных заболеваний, H 1 и H 2 .
Обнаружение меланизации разноцветного лишая на инфицированной коже
Образцы чешуек на коже пациентов с гипопигментированными или гиперпигментированными поражениями ЛВ были получены из дерматологической клиники больницы Махараджа Накорна в Чиангмае, Университет Чиангмая.Кожные чешуйки, использованные в экспериментах, были получены в ходе повседневного ухода за пациентами с дерматологическими заболеваниями. Письменное согласие было получено от всех пациентов, перенесших дерматологический соскоб, и согласие включало, что ткани могут быть использованы в экспериментальных целях, за исключением генетических исследований на людях, без дополнительного согласия. Процедура была одобрена этическим комитетом Университета Чиангмая. Было проведено прямое обследование кожи, и присутствие Malassezia было подтверждено путем культивирования на агаре mDixon, инкубируемом при 30 ° C в течение 2 недель.Дрожжи Malassezia были идентифицированы до видового уровня с помощью полимеразной цепной реакции (ПЦР), как описано [35]. Для исследования меланизации Malassezia in vivo кожу переваривали 1,0 мг / мл протеиназы К (Roche) при 37 ° C в течение 2 часов и трижды промывали PBS (модифицировано из Youngchim et al. [36]) ). Затем кожу зондировали с помощью меланин-связывающего MAb 8D6 и FITC-конъюгированного козьего антимышиного IgM, как описано выше. Были включены отрицательные контроли без первичных MAb.
Статистический анализ
Процент производства филаментов в M. furfur , культивированных в ММ с L-ДОФА и койевой кислотой в различных концентрациях, сравнивали с контролем, M. furfur в ММ или ММ с L-ДОФА с помощью двуххвостого непарного t Стьюдента. -тест с помощью программы Prism 4 (GraphPad). Значение P ≤0,05 считалось значимым.
Результаты
Меланизация
дрожжевых клеток M. furfur in vitroМ.furfur NRBC 0656 культивировали на MM с или без L-DOPA в течение 7–10 дней при 30 ° C. Все культуры ежедневно наблюдали на предмет продукции пигмента. Рост Malassezia потребовал включения источника липидов в культуральную среду. Через 7 дней колонии Malassezia , выращенные на ММ с добавлением L-DOPA, показали темно-коричневый пигмент (рис. 1A), а интенсивность темного цвета увеличивалась при продолжении инкубации. Обработка этих пигментированных клеток протоколом экстракции меланина привела к сбору агрегированных темных частиц.Напротив, дрожжевых клеток M. furfur , выращенных в минимальной среде без L-DOPA, были макроскопически непигментированными (рис. 1В), а непигментированные клетки были полностью солюбилизированы, подвергнув их протоколу экстракции меланина.
ЭПР-спектроскопия
M. furfur Меланин СпектроскопияЭПР меланиноподобных частиц, собранных из M. furfur NBRC 0656, выращенных на минимальной среде с L-DOPA, дала сигналы, указывающие на присутствие стабильной популяции свободных радикалов, соответствующей пигменту, идентифицированному как меланины (рис. 2A. ) [32].Спектр был неотличим от сигналов, производимых меланинами других патогенных грибов, таких как C. neoformans [31], H. capsulatum [25], A. fumigatus [24] и P. marneffei [26]. ]. Черные частицы выделены из М . furfur , выращенный на среде mDixon (рис. 2B) и базальных средах с L-трипом или Arg (рис. 2C и D, соответственно), также давал сигналы ESR, соответствующие меланину. Напротив, M. furfur , культивированный на MM без L-DOPA, полностью растворялся при выполнении протокола экстракции меланина.
Рисунок 2. ЭПР-спектроскопия частиц меланина, экстрагированных из M. furfur NBRC 0656, выращенных на определенной минимальной среде с L-DOPA (A), среде mDixon (B), базовой среде с L-трипом (C) и базовой среде с Arg (D) при 30 ° C в течение 7–10 дней.
https://doi.org/10.1371/journal.pone.0063764.g002
Иммунофлуоресценция (IF) Реакционная способность антител к меланину MAb 8D6 —
M. furfur Дрожжевые клетки до и после экстракции меланинаМ.Клетки дрожжей furfur , выращенные как в среде mDixon, так и в MM с L-DOPA, реагировали с меланин-специфическим MAb 8D6, демонстрируя выраженную флуоресцентную реактивность внутри клеточной стенки (рис. 3A, B). Напротив, M. furfur , выращенный на MM без L-DOPA, не был помечен MAb 8D6. После процедуры экстракции меланина на дрожжевых клетках, выращенных в среде mDixon, полученные частицы в обоих изолятах дрожжевых клеток Malassezia сохранили реактивность к MAb 8D6 (рис. 3C, D). При использовании одного вторичного MAb не было реактивности с дрожжевыми клетками Malassezia или частицами меланина (данные не показаны). S. cerevisiae также был отрицательным в отношении антител к меланину MAb 8D6.
Рисунок 3. Соответствующие иммунофлуоресцентные и светлопольные микроскопические изображения, демонстрирующие мечение дрожжевых клеток M. furfur NBRC 0656 (A, B) и после (C, D) экстракции меланина меланин-связывающим MAb 8D6. Полосы представляют собой 5 мкм.
https://doi.org/10.1371/journal.pone.0063764.g003
Планшетный анализ фенолоксидазы
Планшетный анализ фенолоксидазы основан на способности организма окислять L-ДОФА, включенный в среду, что приводит к образованию черного цвета вокруг суспензий клеток Malassezia .Результаты анализа на планшете с фенолоксидазой (рис. 4) продемонстрировали наличие лакказной активности в суспензиях клеток M. furfur NBRC 0656 и C. neoformans H99 (положительный контроль), но реактивность с S. cerevisiae не наблюдалась. (отрицательный контроль).
Рисунок 4. Анализ на планшете для демонстрации активности фенолоксидазы в течение 10 дней. M. furfur NBRC 0656 высевали в лунки на планшете с лакказным агаром, содержащим 1 мМ L-DOPA в качестве субстрата.
C. neoformans H99 и S. cerevisiae MMCM 5211 использовали в качестве положительного и отрицательного контролей соответственно. Развитие интенсивного черного цвета вокруг лунки указывает на положительный результат теста на фенолоксидазную активность.
https://doi.org/10.1371/journal.pone.0063764.g004
Влияние койевой кислоты на меланизацию
M. furfur NRBC 0656Койевая кислота блокировала меланизацию L-ДОФА в M. furfur (Рисунок S1).В то время как дрожжи на MM с 1,0 мМ L-DOPA приобрели темно-коричневый цвет (рисунок S1B), всего лишь 100 мкг / мл койевой кислоты уменьшили пигментацию M. furfur (рисунок S1C). При 600 мкг / мл койевой кислоты колонии выглядели светло-желтыми (фигура S1F), а при 1000 мкг / мл (фигура S1H) колонии и чашки были эквивалентны по внешнему виду дрожжам, культивируемым на MM без L-DOPA. Следовательно, койевая кислота значительно снижает меланизацию в M . furfur поддерживает участие пути ДОФА в синтезе меланина.Однако, хотя койевая кислота была способна блокировать явно наблюдаемую меланизацию у M. furfur, эти организмы все еще были реактивными к меланин-связывающим MAb (рис. 5). Кроме того, черные частицы получали, когда мицелиальные клетки M. furfur , культивированные в 1 мМ L-DOPA и 1500 мкг / мл койевой кислоты, обрабатывали протоколом экстракции меланина (данные не показаны).
Рис. 5. Соответствующие иммунофлуоресцентные и светлопольные микроскопические изображения, демонстрирующие мечение дрожжевых клеток M.furfur NBRC 0656 (A, B) и CBS 7019 (C, D), выращенные на MM с 1 мМ L-DOPA и 1500 мкг / мл койевой кислоты с помощью меланин-связывающего MAb 8D6. Полосы представляют собой 5 мкм.
https://doi.org/10.1371/journal.pone.0063764.g005
Индукция мицелиальной фазы
M. furfur с L-ДОФА и койевой кислотойВ дополнение к ингибирующему эффекту койевой кислоты на меланизацию Malassezia , мы наблюдали, что добавление койевой кислоты и L-ДОФА в ММ индуцировало превращение дрожжевых клеток в формы коротких нитей (мицелиальная фаза) (рис. 6).Хотя только L-ДОФА индуцировал трансформацию гиф, процент гиф увеличивался дозозависимым образом с добавлением койевой кислоты (рис. 6B-G и 7). Примечательно, что койевая кислота не способна вызывать филаментацию у дрожжей Malassezia в отсутствие L-ДОФА (фигура 6H). Трансформации коротких филаментов наблюдались и у других изолятов M . furfur , включая CBS 6000, 6001, 7019 и два дополнительных штамма от здорового человека, инкубированного с L-DOPA и 1500 мкг / мл койевой кислоты (фиг. 8).Напротив, ни дофамин, ни L-тирозин не были эффективны в стимуляции продукции мицелия в M. furfur при совместном культивировании с койевой кислотой (данные не показаны).
Рисунок 6. Влияние L-DOPA и койевой кислоты на морфологию M. furfur NBRC 0656 при культивировании в MM (A), MM с 1 мМ L-DOPA (B), MM с 1 мM L-DOPA и койевая кислота при 600 (C), 800 (D), 1000 (E), 1200 (F), 1500 (G) мкг / мл и MM с 1000 мкг / мл койевой кислоты (H). Полосы представляют собой 5 мкм.
https: // doi.org / 10.1371 / journal.pone.0063764.g006
Рис. 7. Влияние различных концентраций койевой кислоты на образование волокон в M. furfur NBRC 0656 в микроаэробных условиях.
Данные представляют собой средние значения ± стандартное отклонение среднего (стандартное отклонение средних значений) на основе не менее трех экспериментов. Статистически значимая разница (двусторонний непарный t-критерий Стьюдента) по сравнению с контролем, MM и MM с L-DOPA, * p≤0,05.
https://doi.org/10.1371/journal.pone.0063764.g007
Рисунок 8.Продукция мицелия различных изолятов в M. furfur .
M. furfur CBS 6046 (A), CBS 6000 (B), CBS 6001 (C), CBS 7019 (D) и два изолята от здорового человека (E, F) культивировали на MM с 1 мМ L- ДОФА и койевая кислота в 0,5% -ном агаре в микроаэрофильной среде при 30 ° C в течение 5–7 дней. Концентрация койевой кислоты составляла 1500 мкг / мл для стандартных изолятов (A-D) и 500 мкг / мл для здоровых изолятов (E, F). Полосы представляют собой 5 мкм.
https: // doi.org / 10.1371 / journal.pone.0063764.g008
Обнаружение меланизации в
Malassezia Зараженной кожеСоскобы кожи трех пациентов с PV были окрашены препаратом KOH для подтверждения присутствия почкующихся дрожжевых клеток и коротких нитей в образцах. M. globosa был идентифицирован в двух образцах ЛВ с гипер- и гипопигментированными поражениями, а M. furfur был выделен из гипопигментированного поражения. Круглые почкующиеся дрожжевые клетки и короткие филаменты в образцах чешуек кожи из гиперпигментированных или гипопигментированных поражений ЛВ были сильно реактивными с меланин-связывающим MAb 8D6 (фиг.9).Никакой реактивности не наблюдали, когда конъюгированный с флуоресцеинизотиоцианатом (FITC) козий антимышиный IgM инкубировали без первичного MAb.
Рис. 9. Соответствующие изображения иммунофлуоресценции (A, C, E, G) и светлопольной микроскопии (B, D, F, H), демонстрирующие мечение бластоконидий и коротких нитей со соскобов кожи разноцветного лишая с помощью меланин-связывающих MAb. 8D6.
Полоски составляют 5 мкм.
https://doi.org/10.1371/journal.pone.0063764.g009
Обсуждение
Меланины были признаны важной детерминантой вирулентности нескольких важных грибковых патогенов человека, поскольку имеются обширные доказательства того, что пигмент поддерживает выживание в организме хозяина и способствует способности организмов вызывать заболевания [37]. В этом исследовании мы экспериментально подтвердили, что меланин синтезируется дрожжами Malassezia , что соответствует данным Gaitanis et al. [27]. Наши доказательства, подтверждающие меланизацию дрожжей Malassezia , следующие: (i) анализ спектроскопии ESR черных частиц показал присутствие стабильного свободнорадикального соединения, совместимого с меланином, (ii) реактивность меланин-связывающего MAb на клеточной стенке. дрожжей Malassezia , выращенных in vitro и с пигментированными частицами, полученными из этих клеток; соскоб с инфицированной кожи Malassezia .Результаты СОЭ представляют собой первое биофизическое свидетельство продукции меланина в M. furfur , а анализ ферментативных планшетов — первое химическое свидетельство фенолоксидазы в Malassezia . Кроме того, для обнаружения меланизации у M. furfur как in vitro , так и in vivo применяли меланин-связывающее MAb 8D6, индуцированное против меланина DHN из A. fumigatus . Ранее было показано, что MAb 8D6 реагирует с меланинами DHN и DOPA из других грибов [24].Очевидно, что меланин DHN должен иметь общие эпитопы с меланином DOPA, что также было показано в других исследованиях [18], [38], [39]. Вместе эти наблюдения предоставляют твердые доказательства того, что Malassezia может синтезировать меланин или меланин-подобное соединение in vitro и in vivo . В нашем исследовании дрожжи Malassezia и гифы как в гипер-, так и в гипопигментированных поражениях кожи ЛВ были сильно реактивны с меланин-специфическими МАт, что указывает на то, что меланизация происходит in vivo в обеих формах ЛВ.
Интересно, что исследование Mayser и Pape [13] продемонстрировало, что синтез пигментов и флуорохромов M. furfur обеспечивает мощную защиту от излучения диапазонов UVA и UVB по сравнению с выживаемостью непигментированных клеток. Документация по производству меланина M. furfur подтверждает, что полимер участвует в защите от УФ-излучения, учитывая данные по радиозащиты меланинов грибов [37], [40]. Интересно, что радиозащитная эффективность меланинов грибов является функцией химического состава и пространственного расположения полимеров [41].
Виды Malassezia — это диморфные грибы, способные к дрожжево-мицелиальным изменениям. При заражении Malassezia круглые или овальные дрожжевые клетки переходят в формы мицелия [42]. Таким образом, считается, что образование гиф связано с вирулентностью. Чтобы исследовать патогенность и антимикотическую чувствительность Malassezia , исследователи пытались вызвать рост мицелиальной формы in vitro , но предыдущие попытки создать среду для трансформации гиф оказались безуспешными.В экспериментах с использованием рогового слоя человека исследователи смогли получить только 24% гиф в Pityrosporum ovale ( M. furfur ) [43]. Было высказано предположение, что одно или несколько веществ в роговом слое необходимы для активации образования гиф. В наших экспериментах M. furfur полностью трансформировалось в гифы при добавлении L-DOPA и койевой кислоты в среду MM с липидами. Это открытие поднимает интригующую возможность того, что дрожжи Malassezia способны продуцировать ингибитор тирозиназы для приобретения L-DOPA в среде хозяина.Это предположение подтверждается предыдущим исследованием Наззаро-Порро и Пасси [44], которые обнаружили ингибиторы тирозиназы в культурах Pityrosporum orbiculare ( M. furfur ) с добавлением олеиновой кислоты или вакценовой кислоты. Ингибиторы тирозиназы, секретируемые Malassezia spp, вероятно, подавляют образование меланина, приводящее к депигментации кожи человека, но не влияя на меланизацию Malassezia . Следовательно, этот ингибитор может иметь ингибирующий эффект, отличный от койевой кислоты, которая ингибирует тирозиназу, а также предотвращает превращение o -хинонов L-ДОФА [45].Из-за базовой культуральной среды в наших исследованиях MM (голодная среда) с липидами требуется для индукции мицелия Malassezia in vitro, , что аналогично исследованию Дорна и Ренерта [46].
Помимо ингибирующего действия тирозиназы, койевая кислота, по-видимому, влияет на морфологию дрожжей Malassezia . Койевая кислота обладает множеством активностей в отношении различных грибов, включая противогрибковую активность против Candida albicans , C. neoformans и Trichophyton rubrum [47], а также может влиять на выработку афлатоксина в A.flavus [48]. Похоже, что этот ингибитор может вызвать изменения в дрожжах Malassezia , которые происходят на здоровой коже, заставляя их развивать филаменты и превращаться в паразитическую форму.
В нашем исследовании фенолоксидаза была обнаружена в M. furfur , что предполагает образование меланина ДОФА в этом организме, аналогично исследованию Gaitanis et al. [27]. В дополнение к нашему открытию, что L-DOPA может влиять на синтез мицелия, L-DOPA является важным субстратом для производства меланина в Malassezia spp.Следовательно, преобразование Malassezia в vivo зависит от присутствия L-DOPA в эпидермисе. Было проведено два исследования, подтверждающих наличие L-ДОФА в эпидермисе. Во-первых, L-ДОФА был обнаружен в коже пигментированных животных с помощью газовой хроматографии [49]. Во-вторых, клетки Лангерганса обладают транспортным механизмом L-DOPA [50], что позволяет предположить, что это соединение присутствует и участвует в биологических ответах в местных тканях.
Было показано, что производство волокон в Pityrosporum ( Malaassezia ) более заметно в микроаэрофильной среде [51].Соответственно, наши результаты согласуются с предыдущими исследованиями, которые могут объяснить выживание Malassezia spp в самых глубоких частях волосяных фолликулов [52] или роговом слое [53], [54], растущих как в дрожжах. и мицелиальные формы [52].
В заключение, наши результаты показали, что M. furfur синтезирует меланин или меланин-подобный пигмент при выращивании in vitro и in vivo . Считается, что L-ДОФА и койевая кислота играют роль в запуске синтеза гиф в M.furfur при инкубации в микроаэрофильной среде. Дальнейшие исследования следует изучить на других видах дрожжей Malassezia , выделенных из здоровых и ЛВ кожных поражений как в гипер-, так и в гипопигментированных поражениях, в частности, для идентификации видов и их способности образовывать гифы in vitro . Кроме того, недавно были разработаны стратегии генетических манипуляций для Malassezia spp, и они будут использоваться в проспективных исследованиях, направленных на изучение генов, ответственных за образование меланина, и ингибиторов тирозиназы.
Благодарности
Авторы хотели бы поблагодарить доктора Сири Чевчанвит, Фахарин Пундонтонг за предоставленные образцы кожи разноцветного лишая и доктора Теуна Бекхаута за предоставление стандартных изолятов Malassezia furfur .
Вклад авторов
Задумал и спроектировал эксперименты: SY JDN. Проведены эксперименты: SY. Проанализированы данные: SY JDN SP. Внесенные реактивы / материалы / инструменты анализа: SK NV. Написал бумагу: SY JDN.
Список литературы
- 1. Leeming JP, Notman FH, Holland KT (1989) Распространение и экология Malassezia furfur и кожных бактерий на коже человека. J Appl Bacteriol 67: 47–52.
- 2. Таразоо Б., Кордбаче П., Зайни Ф., Зомородян К., Саадат Ф. и др. (2004) Изучение распространения видов Malassezia у пациентов с разноцветным лишаем и здоровых людей в Тегеране, Иран. BMC Dermatol 1: 4–5.
- 3.Guého E, Midgley G, Guillot J (1996) Род Malassezia с описанием четырех новых видов. Антони Ван Левенгук 69: 337–355.
- 4. Гупта А.К., Кохли Ю., Саммербелл Р.К. (2000) Молекулярная дифференциация семи видов Malassezia . J Clin Microbiol 38: 1869–1875.
- 5. Crespo-Erchiga V, Florencio VD (2006) Malassezia дрожжи и разноцветный лишай. Curr Opin Infect Dis 19: 139–147.
- 6.Gupta AK, Ryder JE, Nicol K, Cooper EA (2003) Поверхностные грибковые инфекции: обновленная информация о разноцветном лишайнике, себорейном дерматите, микозе головы и онихомикозе. Clin Dermatol 21: 417–425.
- 7. Brunke S, Hube B (2006) MfLIP1, ген, кодирующий внеклеточную липазу липид-зависимого гриба Malassezia furfur . Microbiol 152: 547–554.
- 8. Шибате Н., Оканума Н., Хираи К., Арикава К., Кимура М. и др. (2006) Выделение, характеристика и молекулярное клонирование липолитического фермента, секретируемого из Malassezia pachydermatitis .FEMS Microbiol Lett 256: 137–144.
- 9. ДеАнгелис Ю.М., Сондерс К.В., Джонстон К.Р., Ридер Н.Л., Коулман К.Г. и др. (2007) Выделение и экспрессия гена липазы Malassezia globosa , LIP1. J Invest Dermatol 127: 2138–2146.
- 10. Kesavan S, Holland KT, Ingham E (2000) Влияние экстракции липидов на иммуномодулирующую активность Malassezia видов in vitro . Med Mycol 38: 239–247.
- 11. Kesavan S, Walters CE, Holland KT, Ingham E (1998) Эффекты Malassezia на продукцию провоспалительных цитокинов мононуклеарными клетками периферической крови человека in vitro .Med Mycol 36: 97–106.
- 12. Thomas DS, Ingham E, Bojar RA, Holland KT (2008) In vitro модуляция про- и противовоспалительных цитокинов кератиноцитов человека капсулой видов Malassezia . FEMS Immunol Med Microbiol 54: 203–114.
- 13. Mayser P, Pape B (1998) Снижение восприимчивости Malassezia furfur к УФ-свету за счет синтеза производных триптофана. Антони Ван Левенгук 73: 315–319.
- 14.Mayser P, Wille G, Imkampe A, Thoma W, Arnold N, et al. (1998) Синтез флуорохромов и пигментов в Malassezia furfur с использованием триптофана в качестве единственного источника азота. Микозы 41: 265–271.
- 15. Гамильтон А.Дж., Гомес Б.Л. (2002) Меланины в грибковых патогенах. J Med Microbiol 51: 189–191.
- 16. Wheeler MH, Bell AA (1988) Меланины и их значение в патогенных грибах. Curr Top Med Mycol 2: 338–387.
- 17. Носанчук JD, Rosas AL, Lee SC, Casadevall A (2000) Меланизация Cryptococcus neoformans в ткани мозга человека.Ланцет 10: 2049–2050.
- 18. Росас А.Л., Носанчук Дж.Д., Фельдмессер М., Кокс Г.М., МакДейд Х.С. и др. (2000) Синтез полимеризованного меланина Cryptococcus neoformans у инфицированных грызунов. Заражение иммунной 68: 2845–2853.
- 19. Casadevall A, Rosas AL, Nosanchuk JD (2000) Меланин и вирулентность в Cryptococcus neoformans . Curr Opin Microbiol 3: 354–358.
- 20. Ян Б., Кох А., Шмидт А., Лангфельдер К., Ваннер Г. и др.(1997) Выделение и характеристика бесцветного конидиального мутанта Aspergillus fumigatus с измененной конидиальной поверхностью и пониженной вирулентностью. Инфекция иммунной 65: 5110–5117.
- 21. Лангфельдер К., Ян Б., Герингер Х., Шмидт А., Ваннер Г. и др. (1998) Идентификация гена поликетидсинтазы (pksP) Aspergillus fumigatus , участвующего в биосинтезе конидиального пигмента и вирулентности. Med Microbiol Immunol 187: 79–89.
- 22. Tsai HF, Chang CY, Washburn GR, Wheeler MH, Kwon-Chung KJ (1998) Регулируемый онтогенетическим геном alb1 ген Aspergillus fumigatus : его роль в модуляции морфологии и вирулентности конидий.J Bacteriol 180: 3031–3038.
- 23. Tsai HF, Wheeler MH, Chang YC, Wheeler MH, Kwon-Chung KJ (1999) Регулируемый в процессе развития кластер генов, участвующих в биосинтезе конидиального пигмента у Aspergillus fumigatus . J Bacteriol 181: 6469–6477.
- 24. Youngchim S, Morris-Jones R, Hay RJ, Hamilton AJ (2004) Производство меланина Aspergillus fumigatus . J Med Microbiol 53: 175–181.
- 25. Носанчук Дж. Д., Гомес Б. Л., Янгчим С., Диес С., Айсен П. и др.(2002) Histoplasma capsulatum синтезировал меланин-подобные пигменты in vitro и во время инфицирования млекопитающих. Заражение иммунной 70: 5124–5131.
- 26. Youngchim S, Hay RJ, Hamilton AJ (2005) Меланизация Penicillium marneffei in vitro, и in vivo. . Microbiol 151: 291–299.
- 27. Gaitanis G, Chasapi V, Velegraki A (2005) Новое применение красителя Masson-Fontana для демонстрации производства меланин-подобного пигмента Malassezia видов in vitro и в клинических образцах.J Clin Microbiol 43: 4147–4151.
- 28. Kwon-Chung KJ, Hill WB, Bennett JE (1981) Новый специальный краситель для гистопатологической диагностики криптококкоза. J Clin Microbiol 13: 383–387.
- 29. Guillot J, Breugnot C, de Barros M, Chermette R (1998) Полезность модифицированной среды Диксона для количественного культивирования видов Malassezia из кожи собак. J Vet Diagn Invest 10: 384–386.
- 30. Mayser P, Töws A, Krämer HJ, Weiss R (2004) Дальнейшая характеристика производящих пигмент штаммов Malassezia .Микозы 47: 34–39.
- 31. Wang Y, Casadevall A (1996) Меланин, меланин «призрак» и состав меланина в Cryptococcus neoformans : идентификация как лакказа. J Bacteriol 176: 656–664.
- 32. Enochs WS, Nilges MJ, Swartz HM (1993) Стандартизованный тест для идентификации и характеристики меланина с использованием электронной парамагнитной (ЭПР) спектроскопии. Pigm Cell Res 6: 91–99.
- 33. Srinivasan C, Dsouza M, Boominathan K, Reddy CA (1995) Демонстрация лакказы в белой гнили Basidiomycete Phanerochaete chrysosporium BKM-F1767.Appl Environ Microbiol 61: 4274–4277.
- 34. Crowe JD, Olsson S (2001) Индукция активности лакказы в Rhizoctonia solani с помощью антагонистических штаммов Pseudomonas fluorecens и ряда химических обработок. Appl Environ Microbiol 67: 2088–2094.
- 35. Affes M, Salah SB, Makni F, Sellami H, Ayadi A (2009) Молекулярная идентификация видов Malassezia , выделенных от поражений дерматитом. Микозы 52: 251–256.
- 36.Youngchim S, Pornsuwan S, Nosanchuk JD, Dankai W, Vanittanakom N (2011) Меланогенез у дерматофитов in vitro и во время инфекции. Microbiol 157: 2348–2356.
- 37. Носанчук JD, Casadevall A (2003) Вклад меланина в микробный патогенез. Cell Microbiol 5: 203–223.
- 38. Носанчук Ю.Д., Росас А.Л., Касадевалл А (1998) Антительный ответ на грибковый меланин у мышей. J Immunol 160: 6026–6031.
- 39.Моррис-Джонс Р., Янгчим С., Гомес Б.Л., Айсен П., Хэй Р.Дж. и др. (2003) Синтез меланин-подобных пигментов Sporothrix schenckii in vitro и во время инфекции млекопитающих. Infect Immun 71: 4026–4033.
- 40. Носанчук JD, Casadevall A (2006) Влияние меланина на вирулентность микробов и клиническую устойчивость к антимикробным соединениям. Антимикробные агенты Chemother 50: 3519–3528.
- 41. Дадачева Э., Брайан Р.А., Хауэлл Р.К., Швейцер А.Д., Айзен П. и др.(2008) Радиозащитные свойства грибкового меланина зависят от его химического состава, стабильного присутствия радикалов и пространственного расположения. Pigment Cell Melanoma Res 21: 192–199.
- 42. Faergemann J, Fredriksson T (1981) Экспериментальные инфекции у кроликов и людей с Pityrosporum orbiculare и P. ovale . J Invest Dermatol 77: 314–318.
- 43. Faergemann J (1989) Новая модель роста и производства волокон Pityrosporum ovale ( orbicular ) на роговом слое человека in vitro .J Invest Dermatol 92: 117–119.
- 44. Наззаро-Порро М., Пасси С. (1978) Идентификация ингибиторов тирозиназы в культурах Pityrosporum . J Invest Deramatol 71: 205–208.
- 45. Кан В. (1995) Влияние койевой кислоты на окисление DL-ДОФА, норэпинефрина и дофамина тирозиназой грибов. Pig Cell Res 8: 234–240.
- 46. Дорн М., Рёнерт К. (1977) Диморфизм Pityrosporum orbiculare в определенной культуральной среде.J Invest dermatol 69: 244–248.
- 47. Chee HY, Lee EH (2003) Фунгистатическая активность койевой кислоты против патогенных грибов человека и ингибирование продукции меланина в Cryptococcus neoformans . Микобиология 31: 248–250.
- 48. Эль-Хадем М., Тевфик М.С., Хамди Ю.А. (1976) Стимулирующее действие койевой кислоты на производство афлатоксина изолятами Aspergillus flavus Link. Zentralbl Bakteriol Parasitenkd Infektionskr Hyg. 131: 497–500.
- 49. Эйзенхоф Дж., Тиан Х., Холмс С., Мацунага Дж., Роффлер-Тарлов С. и др. (2003) Тирозиназа: важный фактор развития периферического дофамина. Журнал FASEB 17: 1248–1255.
- 50. Falck B, Bendsoe N, Ronquist G (2003) Новый механизм притока аминокислот в эпидермальные клетки Лангерганса человека: система противотранспорта L-допа / протонов. Exp Dermatol 12: 602–609.
- 51. Faergemann J, Bernander S (1981) Микроаэрофильный и анаэробный рост Pityrosporum orbiculare.. Sabouraudia 19: 171–179.
- 52. Маринаро Р.Э., Гершенбаум М.Р., Ройзен Ф.Дж., Папа С.М. (1978) Разноцветный лишай: вид с помощью сканирующего электронного микроскопа. Дж. Кутан Патол 5: 15–22.
- 53. Montes LE (1970) Системные аномалии и внутриклеточные очаги инфекции рогового слоя. JAMA 213: 1469–1472.
- 54. Tosti A, Villardita S, Fazzini ML (1972) Паразитарная колонизация рогового слоя разноцветного лишая. J Invest Dermatol 59: 233–237.
|