Совместимость глицина и алкоголя: влияние на организм, противопоказания
В этой статье будет рассказано, как взаимодействуют глицин и алкоголь – совместимость приема аминокислоты и действие алкоголя на организм.
Свойства глицина
Аминокислоты это важнейшие компоненты всех метаболических активностей и жизненных процессов человека. Так же это одна из самых простых видов аминокислот, встречающихся в природе.
Итак, для чего он нужен:
- Это кристаллическое твердое вещество также используется в качестве подсластителя или усилителя вкуса. Некоторые пищевые добавки и различные белковые напитки содержат глицин.
- Некоторые препараты включают глицин, который помогает в улучшении абсорбции желудка.
- Это действует как буферный агент в анальгетиках, антацидах, антиперспирантах, косметике, туалетных принадлежностях и т. д.
- Он действует как промежуточное звено в процессе производства различных химических продуктов, а также используется при изготовлении гербицидного глифосата.


Но в данном контексте среди свойств глицина выделяются такие:
- Укрепляет иммунитет
- Способствует улучшению функций мозга.
- Укрепляет ЦНС, предотвращая поведенческие расстройства.
- Защищает печень от вредных последствий алкоголя
Глицин – это аминоуксусная кислота, которая содержится в определенных продуктах. Он называется «биотический». Это всепоглощающее вещество, которое успешно и давно используется в медицинской практике.
к содержанию ↑Алкоголь – влияние на организм
Употребление алкоголя приводит к:
| Область влияния | Подробности |
| Покраснения на лице. | Поскольку алкоголь увеличивает кровообращение, это приводит к тому, что ваше лицо становится красным в основном в щеках и области носа. |
| Ухудшение памяти. | Для любителей выпить, доказано, что алкоголь способен устроить в голове беспорядок, поэтому можно наблюдать не только кратковременную потерю памяти, но и забыть о том, что происходило на прошлой неделе. |
| Большая потребность во сне. | Еще один эффект алкоголя на организм заключается в том, что он вносит коррективы в привычный режим сна. И из-за этого, утро сопровождается не только похмельем но и усталостью, за счет сил, использованных для восстановления. |
| Нагрузка на сердце. | Алкоголь является сосудорасширяющим средством – он позволяет периферическим кровеносным сосудам расслабляться, позволяя большему количеству крови течь через кожу и ткани, что приводит к снижению артериального давления. Чтобы поддерживать достаточный приток крови к органам, частота сердечных сокращений возрастает. Скорость дыхания также может ускориться. |
| Нагрузка на печень. | Алкоголь метаболизируется в печени, а чрезмерное употребление алкоголя может привести к острой и хронической болезни печени. Когда печень разрушает спирт, образуются побочные продукты, такие как ацетальдегид, некоторые из которых более токсичны для организма, чем сам алкоголь. |
Некоторые люди, злоупотребляющие алкогольными напитками, могут развить физическую и эмоциональную зависимость от алкоголя. А его выведение из организма, после всего прочего может быть сложным и опасным для жизни. А потому часто нужна профессиональная помощь, чтобы избавится от алкогольной зависимости.
к содержанию ↑Совместимость алкоголя и глицина
Основным принципом работы препарата в том, что его компоненты совместимы и взаимодействуют с молекулами этанола, помогая организму производить максимум специальных ферментов, что бы нейтрализовать продукты распада алкоголя. Благодаря этому, препарат эффективно снижает похмельный синдром.
к содержанию ↑Помощь при похмельном синдроме
Этот биотик является хорошим средством для похмелья. Таблетки достаточно эффективны, чтобы остановить негативное воздействие алкогольных метаболитов. Одним из наиболее опасных продуктов расщепления этанола является ацетальдегид. Именно его влияние и вызывает развитие синдрома отмены, что приводит к следующим симптомам:
Именно его влияние и вызывает развитие синдрома отмены, что приводит к следующим симптомам:
- боль головы;
- плохое самочувствие;
- слабость;
- позывы на рвоту;
- тремор конечностей;
- лихорадка;
- болезненность мышц и суставов.
Как принимать глицин при интоксикации организма алкоголем
Инструкция по применению взрослым при похмелье:
Чтобы облегчить симптомы похмелья, то есть при интоксикации, рекомендуется растворить таблетку, помещая ее под язык.
Как принимать:
По 2 таблетки, через каждые полтора часа, в общей сложности до 5 раз.
Также не нужно забывать, что действие биотика кумулятивное и сверхбыстрый эффект не стоит ждать.
к содержанию ↑Вывод из запойного состояния
Благодаря своим эффективным успокаивающим способностям, медицинская кислота успешно используется врачами для лечения алкоголизма. Известно, что глицин эффективно уменьшает тягу пить и создает успокаивающие действие для нервной системы. Более того, имеет полную совместимость с другими препаратами.
Более того, имеет полную совместимость с другими препаратами.
Для лучшего эффекта, употребление таблеток лучше сопровождать употреблением желатина (мясо или рыба) или фруктового желе. Этот совет основан на том, что терапевтическая кислота включена в компоненты хрящевой ткани. И желатин содержит этот материал в объеме около 20%.
Глицин, он же аминоуксусная кислота, отлично защищает нейроны (клетки мозга) от вредоносного и разрушительного воздействия метаболитов этанола:
- Ингибирует образование глутаминовой кислоты и ГАМК. Эти соединения и вызывают общее чувство опьянения.
- Подавляет продукцию определенных соединений, которые стимулируют нейроны головного мозга при употреблении алкоголя.
Всем известно, что красное вино, допустимо в небольших количествах, но вот излишнее употребление алкоголя вызывает снижение уровня тестостерона у мужчин и вызывает и импотенцию. У женщин менструальные циклы могут быть нарушены, и затронута фертильность.
Глицин снижает вредное влияние алкоголя на организм, и ключевым словом будет именно «снижает», а не устраняет полностью. Тем не менее, нельзя забывать, что превышение дозировки, хоть и крайне редко, но способно привести к аллергическим реакциям.
Тем не менее, нельзя забывать, что превышение дозировки, хоть и крайне редко, но способно привести к аллергическим реакциям.
Особенности национального похмелья — Ведомости
Тяжелое похмелье утром 1 января — не самый лучший итог новогодней вечеринки. Чтобы утро нового года не было мучительно тяжелым, накануне стоит прислушаться к советам специалистов.
Профилактика похмелья
Нельзя пить на голодный желудок, так как в этом случае алкоголь быстро попадет в кровь. Оказавшись за новогодним столом, в первую очередь нужно чем-нибудь перекусить. В качестве закуски рекомендуется белковая пища — рыба, нежирное мясо — и сложные углеводы — картофель, рис, макароны.
Справиться с обезвоживанием, которое вызывают спиртосодержащие напитки, поможет вода. Не стоит запивать пищу алкоголем, для этого есть соки и вода. Приниматься за алкогольные напитки желательно только во время тостов.
Следует помнить, что эффект от принятой «дозы» можно ощутить сполна через 20 минут. А если «частить», то порции быстро наслоятся друг на друга и последствия станут непредсказуемыми.
А если «частить», то порции быстро наслоятся друг на друга и последствия станут непредсказуемыми.
Не следует пить слишком много шампанского и других газированных напитков. Обычно в новогоднюю ночь вслед за шампанским следуют другие напитки, и углекислый газ, который содержится в «шипучке», увеличивает всасываемость алкоголя. Поэтому, по возможности, не нужно смешивать крепкий алкоголь с шампанским, равно как и другие напитки. Если все же приходится нарушить это железное правило, то делать это надо «не понижая градуса».
Прийти в себя поможет прогулка на свежем воздухе. После этого лучше перейти на безалкогольные напитки. Кроме того, рекомендуется воздержаться от большого количества чая или кофе, которые только усугубят обезвоживание, а значит и похмелье.
Не рекомендуется злоупотреблять соленым, так как оно усилит жажду и приведет к увеличению дозы спиртного. Также не стоит налегать на сладкое — простые сахара быстро попадают в кровоток, быстро из него выводятся и ускоряют всасывание алкоголя.
Лечение похмелья
Если с утра все-таки чувствуется похмелье, нужно выпить как можно больше воды и принять активированный уголь. Кстати, в Англии XIX века вместо современных препаратов выпивали кружку теплого молока с ложкой сажи.
Утром по возможности надо слегка перекусить нежирной пищей, например, тостами. После чего выйти на свежий воздух.
Если вышеперечисленные советы не дали должного эффекта и головная боль напоминает о «вчерашнем», на помощь приходит классический рецепт — огуречный рассол. Эффективность этого самого народного средства от похмелья даже доказана специалистами. Если рассола нет, то можно заменить его на квас. Сравнительно эффективными «народными» средствами от похмелья считаются кефир, нежирное молоко, квашеная капуста, моченые яблоки, соленые огурцы, квас, томатный сок с красным (в крайнем случае — с черным) перцем.
Итальянцы «наутро» едят и пьют только белое — рис, вино, пасту — и воздерживаются от красного — колбас, помидоров и т. п. Французы варят луковый суп и крепкий черный кофе с солью. Монголы придумали бороться с похмельем поеданием маринованных овечьих глаз, которые запивают томатным соком. А вот финны спасаются сауной и холодным пивом с маринованной селедкой.
п. Французы варят луковый суп и крепкий черный кофе с солью. Монголы придумали бороться с похмельем поеданием маринованных овечьих глаз, которые запивают томатным соком. А вот финны спасаются сауной и холодным пивом с маринованной селедкой.
Как только пройдут первые симптомы похмелья, нужно плотно позавтракать и принять поливитаминные препараты, аскорбиновую кислоту, одну-две таблетки глицина, улучшающего состояние нервной системы. В течение дня лучше избегать физических нагрузок и тем более не следует заниматься спортом. Зато прогулка на свежем воздухе улучшит кровоток и ускорит выведение алкоголя из организма.
Пять секретов для победы над похмельем
Как хочется погрузиться с головой в праздник! Накануне Нового года половина соотечественников мечется, накрывая стол, а другая половина мечтает выпить. Да, первая тоже мечтает выпить, но после того, как накроет стол. Ведь всем хочется погрузиться в волшебное состояние, когда все легко и верится в чудо, а бокал вина для большинства из нас — как сигнал для наступления праздника.
Однако именно перед Новым годом, несмотря на хлопоты, стоит запастись всем тем, что спасет вас на следующий день. Ведь мало кто думает о похмелье, которое накрывает значительную часть человечества в первый день нового года.
Секрет первый. Заранее зайдите в аптеку
Запомните: никакие лекарственные препараты нельзя принимать вместе с алкоголем! А вот подготовить организм заранее можно и нужно. Так же как и помочь ему на следующий день. В первую очередь, накануне праздника нам нужны витамины группы «Б». Они усиливают выработку ферментов, которые потом помогут переработать и вывести алкоголь. Принять нужно ударную дозу: днем 31 декабря и потом еще раз часа за три до застолья. Взрослому человеку надо примерно по 100 мг активного вещества за один прием.
Можно воспользоваться также препаратом «Эссенциале», который восстанавливает клетки печени и увеличивает ее способность противостоять агрессивным факторам, таким как алкоголь и острая жирная пища. Его хорошо принимать профилактически несколько дней до Нового года и неделю после него. Но если вы поздно спохватились, то хотя бы примите две капсулы утром 31 декабря и еще две – перед началом банкета.
Но если вы поздно спохватились, то хотя бы примите две капсулы утром 31 декабря и еще две – перед началом банкета.
Также в ожидании праздника хорошо воспользоваться желчегонными растительными препаратами (принять их утром и еще раз за два часа до застолья). Это может быть препарат Liv-52, желчегонный травяной сбор или сироп шиповника.
Если у вас нет язвы, то купите аспирин, лучше шипучий, с витамином С: таблетку надо растворить в стакане воды и принять вечером 31 декабря, а еще одна скорее всего понадобится вам на следующий день после праздника.
На следующий день вам, возможно, понадобятся препараты, которые помогут избавить организм от интоксикации. Поэтому сразу купите адсорбенты. Это могут быть препараты «Энтеросгель», «Полисорб», «Смекта» или активированный уголь.
Для снятия мышечной и головной боли подойдут шипучие таблетки «Алка-Зельтцер», «Алька-Прим», «Упсарин», «Аспирин-С» или «Эффералган». Внимание! Принимать их на пустой желудок нельзя, и они категорически запрещены для язвенников.
Еще от похмельного синдрома помогают такие препараты, как глицин и цитраргинин. Первый улучшает эмоциональное состояние, избавляет от подавленности и повышает умственную активность, а второй усиливает активность печени, заставляя ее активнее выводить токсины из организма. Только принимать и тот и другой препарат надо довольно большими дозами: глицин по пять таблеток на прием, а цитраргинин по ампуле три раза в день, разводя содержимое ампулы в половине стакана воды.
Секрет второй. Грамотная периодичность тостов и понижать ли градус?
Залог позитивного праздника — грамотная периодичность тостов. Организм сжигает алкоголь в определенном темпе, примерно 10 мл за 20 минут. Дайте ему больше времени, чтобы сжечь алкоголь, и тогда меньшее количество попадет в кровь и мозг. Следите, чтобы тосты не следовали друг за другом слишком часто. Иначе все присутствующие рискуют встретить Новый год лицом в салате, а не в радостном и приподнятом настроении. Вам это надо?
А вот распространенное мнение о том, что нельзя понижать градус, требует пристального внимания. Дело в том, что для любимого россиянами напитка под названием «водка» это, безусловно, верно. Мало того, водка вообще не терпит никакого соседства с другими алкогольными напитками. Начав с шампанского, перейдя к вину и окончив водкой, вы можете испытать на следующий день тяжелейшее похмелье, хотя и не понижали градус. Так что, если уж пьете водку, начинайте и заканчивайте только ею.
Дело в том, что для любимого россиянами напитка под названием «водка» это, безусловно, верно. Мало того, водка вообще не терпит никакого соседства с другими алкогольными напитками. Начав с шампанского, перейдя к вину и окончив водкой, вы можете испытать на следующий день тяжелейшее похмелье, хотя и не понижали градус. Так что, если уж пьете водку, начинайте и заканчивайте только ею.
А для остальных напитков это правило не так важно. В европейской гастрономической традиции, например, вполне принято начинать с аперитива, которым может быть даже крепкий напиток: например, коньяк или биттер. А затем под салаты и закуски последует легкое белое вино (понижение градуса!). После него под рыбу или птицу вам предложат более плотное и сложное белое вино или легкое красное, под горячие мясные блюда — насыщенное, танинное красное вино, а в конце трапезы, на дижестив, полагаются крепленые вина или дистилляты. И никакого похмелья, хотя градус «гулял» туда-сюда!
Специалисты говорят, что важнее соблюдение другого правила: напитки должны быть однородными по сырьевому происхождению. Если уж начали с того, что делают из винограда, то и продолжайте другими напитками из винограда.
Если уж начали с того, что делают из винограда, то и продолжайте другими напитками из винограда.
Врачи категорически не советуют экспериментировать, комбинируя то, что сделано из разного сырья: например, портвейн и виски, сакэ и пиво, шампанское и водку. И, конечно, всегда надо соблюдать разумную меру. Иначе утро следующего дня добрым не будет.
Секрет третий. Правильная закуска
Алкогольные напитки требуют непременной закуски. Это известно всем. Народная мудрость гласит даже: «запивающий водку закусывает собственной печенью». И в этом есть изрядная доля правды. Только вот обильно есть совсем не обязательно.
Диетологи и токсикологи в один голос утверждают, что идеальная закуска к алкогольным напиткам должна быть быстро перевариваемой и не слишком обильной. А совсем не жирной, как принято думать в народе. Ведь жирная и острая пища негативно влияет на печень, которая и так работает на пределе своих возможностей, перерабатывая и выводя из крови алкоголь.
Лучше всего для закуски подходят квашеная капуста, овощные салаты и сочные фрукты, особенно виноград и цитрусовые. Хуже всего — жирное мясо и колбасы, а также грибы. А еще, чтобы помогать организму выводить алкоголь, нужно много безалкогольных напитков. Полезны тоники, так как они содержат хинин, поддерживающий работу печени, а также обладают мембраностабилизирующим действием, то есть уменьшают проникновение токсинов из кишечника в кровь. Хорошо также пить фруктовые соки и зеленый чай. А вот газированные сладкие напитки вроде кока-колы и лимонадов только во вред.
Секрет четвертый. Вода вам в помощь
Если вы все же переусердствовали с алкоголем, и утром вам трудно пошевельнуться, то первое, что нужно себя заставить сделать — это выпить стакан простой воды. Если дело совсем плохо, уместно поступить грубо: выпить побольше воды и вызвать рвоту. Ведь вам нужно избавиться от остатков алкоголя и токсинов. Еще один радикальный метод — обычная очистительная клизма.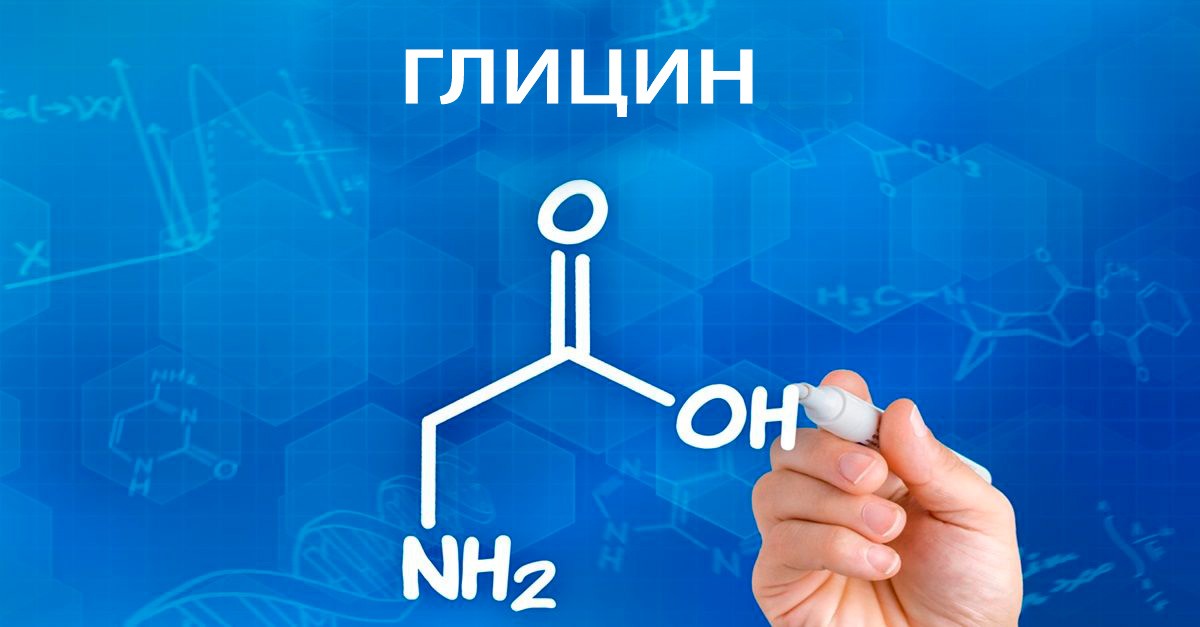 Ведь только 30% алкоголя всасывается в полости рта и в желудке, а остальная часть поступает в кровь из кишечника человека. Значит, освободив кишечник, мы сразу облегчим состояние и предотвратим дальнейшую интоксикацию.
Ведь только 30% алкоголя всасывается в полости рта и в желудке, а остальная часть поступает в кровь из кишечника человека. Значит, освободив кишечник, мы сразу облегчим состояние и предотвратим дальнейшую интоксикацию.
А после этого необходимо тщательно почистить зубы и принять душ. Конечно, в идеале нужен контрастный душ, но те, кто страдает похмельным синдромом, знают, как трудно заставить себя это сделать. Программа-минимум — хотя бы помыть голову. После этого уже станет лучше.
После водных процедур наступает время принимать адсорбенты. Все другие препараты вы примете спустя час, иначе адсорбенты нейтрализуют их действие. Выпейте «Энтеросгель», «Смекту» или активированный уголь. Чтобы вывести из организма токсины, надо пить много жидкости. Подойдут простая вода или минералка, зеленый чай, отвар шиповника, травяной мочегонный сбор, натуральный кофе (только если вы не страдаете повышенным давлением).
Через час нужно заставить себя немного поесть и только после этого принять «Аспирин-С», «Эффералган» или «Алка-Зельтцер». А еще через час — глицин. В течение всего дня следует принимать как можно больше жидкости: соки, бульоны. Хорошо также есть жидкую пищу: борщ, солянку или щи из квашеной капусты.
А еще через час — глицин. В течение всего дня следует принимать как можно больше жидкости: соки, бульоны. Хорошо также есть жидкую пищу: борщ, солянку или щи из квашеной капусты.
Секрет пятый. Занимайтесь сексом, это полезно!
Лучшая антипохмельная терапия — это утренний секс в первый день года. Тем более что многим мужчинам после обильного возлияния особенно сильно хочется заняться сексом. Этому есть вполне научное объяснение. Ведь разрушением неотработанных мужских половых гормонов занимается печень. А когда она очень занята переработкой алкоголя, то ни на что другое ее не хватает. Половые гормоны начинают накапливаться в организме и возникает повышенное сексуальное желание.
Секс способствует победе над похмельным синдромом, ведь он ускоряет процесс обмена веществ, а значит токсины активнее перерабатываются и выбрасываются организмом. К тому же во время полового акта значительно повышается уровень эндорфинов в крови и человек чувствует себя гораздо лучше. Так что не пренебрегайте намеками организма. Хочется секса — занимайтесь им, и будет вам счастье весь год!
Так что не пренебрегайте намеками организма. Хочется секса — занимайтесь им, и будет вам счастье весь год!
Татьяна Рублева
Глицин после запоя – действие и способы применения
Глицин знаком многим людям как препарат, стимулирующий умственную активность и поддерживающий эмоциональное равновесие в напряженные периоды жизни. Его принимают студенты и школьники накануне экзаменов и спортсмены во время подготовки к соревнованиям. Но не всем известна способность этой аминокислоты облегчать выход из запоя и предупреждать расстройства психики, спровоцированные употреблением алкоголя. Глицин при запоях назначается как вспомогательное средство, стабилизирующее эмоциональное состояние больного, ускоряющее детоксикацию и стимулирующее обменные процессы в нервной системе.
Что такое глицин?
Глицин (аминоуксусная кислота) относится к аминокислотам – структурным компонентам белков. В нервной системе он действует как нейромедиатор, то есть участвует в передаче нервного сигнала. Глициновые рецепторы расположены на многих участках головного и спинного мозга. Связываясь с ними, глицин оказывает тормозящее действие на нейроны и блокирует выделение аминокислот, поддерживающих возбуждение. Таким образом, Глицин относится к категории успокаивающих, легких противотревожных и антидепрессивных средств. При лечении алкогольной и опиатной абстиненции используется в качестве вспомогательного препарата со слабовыраженным ноотропным, седативным и транквилизирующим действием.
В нервной системе он действует как нейромедиатор, то есть участвует в передаче нервного сигнала. Глициновые рецепторы расположены на многих участках головного и спинного мозга. Связываясь с ними, глицин оказывает тормозящее действие на нейроны и блокирует выделение аминокислот, поддерживающих возбуждение. Таким образом, Глицин относится к категории успокаивающих, легких противотревожных и антидепрессивных средств. При лечении алкогольной и опиатной абстиненции используется в качестве вспомогательного препарата со слабовыраженным ноотропным, седативным и транквилизирующим действием.
Глицин для лечения запоя
Глицин не является лекарственным средством. Его действие при запое не подвергалось крупномасштабным научным исследованиям, но имеется богатый клинический опыт практикующих врачей, согласно которому прием таблетированной формы аминоуксусной кислоты улучшает физическое самочувствие и эмоциональное состояние больных. Глицин – биологически активная добавка, которая ускоряет и облегчает процесс выхода из запоя. Но он не может заменить дезинтоксикационные растворы для внутривенного введения, симптоматические препараты для устранения рвоты, тахикардии, судорог и других проявления алкогольного отравления.
Но он не может заменить дезинтоксикационные растворы для внутривенного введения, симптоматические препараты для устранения рвоты, тахикардии, судорог и других проявления алкогольного отравления.
Механизм действия Глицина при запое
Во время запоя в организм поступает большое количество этилового спирта, который не успевает полностью метаболизироваться. Накапливается промежуточный токсичный продукт обмена – ацетальдегид. Его излишек становится причиной головной боли, тошноты, рвоты, учащенного сердцебиения, повышения артериального давления, жажды. Молекулы глицина вступают в реакцию ацетальдегидом, в результате чего образуется ацетилглицин – безвредное соединение, участвующее в производстве белков, ферментов и гормонов. Поэтому глицин при запое действует как дезинтоксикационный препарат.
Другой эффект от его приема – усиление процессов торможения в нервной системе. Глицин связывается со специфическими рецепторами нейронов, блокирует выведение из клеток глутаминовой кислоты, «возбуждающей» нервную ткань, и усиливает выброс гамма-аминомасляной кислоты, оказывающей тормозящий эффект. В результате больной становится более спокойным и эмоционально уравновешенным.
В результате больной становится более спокойным и эмоционально уравновешенным.
Эффект от приема Глицина
Спиртные напитки нарушают функциональность нервных клеток: развиваются головные боли, головокружения, тремор, поведение становится неконтролируемым, мысли – спутанными. Глицин уменьшает вредное влияние алкоголя на мозг. Согласно инструкции к препарату его применение во время запоя приводит к следующим эффектам:
- уменьшается нервозность и раздражительность
- нормализуется настроение, редуцируются симптомы депрессии
- нормализуется обмен веществ, улучшается самочувствие
- ослабляется тяга к алкоголю
- становятся менее выраженными проявления интоксикации
Способ применения Глицина
Глицин после запоя рекомендуется принимать по 1-2 таблетки каждый час или каждые полтора часа. Максимальная суточная доза – 10 таблеток (1 грамм). Ее превышение способно вызвать расстройство пищеварения, но не ускорит процесс выздоровления. Если ежечасный прием препарата не возможен, то допустимо принимать по 3 таблетки 3 раза в день в сочетании с витаминами группы B, улучшающими усвоение аминокислоты.
Если ежечасный прием препарата не возможен, то допустимо принимать по 3 таблетки 3 раза в день в сочетании с витаминами группы B, улучшающими усвоение аминокислоты.
Таблетку Глицина нужно класть под язык и рассасывать до полного растворения. Терапевтический курс длится от 2 до 4 недель. Если запои регулярны, то лечение стоит повторить спустя 10 дней. Глицин, принятый за 1-2 часа до алкоголя, уменьшает вредное влияние этилового спирта на организм.
Ограничения и побочные эффекты Глицина
Глицин без опаски используется для лечения и профилактики запоев так как практически не имеет побочных эффектов. Единственное противопоказание – индивидуальная непереносимость аминоуксусной кислоты. Она проявляется расстройством функций желудочно-кишечного тракта (легкой диареей) и местными аллергическими реакциями, чаще всего – зудом. Большинство пациентов, вне зависимости от возраста и состояния здоровья, хорошо переносят препарат. Стоит также помнить, что Глицин понижает активность нейролептиков, антидепрессантов, противоэпилептических средств, анксиолитиков и снотворных средств.
Выводы
Глицин при запоях применяется как вспомогательное терапевтическое средство. Клинический опыт доказывает, что препарат улучшает эмоциональное состояние пациентов, ускоряет процесс детоксикации, восстанавливает обменные процессы в нервной системе. В результате курсового лечения уменьшается раздражительность и подавленность больных, быстрее устраняются симптомы отравления, снижается тяга к употреблению спиртных напитков. Оптимальный режим приема Глицина: по 1-2 таблетки каждый час (не более 10 шт/день). Курс лечения – 2-4 недели. Стоит помнить, что препарат не имеет побочных эффектов, но может оказаться индивидуально непереносимым.
топ-5 способов борьбы россиян с тревогой. Инфографика :: Общество :: РБК
Общение с близкими — это главный метод борьбы со стрессом у россиян, показывает исследование «Ромира». А еще, чтобы бороться со стрессом, россияне любят есть сладкое, пить алкоголь и гладить животных
- Для борьбы со стрессом мужчины предпочитают употреблять алкоголь (25% популярности среди мужчин и 17% в общей выборке) и проводить время с друзьями или родственниками. Женщины также проводят время с родными и друзьями, но алкоголю предпочитают сладкое.
- Наиболее популярными алкогольными напитками стали пиво и вино, за ними идут водка и джин. Среди сладостей наибольшее предпочтение россияне отдают печенью, далее следуют конфеты-батончики, шоколад и торты.
- В топ-3 успокоительных препаратов вошли глицин, корвалол, валериана.
- Среди тех россиян, которые борются со стрессом с помощью домашних животных, 42% предпочитают собак, 33% — кошек.
Подробнее о том, что вызывает тревогу у россиян и что помогает им с ней бороться — в материале РБК.
Ученые назвали средство от магнитных бурь — Российская газета
Чем опасны изменения космической погоды? И можно ли снизить негативное влияние магнитных бурь на человека? Об этом корреспондент «РГ» беседует с руководителем лаборатории метеопатологии и магнитобиологии Научного клинического центра доктором медицинских наук Юрием Гурфинкелем.
Юрий Ильич, магнитными бурями нас пугают с завидной регулярностью. Они действительно так опасны для здоровья?
Юрий Гурфинкель: На возмущение геомагнитного поля реагируют абсолютно все. Другое дело — как. Если человек молод и здоров, то в дни магнитных бурь он может почувствовать лишь легкую сонливость. Гораздо хуже пожилым и хроникам. В группе риска прежде всего пациенты с сердечной патологией и перенесшие нарушения мозгового кровообращения.
Механизмы влияния космической погоды на человека до конца еще не изучены. Однако есть данные из Франции, Германии, Китая, Израиля, Литвы, Грузии, ряда российских клиник — везде, где наблюдались больные с ишемией сердца, во время магнитных бурь возрастало примерно в два раза не только число инфарктов, но и смертность от них.
Что происходит с человеком?
Юрий Гурфинкель: Наш организм — это своего рода биокомпьютер, чувствительность которого наиболее высока в диапазоне магнитного поля Земли (до 50 микротесл). Джозеф Киршвинк и его коллеги из Калифорнийского технологического института установили наличие кристаллов магнетита в тканях мозга приматов и в надпочечниках человека. Количество этих наночастиц, восприимчивых к магнитному полю, составляет от одного до 10 миллионов на один грамм ткани. Они «улавливают» колебания в магнитосфере Земли. А это приводит, например, к выбросу адреналина и глюкокортикоидов (гормонов надпочечников).
Адреналин, в свою очередь, повышает свертываемость крови. Во время магнитных бурь она возрастает. Замедляется течение крови в капиллярах. А эритроциты, выражаясь научным языком, агрегируют, то есть склеиваются. Это увеличивает риск образования тромбов. Ухудшается общее состояние, нарушается работа сердечно-сосудистой системы и кровотока в мозгу. Наши исследования показали: движущаяся по сосудам кровь сама по себе может являться сенсором магнитных полей, поскольку красные кровяные клетки содержат гемоглобин и в силу этого обладают магнитным моментом.
Поэтому во время бури нарушается сон и возрастает число психических нарушений?
Юрий Гурфинкель: Геомагнитные возмущения организм человека воспринимает как стресс. Меняется гормональный фон в организме, нарушается работа сосудистой системы, в том числе и в сосудах головного мозга. При этом резко снижается выработка мелатонина, который продуцирует шишковидная железа, располагающаяся в центре головного мозга и управляющая процессами бодрствования и сна. В это время даже здоровый человек может пережить перепады настроения, а у некоторых это сопровождается всплесками агрессии.
Кто особенно подвержен влиянию магнитных бурь?
Юрий Гурфинкель: От 10 до 20 процентов молодых людей имеют повышенную чувствительность к геомагнитным возмущениям. Но чем старше человек, тем болезненней организм реагирует на плохую космическую погоду. Примерно 40 процентов 60-летних плохо себя чувствуют из-за волнения магнитного поля Земли. А среди 70-летних- уже больше чем у половины.
А как я могу понять, что источник моего недуга — повышенная солнечная активность?
Юрий Гурфинкель: Если «земная» причина ухудшения состояния исключена, специалисты советуют последить за прогнозом космической погоды. Например, есть портал Института земного магнетизма, ионосферы и распространения радиоволн РАН. Если вы заметили, что вечером «прихватило» сердце, стоит проверить: не было ли в это время магнитной бури. Когда таких совпадений будет 8 из десяти, можно говорить о том, что у вас повышенная восприимчивость. Однако не стоит ориентироваться на долгосрочные прогнозы, которые печатают некоторые СМИ. Достаточно достоверную картину можно составить только на неделю вперед.
Можно ли избежать влияния магнитных бурь на организм?
Юрий Гурфинкель: Нет. Однако можно соблюдать нехитрые правила, чтобы минимизировать негативный эффект. Например, в тяжелые дни лучше ограничить физическую нагрузку, избегать нарушений режима сна, командировок с длительными перелетами. Желательно чаще находиться на свежем воздухе, употреблять достаточное количество жидкости, овощей и фруктов. Важно измерять артериальное давление, вовремя принимать лекарства. Пожилым людям крепкие алкогольные напитки лучше исключить вообще. Но если здоровье позволяет, можно выпить бокал натурального красного вина. Из-за интенсивной выработки адреналина магнитные бури могут вызвать повышенный аппетит. В таком случае не надо увлекаться мясной пищей, лучше обратить внимание на овощи, фрукты, рыбу.
А что делать, если во время магнитной бури «прихватило» прямо на работе?
Юрий Гурфинкель: Самое простое средство — принять обычный аспирин. Во время магнитной бури наиболее эффективен прием 150-200 мг этого препарата. Или комбинацию аспирина и плавикса (75 мг). Но только в том случае, если это пациенту не противопоказано, так как бесконтрольный прием этих препаратов может спровоцировать эрозии и даже язву желудка. Можно рекомендовать прием глицина. Эта незаменимая аминокислота благоприятно действует на головной мозг, уменьшает повышенную возбудимость.
Справка «РГ»
Во время солнечной вспышки всего за несколько минут происходит гигантский выброс энергии и к Земле отправляются потоки заряженных частиц. А через два-три дня нашей планеты достигают облака плазмы, которые и провоцируют бури — возмущение магнитного поля Земли. Периоды солнечной активности повторяются в среднем раз в 11 лет. И сейчас мы переживаем как раз один из них. По оценкам специалистов, в среднем магнитные бури случаются 5-6 раз в год и длятся 2-3 дня.
Глицин и алкоголь — совместимость, последствия
Глицин – это аминокислота, обладающая седативными транквилизирующим действием. Она активизирует обмен веществ, выводит из организма токсины, в том числе после принятия спиртного. Но насколько совместимы Глицин и алкоголь? Можно ли принимать их одновременно? Если соблюдать некоторые рекомендации, то препарат способен значительно облегчить состояние опьянения и похмельный синдром.
Свойства Глицина
Глицин выпускается в форме таблеток, имеет доступную цену и продается в каждой аптеке. Лекарство пользуется большой популярностью из-за широкого перечня свойств. Оно применяется при недомоганиях и для стимуляции организма:
- оказывает успокаивающее, антидепрессивное действие при психоэмоциональном перенапряжении;
- имеет слабовыраженный транквилизирующий эффект;
- способствует улучшению умственной деятельности, памяти и внимания, повышает работоспособность;
- регулирует обмен веществ;
- нейтрализует действие лекарственных и спиртных токсинов, сокращает поражение головного мозга алкоголем, выступает в роли вспомогательного средства при лечении алкоголизма;
- активизирует защиту центральной нервной системы;
- улучшает настроение, выводит из стрессового состояния, уменьшает конфликтность, раздражительность;
- способствует нормализации сна, облегчает процесс засыпания;
- применяется при ишемическом инсульте;
- уменьшает последствия черепно-мозговых травм;
- оказывает благотворное воздействие при вегетососудистой дистонии;
- снижает влечение к сладкому.
Взаимодействие с алкоголем
Лекарственные средства не рекомендуется смешивать вместе с алкоголем. Глицин следует пить до или после спиртных напитков. Для замедления процесса опьянения допускается одновременный прием.
Глицин с алкоголем можно употреблять во время застолья по 1 таблетке каждые 1,5-2 часа. Такое соединение не даст спиртному оказать сильное разрушающее влияние на клетки мозга. Но при этом следует наблюдать за реакцией организма, так как не исключается обратная реакция и непредсказуемые последствия.
Глицин и алкоголь могут привести к наркотическому опьянению, агрессии, дезориентации и другим побочным эффектам вплоть до отравления. Поэтому принимать Глицин с алкоголем нужно очень осторожно, без проявления фанатизма.
Нагрузки алкоголя на организм должны контролироваться. Нельзя возлагать все надежды на один Глицин. Он может смягчить удар негативного влияния на здоровье, но неспособен полностью предотвратить все разрушающие последствия. Поэтому не стоит проводить соревнования между действием алкоголя и эффективностью препарата. Такие испытания чреваты серьезными нарушениями функций органов, а также хроническими заболеваниями.
Помощь при систематическом употреблении алкоголя
Глицин при алкоголизме назначают для лечения пациентов со стойкой зависимостью. Попадая в организм, его компоненты усиливают метаболизм, что ускоряет процесс выведения этанола. Препарат ликвидирует токсическое действие алкоголя на печень, почки, желудочно-кишечный тракт и на работу мозга. Таким образом, он выводит пациента из запойного состояния, снимает опьянение.
Во время длительного приема алкогольных напитков многие зависимые перестают нормально питаться, некоторые вовсе отказываются от пищи. Происходит это из-за высокой калорийности спиртных напитков. Организм воспринимает их как единственный источник питательных элементов и быстро впитывает содержимое. Это приводит к интоксикации, дисфункции ЖКТ, нарушению обмена веществ, интоксикации.
Для чего нужен Глицин алкоголикам? Препарат не только выводит токсины. Он разрушает замкнутый круг, состоящий из приема спиртного и похмелья. Благодаря Глицину опьянение ослабевает, алкоголь оперативно выводится. Человек не чувствует при этом дискомфорта, вызванного похмельным синдромом, который в обычной ситуации побуждает снова принимать крепкие напитки как панацею. Желание употреблять алкоголь угасает. Человек начинает ощущать голод и тягу к обычной еде, постепенно начинает нормально есть.
Влияние Глицина на эмоциональное состояние алкоголика при выходе из запоя не менее значимо. Средство подавляет агрессию, раздражительность, вызванные похмельным синдромом, улучшает социальную адаптацию, положительно воздействует на настроение. Это очень важно для нестабильной психике зависимого от алкоголя человека, который привык топить в спиртном все негативные эмоции.
Как действовать при интоксикации организма
Принимать Глицин при алкогольной интоксикации нужно по следующей схеме: 1 таблетка (100 мг) через каждый час 5-7 раз за сутки или три приема по 2 таблетки с интервалом в 3 часа, в зависимости от степени тяжести отравления. Препарат ложится под язык и рассасывается до полного растворения. Согласно инструкции его можно принимать в виде порошка, раздробив пилюлю. Главное – не переусердствовать и не допустить передозировки, которая влечет за собой непредвиденные последствия.
Не менее эффективно помогает Глицин от запаха алкоголя. Приняв 2 таблетки утром после застолья, можно быстро избавиться от амбре и смело отправляться на работу. Никакие похмельные синдромы не будут мешать трудовому процессу. Глицин вернет организм в нормальное физическое и психоэмоциональное состояние.
Противопоказания и побочные эффекты
Совместимость Глицина и алкоголя допустима, но не для всех. Как любой другой медикамент он способен вызвать аллергическую реакцию, которая обостряется при употреблении спиртного. К тому же не исключена индивидуальная непереносимость вспомогательных компонентов препарата.
Если наблюдается нестабильность артериального давления или падение его ниже обычного уровня, следует прекратить прием Глицина. Дальнейшее употребление и совмещение средства с алкоголем должны быть согласованы с врачом.
Несмотря на то что Глицин назначают для нормализации сна, в пределах допустимых доз он не вызывает сонливости, потери внимания, заторможенности. К тому же препарат проявляет хорошую совместимость с различными лекарственными медикаментами. Поэтому нет необходимости отменять назначенное лечение при каких-либо заболеваниях.
Источники:
Видаль: https://www.vidal.ru/drugs/glycine__4429
ГРЛС: https://grls.rosminzdrav.ru/Grls_View_v2.aspx?routingGuid=80b010cc-df9b-4601-90e4-d8cd11a43524&t=
Нашли ошибку? Выделите ее и нажмите Ctrl + Enter
Глицин предотвращает вызванное алкоголем повреждение печени за счет снижения содержания алкоголя в желудке крысы
Предпосылки и цели: Инактивация клеток Купфера предотвращает вызванное алкоголем повреждение печени, и в этот механизм, вероятно, вовлечена гипоксия, возникающая после гиперметаболического состояния, вызванного активированными клетками Купфера. Известно, что глицин предотвращает реперфузионное повреждение печени. Целью этого исследования было определить, предотвращает ли глицин вызванное алкоголем повреждение печени in vivo.
Методы: Самцы крыс Wistar подвергались воздействию этанола (10-12 г / кг-1 день-1) непрерывно в течение до 4 недель по протоколу внутрижелудочного кормления. Влияние глицина на метаболизм этанола при первом прохождении также исследовали in vivo, и влияние на метаболизм алкоголя оценивали конкретно в перфузированной печени.
Полученные результаты: Глицин резко снижает концентрацию этанола в моче, дыхании, периферической крови, портальной крови, кале и содержимом желудка.Уровни аспартатаминотрансферазы в сыворотке повысились до 183 Ед / л после 4 недель лечения этанолом. Напротив, значения были значительно ниже у крыс, получавших глицин вместе с этанолом. Глицин также значительно уменьшил стеатоз и некроз печени. Глицин резко увеличивал элиминацию этанола при первом прохождении in vivo, но не влиял на метаболизм алкоголя в перфузируемой печени.
Выводы: Глицин сводит к минимуму вызванное алкоголем повреждение печени in vivo, предотвращая попадание этанола в печень, активируя метаболизм первого прохождения в желудке.
Глицин как лечебный иммуно-нутриент при алкогольной болезни печени
Активация клеток Купфера эндотоксином кишечного происхождения является важным фактором гепатотоксичности этанола. Кроме того, было показано, что этанол модулирует как экспрессию, так и активность нескольких внутриклеточных сигнальных молекул и факторов транскрипции в клетках Купфера, а постоянное лечение этанолом повышает чувствительность клеток Купфера к эндотоксину.Эти данные свидетельствуют о том, что ингибирование активации клеток Купфера эффективно для клинического применения при алкогольном гепатите. В последнее время накапливаются данные, свидетельствующие о возможности использования глицина в качестве иммуномодулирующей аминокислоты. Было показано, что диета, содержащая глицин, улучшает выживаемость при эндотоксиновом шоке, предотвращая активацию клеток Купфера. Глицин, скорее всего, предотвращает индуцированное ЛПС повышение внутриклеточной концентрации Са в клетках Купфера, тем самым сводя к минимуму передачу сигналов рецептора ЛПС и продукцию цитокинов.Действительно, глицин предотвращает вызванное алкоголем повреждение печени у крыс, длительное время кормящих этанолом (Tsukamoto-French), за счет снижения выработки TNF-альфа в печени. Более того, глицин защищает от апоптоза синусоидальных эндотелиальных клеток (SEC), который является одним из начальных событий в развитии повреждения печени. С другой стороны, эпидемиологические данные определили, что хроническое употребление алкоголя является значительным фактором риска канцерогенеза. Интересно, что глицин подавляет рост опухоли in vivo, скорее всего, из-за подавления ангиогенеза.Было показано, что ингибирующий эффект глицина на рост и миграцию эндотелиальных клеток обусловлен активацией глицинового канала Cl. Предполагается, что открытие этого анионного канала гиперполяризует клеточную мембрану, блокирует приток Са через потенциалзависимый канал Са, тем самым притупляя передачу сигналов, опосредованную фактором роста. Таким образом, глицин можно использовать не только для лечения алкогольного гепатита, но и для химиопрофилактики и лечения гепатоцеллюлярной карциномы при алкогольном циррозе печени.В совокупности можно сделать вывод, что глицин является мощным терапевтическим иммуно-питательным веществом при различных хронических заболеваниях печени, включая алкогольную болезнь печени (ALD).
S02-2РОЛЬ ГЛИЦИНОВЫХ РЕЦЕПТОРОВ В ПОВЕДЕНИИ ПОТРЕБЛЕНИЯ ЭТАНОЛА | Алкоголь и алкоголизм
Аминокислота таурин, как ранее было показано, влияет на вызванное этанолом повышение уровня дофамина в системе вознаграждения мозга, скорее всего, через рецепторы глицина, расположенные в прилежащем ядре.Было также показано, что эти рецепторы глицина влияют на добровольное потребление этанола как у крыс, так и у людей после введения ингибитора захвата глицина. Была выдвинута гипотеза, что этанол в первую очередь действует в ядре accubens, где вызывающий привыкание наркотик вызывает увеличение эндогенного таурина. Повышение уровня таурина в сочетании с этанолом активирует нейронную цепь, включающую глициновые рецепторы прилежащего ядра, а также никотиновые рецепторы ацетилхолина, расположенные в передней вентральной тегментальной области, чтобы вызвать повышение уровня дофамина в прилежащем ядре.
В настоящем исследовании мы провели пять отдельных экспериментов, в которых крыс линии Вистар проверяли на предмет добровольного потребления этанола в модели прерывистого потребления этанола. После семи недель потребления этанола крыс с самым высоким и самым низким потреблением этанола снабжали зондами для микродиализа in vivo в прилежащем ядре. В день диализного эксперимента все крысы получили острую инъекцию этанола, и у каждого животного контролировали внеклеточные уровни как дофамина, так и таурина.В соответствии с литературными данными мы обнаружили, что крысы с самым высоким потреблением этанола имели значительно более низкий эндогенный тонус дофамина в прилежащем ядре по сравнению с крысами с низким потреблением этанола. Эти животные с высоким потреблением этанола также ответили на острую инъекцию этанола более медленным и ослабленным повышением таурина. Это интересно, поскольку подчеркивает участие тауриновых и прилежащих рецепторов глицина в произвольном потреблении этанола.
Таким образом, настоящее исследование предполагает, что рецепторы таурина и глицина важны как для повышения дофаминовых свойств этанола, так и для поведения при потреблении этанола.
© Автор, 2017. Медицинский совет по алкоголю и Oxford University Press. Все права защищены
Глицин и пероральный D-циклосерин у больных алкоголизмом и здоровых субъектов — Просмотр полного текста
Вопрос № 1: Улучшит ли глицин когнитивные нарушения? Гипотеза № 1. Основываясь на положительных результатах, полученных с глицином и милацемидом, пролекарством глицина, при шизофрении и деменции, мы ожидаем, что глицин улучшит когнитивный дефицит.
Вопрос № 2: Будут ли пациенты-алкоголики проявлять усиленные эндокринные эффекты глицина? Гипотеза № 2: Основываясь на дозозависимых эффектах глицина у здоровых людей, мы ожидаем, что глицин будет увеличивать эндокринный ответ на глицин у пациентов-алкоголиков с предположительно дисрегулируемой функцией рецепторов NMDA.
Вопрос № 3: Будет ли D-циклосерин иметь эффекты, подобные этанолу? Гипотеза № 3: Если подавление функции рецептора NMDA является фундаментальным для субъективных эффектов этанола, то свойства антагониста NMDA D-циклосерина следует признать этанолоподобными (по сравнению с плацебо) у недавно прошедших детоксикацию алкоголиков и здоровых субъектов.
Вопрос № 4: Обратит ли D-циклосерин когнитивные преимущества глицина? Гипотеза 4. Основываясь на дозозависимой активности антагониста NMDA D-циклосерина, мы ожидаем, что D-циклосерин будет конкурировать с агонистической активностью глицина и, следовательно, будет обращать вспять когнитивные преимущества глицина.
Вопрос № 5: Подавляет ли D-циклосерин эндокринные эффекты глицина? Гипотеза № 5: Если агонистическая активность глицина необходима для определения эндокринной реакции, то дозозависимые свойства антагониста NMDA D-циклосерина должны блокировать эти эффекты.
Целью данного исследования является изучение взаимодействия между глицином и D-циклосерином у больных алкоголизмом и здоровых людей. Доклинические исследования показали, что соединения, действующие на глициновый сайт рецепторного комплекса N-метил-D-аспартат (NMDA), такие как глицин, могут обращать действие этанола на функцию рецептора NMDA (Rabe et al., 1990). Аминокислота глицин является коагонистом рецепторного комплекса NMDA (Kemp et al., 1993). Он связывается с нечувствительным к стрихнину сайтом и положительно модулирует рецептор NMDA (Mc Donald et al., 1990). Физиологически сайт глицина не насыщен, и введение глицина может усиливать опосредованные рецептором NMDA ответы. Напротив, D-циклосерин (Hood et al., 1989) является частичным агонистом глицинового сайта рецептора NMDA с дозозависимыми антагонистическими свойствами. Антагонистическая активность D-циклосерина в отношении NMDA должна вызывать эффекты, подобные этанолу, которые могут быть отменены агонистом глицином. Это исследование предназначено для оценки возможного вклада глицинового сайта в снижение когнитивного дефицита при алкоголизме и дополняет текущую работу VA Connecticut Healthcare System по антагонистам NMDA у алкоголиков и здоровых субъектов.
Нацелена на обратный захват глицина при поиске алкоголя и рецидивах
Abstract
Недавно было продемонстрировано, что фармакологическая блокада транспортера глицина 1 (GlyT1) снижает потребление алкоголя и снижает рецидивы у крыс. Целью настоящего исследования было дальнейшее изучение роли GlyT1 в алкогольном рецидивирующем поведении. Для этой цели мы использовали три различных блокатора GlyT1 — SSR504734, A-1246399 и RO4993850 — и протестировали их влияние на потребление алкоголя и рецидивы потребления.Были использованы две поведенческие модели: модель эффекта алкогольной депривации и модель восстановления, вызванного сигналом. Наши данные показывают, что все три блокатора GlyT1 снижают потребление алкоголя, подобное рецидиву, и вызывают минимальные побочные эффекты или не вызывают их отсутствия, что измеряется по изменениям активности в домашней клетке, потреблению воды и массе тела. В тесте на восстановление блокаторы GlyT1 полностью исключали реакцию поиска алкоголя. Кроме того, мы протестировали другие ассоциации наркотиков / сигналов и обнаружили, что реакция на поиск кокаина также отменяется блокадой GlyT1.Наши данные подтверждают, что GlyT1 может быть использован в качестве мишени для разработки новых противоэрозионных и противорецидивных препаратов.
Введение
Утвержденные фармакологические методы лечения расстройства, связанного с употреблением алкоголя, имеют ограниченную эффективность, и требуются новые лекарственные препараты. Глицинергическая система из-за ее взаимодействия с несколькими эффектами, вызванными алкоголем, является одной из предварительных целей, и действительно появляется все больше доказательств того, что воздействие на глицинергическую систему может препятствовать потреблению алкоголя и рецидивам алкоголя (Molander et al., 2005; 2007; Vengeliene et al., 2010; Lidö et al., 2012, 2017). Это неудивительно, поскольку известно, что активность дофаминергической системы подкрепления в значительной степени находится под контролем нескольких систем нейромедиаторов, на которые напрямую воздействует алкоголь, включая стрихнин-чувствительные рецепторы глицина (GlyR) и N-метил-d-аспартат. рецепторы (NMDAR), которые также имеют сайт связывания глицина (Molander, Söderpalm, 2005; Vengeliene et al., 2008; Spanagel, 2009; Jonsson et al., 2014; Clarke et al., 2015; Söderpalm et al., 2017).
Одной из ключевых мишеней для модуляции глицинергической активности является переносчик глицина 1 (GlyT1). Генетическая инактивация или фармакологическая блокада GlyT1, как было показано, приводит как к глицинергическому сверхингибированию, так и к облегчению ответов NMDAR через сайт связывания глицина (Chen et al., 2003; Gomeza et al., 2003). Блокада GlyT1 с помощью Org-25935 снижает как исходное, так и рецидивирующее потребление алкоголя (Molander et al., 2007; Vengeliene et al., 2010; Lidö et al., 2017). Этот блокатор действует в основном через GlyT1 и, как известно, значительно повышает уровни внеклеточного глицина после его системного введения (Lidö et al., 2009). Однако доклинические данные с Org-25935 не были подтверждены в недавнем клиническом исследовании. Многоцентровое рандомизированное двойное слепое плацебо-контролируемое клиническое исследование Org-25935 не показало преимущества этого лечения по сравнению с плацебо в предотвращении чрезмерного употребления алкоголя и никакого влияния на какие-либо параметры, связанные с рецидивом (de Bejczy et al., 2014) .Это вызвало сомнения в том, может ли нацеливание на GlyT1 быть полезным для лечения наркозависимости и насколько хорошо доклинические данные отражаются в клинических условиях. С другой стороны, неудача клинического исследования могла быть вызвана такими факторами, как неадекватная дозировка или недостаточная мощность [см. De Bejczy et al. (2014) для обсуждения дальнейших ограничений клинического исследования].
К настоящему времени Org-25935 был протестирован в двух доклинических исследованиях, в обоих из которых использовались добровольно потребляющие алкоголь крысы либо в исходных условиях, либо в условиях рецидива (Molander et al., 2007; Vengeliene et al., 2010; Lidö et al., 2017). Очевидно, что для более полного анализа этой фармакологической мишени необходимо получить больше информации с помощью доклинических исследований. Таким образом, целью данного исследования было выяснить, могут ли различные блокаторы GlyT1 изменить поведение, связанное с поиском алкоголя и схожее с рецидивами. С этой целью мы провели исследование зависимости зависимости от дозы трех блокаторов GlyT1: SSR504734 (Depoortère et al., 2005), A-1246399 и RO4993850 (Kohli et al., 2016).Для изучения эффекта блокады GlyT1 были использованы две модели поведения: модель эффекта алкогольной депривации (ADE) и модель восстановления, вызванного сигналом. В модели ADE подобное рецидиву употребление алкоголя измеряется как устойчивое временное увеличение добровольного потребления алкоголя после периода депривации. После повторяющихся фаз депривации алкогольное поведение становится компульсивным (Vengeliene et al., 2013, 2014), что похоже на ситуацию рецидива у пациентов-алкоголиков. Доклинические исследования показали, что подострое лечение блокатором GlyT1 Org-25935 или антагонистами NMDAR ослабляет экспрессию ADE (Vengeliene et al., 2005, 2010).
В модели восстановления повторное приобретение реакции на оперантную задачу исследуется путем повторного воздействия на крыс экологических стимулов, связанных с алкоголем (De Wit and Stewart, 1981; Sanchis-Segura and Spanagel, 2006; Bossert et al., 2013) . Было показано, что несколько антагонистов NMDAR ослабляют поиск алкоголя, вызванный сигналом, но часто также вызывают нарушение моторного поведения (Bäckström and Hyytiä, 2004; Bachteler et al., 2005; Spanagel, 2009; Holmes et al., 2013).Самый эффективный способ уменьшить поведение, связанное с поиском лекарств, хотя этот рецептор, по-видимому, заключается в нацеливании на его сайт связывания глицина (Bäckström and Hyytiä, 2004, 2006). На сегодняшний день соединения, которые мешают GlyRs или GlyT1, не тестировались на индуцированном сигналом восстановлении поведения, связанного с поиском алкоголя или кокаина.
Наша рабочая гипотеза заключалась в том, что GlyT1 является жизнеспособной мишенью для лечения тяги к алкоголю и рецидива, и что тестирование блокаторов GlyT1 с несвязанными базовыми химическими структурами в моделях ADE и восстановления подтвердило бы эту гипотезу.Мы также проверили профиль побочных эффектов трех блокаторов GlyT1 на животных, которые длительно злоупотребляли алкоголем. И, наконец, мы проверили, является ли GlyT1 селективной мишенью для лечения алкогольной зависимости или другие наркотики, вызывающие злоупотребление, также будут затронуты блокадой GlyT1.
Методы
Животные
Девяносто пять двухмесячных крыс-самцов линии Wistar (из нашей собственной племенной колонии в CIMH, Мангейм, Германия) были использованы для экспериментов ADE, тридцать три двухмесячных самца крыс Wistar крыс (Harlan Laboratories, Venray, Нидерланды) использовали для экспериментов по восстановлению, вызванных этаноловым сигналом, и пятнадцать двухмесячных самцов крыс Sprague-Dawley (Charles River, Sulzfeld, Германия) использовали для экспериментов по восстановлению, вызванного кокаиновым сигналом.Всех животных содержали индивидуально в стандартных клетках для крыс (Ehret, Emmendingen, Germany) при 12-часовом искусственном цикле свет-темнота. Стандартный лабораторный корм для крыс (Ssniff Spezialdiäten GmbH, Soest, Германия) и водопроводная вода предоставлялись ad libitum в течение всего экспериментального периода (если не указано иное). Все экспериментальные процедуры одобрены Комитетом по уходу за животными и их использованию (Regierungspräsidium Karlsruhe, Карлсруэ, Германия) и проводились в соответствии с местным Законом о защите животных и Директивой Совета Европейских сообществ от 24 ноября 1986 г. (86/609 / EEC). ).
Наркотики
Питьевые спиртовые растворы готовили из 96% этанола (Sigma-Aldrich, Тауфкирхен, Германия), а затем разбавляли водопроводной водой. Гидрохлорид кокаина (Sigma-Aldrich) растворяли в стерильном физиологическом растворе. SSR504734 (предоставленный AbbVie, Людвигсхафен, Германия) растворяли в воде для инъекций (aqua ad iniectabilia; Braun, Melsungen AG, Германия). A-1246399 (предоставленный AbbVie) растворяли в полиэтиленгликоле 400 (PEG 400; Sigma-Aldrich Co., St.Louis, MO), а затем разбавили водой для инъекций до конечной концентрации PEG 400, равной 20%. SSR504734 и A-1246399 были свежеприготовленными и вводились внутрибрюшинно в объеме 3 мл / кг. RO4993850 был щедро предоставлен F. Hoffmann-La Roche (Базель, Швейцария) в качестве готового к использованию материала и вводился перорально в объеме 5 мл / кг.
Долгосрочное добровольное употребление алкоголя с повторяющимися фазами депривации
После 2 недель привыкания к комнате для животных крысам предоставили неограниченный доступ к водопроводной воде и 5, 10 и 20% растворам этанола (об. / Об.) В качестве хорошо.Употребление алкоголя и воды контролировали ежедневно / еженедельно путем взвешивания бутылок. На основе этих данных было рассчитано потребление воды (миллилитры на килограмм веса тела в день; мл / кг в день) и потребление алкоголя (граммы чистого спирта на килограмм веса тела в день; г / кг в день). Первый двухнедельный период депривации вводился после 8 недель непрерывного употребления алкоголя. После периода депривации крысам снова давали доступ к алкоголю и случайным образом вводили еще четыре-шесть периодов депривации.Длительная процедура добровольного употребления алкоголя, включая все фазы депривации, длилась около 1 года.
Употребление алкоголя при рецидиве (фармакологические исследования I).
Для изучения влияния медикаментозного лечения на экспрессию ADE крыс разделили на группы ( n = 7–8 на группу) таким образом, чтобы среднее исходное общее потребление алкоголя было примерно одинаковым в каждой группе. Исходное употребление алкоголя контролировалось ежедневно в течение 1 недели. После последнего дня базовых измерений бутылки с этанолом вынимали из клеток, оставляя животным свободный доступ к пище и воде в течение 2 недель.После этого первые восемь групп животных были подвергнуты в общей сложности пяти внутрибрюшинным инъекциям (один раз каждые 12 часов) либо носителя, либо SSR504734 (1, 3 и 10 мг / кг) или A-1246399 (3, 10 и 30 мг / кг). мг / кг). Остальные четыре группы получали три приема внутрь (один раз в день) либо носителя, либо RO4993850 (3, 10 и 30 мг / кг). Дозы SSR504734 были выбраны в соответствии с ранее опубликованными исследованиями (Depoortère et al., 2005). Дозы A-1246399 были выбраны на основе увеличения глицина в префронтальной коре головного мозга крыс, измеренного с помощью микродиализа, что, соответственно, дало увеличение по сравнению с исходным уровнем, равным 1.3 и 1,8 раза при 3 и 10 мг / кг внутрибрюшинно соответственно. Эти дозы значительно превышали дозы, связанные с половинной максимальной занятостью цели (1,3 мг / кг внутрибрюшинно у крыс, оцениваемой с помощью анализа связывания ex vivo) и половинной максимальной активацией NMDAR (0,8 мг / кг внутрибрюшинно у мышей; оценивалась путем отмены L-687,417-индуцированная гиперлокомоция). Дозы RO4993850 были выбраны на основе увеличения глицина в полосатом теле крысы, измеренного с помощью микродиализа, что, соответственно, дало увеличение по сравнению с исходным уровнем, равным 1.5-, 1,8- и 2,1-кратный. Самая высокая доза дала увеличение, соответствующее максимальному эффекту, индуцированному в спинномозговой жидкости здоровых добровольцев (Hofmann et al., 2016). Бутылки со спиртом были введены повторно после второго введения лекарства, и было определено возникновение ADE. Общее количество этанола (г / кг в день) и потребление воды (мл / кг в день) измеряли ежедневно в течение следующей недели.
Измерение двигательной активности в домашней клетке с помощью системы E-Motion.
Для проверки любых седативных эффектов, возникающих в результате лечения препаратом, двигательную активность в домашней клетке контролировали с помощью инфракрасного датчика, подключенного к системе записи и хранения данных (Mouse-E-Motion; Infra-e-motion, Henstedt -Ульцбург, Германия).Устройство Mouse-E-Motion помещали над каждой клеткой (30 см от дна), чтобы крысу можно было обнаружить в любом месте внутри клетки. Каждую секунду устройство производило замеры того, двигалась крыса или нет. Датчик мог обнаруживать движение тела крысы на расстояние не менее 1,5 см от одной точки отбора проб к следующей. Данные, измеренные каждым устройством Mouse-E-Motion, загружали в персональный компьютер и обрабатывали с помощью Microsoft Excel. Мониторинг двигательной активности начался за 4 дня до процедуры лечения препаратом и продолжался еще четыре дня после лечения.Процент двигательной активности каждой крысы во время и после дней лечения рассчитывали с использованием данных активности «до лечения» в качестве эталона.
Возобновление употребления алкоголя, вызванное иском
Аппарат для самостоятельного введения оперантного алкоголя.
Восстановление поиска алкоголя, вызванное репликами, проводилось в операционных камерах (MED Associates Inc., Сент-Олбанс, штат Вирджиния), заключенных в вентилируемые звукопоглощающие кабины. Камеры снабжены ответным рычагом на каждой боковой панели камеры.Реакция на активный рычаг активировала шприцевой насос, который доставил каплю жидкости ∼30– мкм л в сосуд для жидкости рядом с ней. Ответы на неактивный рычаг регистрировались, но не имели запрограммированных последствий. Световой стимул (домашний свет) был установлен над левым и правым рычагами реакции камеры самоуправления. IBM-совместимый компьютер контролировал подачу жидкостей, представление стимулов и запись данных.
Фаза кондиционирования и прекращения самостоятельного приема алкоголя.
Все сеансы обучения животных и тестирования проводились во время темной фазы их цикла свет / темнота. Животных обучали самостоятельно вводить 10% (об. / Об.) Этанола или воды в ежедневных 30-минутных сеансах с использованием графика фиксированного соотношения 1 (FR 1). Целью фазы кондиционирования было научить животных различать наличие этанола (усиление) и воды (отсутствие усиления). Дискриминационные стимулы, предсказывающие доступность этанола или воды, предъявлялись во время каждого сеанса самостоятельного введения этанола или воды.Экстракт апельсинового аромата служил контекстным стимулом (S +) для этанола, тогда как о доступности воды сигнализировал экстракт лимонной травы (S–). Эти обонятельные стимулы генерировались путем внесения нескольких капель соответствующего экстракта в подстилку оперантной камеры перед каждым сеансом. Кроме того, каждое нажатие на рычаг, приводящее к доставке этанола, сопровождалось 5-секундным условным стимулом мигающим светом (CS +), тогда как 5-секундный стимул постоянного света (CS–) предъявлялся с доставкой воды.Пятисекундный период служил «тайм-аутом», в течение которого ответы записывались, но не подкреплялись. В конце каждого сеанса подстилку камеры меняли и подносы тщательно очищали. В течение первых 2 дней кондиционирования животных содержали без жидкости в течение 20 часов в день. Затем проводили сеансы алкоголя и воды без лишения жидкости случайным образом до тех пор, пока животные не получили в общей сложности 10 сеансов алкоголя и 10 водных сеансов.
После завершения фазы кондиционирования крыс подвергали ежедневным 30-минутным сеансам вымирания в течение 5 последовательных дней, что в целом было достаточно для достижения пониженной скорости реакции, приближающейся к критерию вымирания, равному 20% от последних сеансов кондиционирования.Сеансы угасания начинались с вытягивания рычагов без предъявления обонятельных дискриминирующих стимулов. Ответы на ранее активный рычаг активировали шприцевой насос, не приводя к доставке спирта или воды или предъявлению зависимых от ответа сигналов (мигающий или постоянный свет стимула).
Восстановление, вызванное этанолом (фармакологические исследования II).
Испытания по восстановлению начались через 3 дня после последнего сеанса вымирания. В этих тестах крысы подвергались воздействию тех же условий, что и во время фазы кондиционирования, за исключением того, что жидкости (этанол или вода) не были доступны.Сеансы инициировались вытягиванием обоих рычагов и предъявлением дискриминационных стимулов, связанных с этанолом (S +) или водой (S–). Ответы на активный рычаг сопровождались активацией шприцевого насоса и представлением CS + в состоянии S + или CS– в состоянии S–. Половину животных тестировали в условиях S + / CS + на 1 день и в условиях S– / CS– на 2 день. Условия для другой половины животных были обратными. Количество ответов на оба активных (т.например, связанный с этанолом рычаг для состояния S + / CS + и связанный с водой рычаг для состояния S– / CS–) и неактивный рычаг (т. е. связанный с водой рычаг для состояния S + / CS + и связанный с этанолом рычаг для S– / CS. — состояние) регистрировали на протяжении всего эксперимента.
Чтобы проверить влияние SSR504734 и A-1246399 на возобновление поиска алкоголя, вызванное сигналом, животные были разделены на четыре группы на основе их показателей во время последних сеансов кондиционирования и угасания ( n = 8–9 на группа).Для теста две группы получали либо 10 мг / кг SSR504734, либо 30 мг / кг A-1246399, тогда как двум другим группам животных вводили соответствующий носитель. Введение препарата производилось за 30 минут до процедуры восстановления. Обратите внимание, что более низкие дозы этих соединений не были эффективны в предыдущем тесте ADE и были исключены из дальнейшего исследования, а контракт на исследование RO4993850 не охватывал тест восстановления, вызванного сигналом.
Возобновление поиска кокаина, вызванное сигналом
Чтобы проверить, влияет ли блокада GlyT1 только на эффекты, связанные с алкоголем, или влияют ли также долгосрочные эффекты других наркотиков, вызывающих злоупотребление, таких как кокаин, отдельная группа крыс в модель восстановления, вызванного кокаиновым сигналом.
Имплантация катетера.
Крыс анестезировали 4% изофлураном и поддерживали анестезию ~ 2% изофлурана в течение всей операции. Катетер, состоящий из микроренатановой трубки (внутренний диаметр: 0,58 мм, внешний диаметр: 0,94 мм; Bilaney Consultants, Дюссельдорф, Германия), был имплантирован в правую яремную вену, а совместимый задний держатель (Bilaney Consultants) был введен под кожа и выступала в срединно-лопаточной области. Крысам давали выздоровление за 9–14 дней до начала сеансов самостоятельного введения кокаина.Катетеры ежедневно промывали гепаринизированным раствором (100 МЕ / мл), содержащим 1 мг / мл энрофлоксацина (Baytril, Bayer HealthCare LLC Animal Health Division, Shawnee Mission, KS).
Аппарат самоуправления.
Возобновление поиска кокаина, вызванное сигналом, проводилось в операционных камерах (Imetronic, Пессак, Франция), заключенных в вентилируемые звукоизолирующие кабины. На противоположных стенках камер на высоте 5 см над полом решетки располагались два носовых отверстия. Ответы на «тыканье носом» регистрировались путем прерывания светового луча, проецируемого через отверстие.Прокалывание одной (активной) лунки привело к доставке 40 мкл л (0,26 мг) кокаина в течение 2 секунд. Было зафиксировано протыкание в другом (неактивном) отверстии, но не имело никаких запрограммированных последствий. Белая сигнальная лампа была расположена на 9,5 см выше активного отверстия, а синяя сигнальная лампа находилась на противоположной стене на 33 см выше пола решетки. Звуковой генератор («гудок», 3 кГц, 60 дБ) располагался на задней стене в 40 см от пола решетки. Эксперименты контролировались, а данные собирались с помощью Windows-совместимого программного обеспечения SK_AA.
Фаза кондиционирования и прекращения самостоятельного приема кокаина.
Все сеансы обучения животных и тестирования проводились во время темной фазы их цикла свет / темнота. Крыс обучали самостоятельному введению кокаина за 2-часовые ежедневные сеансы, пока не установился стабильный ответ. В течение первых трех сеансов животные обучались по графику подкрепления с фиксированным соотношением 3 (FR 3). График FR 5 использовался для оставшейся части фазы кондиционирования. О наличии кокаина сигнализировал синий световой сигнал, который постоянно горел во время сеанса.После необходимого количества ударов носом в активную лунку загорелась белая сигнальная лампочка и раздался звуковой сигнал; через 1 секунду был активирован инфузионный насос. Белый световой сигнал и звуковой сигнал поддерживались в течение 4 секунд в общей сложности. Вливание кокаина сопровождалось 40-секундным перерывом, в течение которого тыкание носом в любое отверстие не имело запланированных последствий.
После завершения фазы кондиционирования крысы проходили обучение вымиранию, во время которого тыкание носом не приводило ни к введению кокаина, ни к предъявлению сигналов, зависящих от ответа.Вымирание длилось 17 сеансов, что в сумме было достаточно для достижения пониженной скорости реакции, приближающейся к критерию вымирания, равному 20% от последних сеансов кондиционирования.
Восстановление, вызванное кокаином (фармакологические исследования III).
Тестирование восстановления было проведено на следующий день после последнего сеанса исчезновения. В этом тесте крысы подвергались воздействию тех же условий, что и во время фазы кондиционирования, за исключением того, что кокаин не был доступен. Камера оперантов постоянно освещалась синим сигнальным светом, который служил контекстным стимулом для доступности кокаина.После необходимого количества ударов носом в активную лунку загорелась белая сигнальная лампочка и раздался звуковой сигнал; через 1 секунду был активирован инфузионный насос. Количество тыкований носом как в активных, так и в неактивных отверстиях регистрировалось на протяжении всего теста.
Чтобы проверить влияние SSR504734 и A-1246399 на возобновление поиска кокаина, вызванное сигналом, животные были разделены на две группы на основе их показателей во время последних сеансов кондиционирования и угасания ( n = 6-8 на группа).Для теста одной группе животных вводили носитель, а другой — 10 мг / кг SSR504734. Через две недели крыс распределили в новые экспериментальные группы в соответствии с планом латинского квадрата. Одной группе крыс вводили 30 мг / кг A-1246399, а другой группе вводили соответствующий носитель. Введение препарата производилось за 30 минут до повторного теста.
Статистический анализ
Данные, полученные на основе измерений ADE (общее потребление алкоголя, потребление воды) и двигательной активности, анализировали с использованием двухфакторного дисперсионного анализа (ANOVA) с повторными измерениями (факторы: группа лечения и день).Данные двигательной активности анализировали с использованием записи 12-часовых интервалов после инъекции, которые соответствовали активной фазе животных. Анализ данных о влиянии лечения на изменение массы тела крысы проводили с использованием однофакторного дисперсионного анализа (фактор: группа лечения). Данные, полученные в результате экспериментов по поиску алкоголя и кокаина, были проанализированы с помощью трехфакторного дисперсионного анализа ANOVA с повторными измерениями [факторы были: группа лечения, реакции рычага / толчки носа (активные vs.неактивный) и сеанс (исчезновение vs. восстановление)]. При обнаружении значительных различий проводились апостериорные тесты Стьюдента Ньюмана-Кеулса. Выбранный уровень значимости P <0,05.
Результаты
SSR504734, A-1246399 и RO4993850 Уменьшение количества алкогольных напитков, подобных рецидивам.
Во время тестирования препарата повторное употребление алкоголя в группах животных, получавших носитель, не отличалось от предыдущих фаз депривации (данные не показаны). Двусторонний дисперсионный анализ ANOVA с повторными измерениями продемонстрировал общее увеличение потребления алкоголя после фазы депривации по сравнению с исходным уровнем потребления алкоголя [фактор-день: F (6,168) = 69.3, P <0,0001; F (6162) = 53,3, P <0,0001 и F (6168) = 8,2, P <0,0001 для групп обработки SSR504734, A-1246399 и RO4993850 соответственно]. Кроме того, анализ данных показал, что все три вида лечения - SSR504734, A-1246399 и RO4993850 - значительно снижали потребление алкоголя в дни после абстиненции по сравнению с таковым у животных, получавших носитель [факторная группа лечения × дневной эффект взаимодействия: F (18 168) = 3,1 , P <0,0001; F (18,162) = 3.3, P <0,0001 и F (18 168) = 4,3, P <0,0001 для групп обработки SSR504734, A-1246399 и RO4993850 соответственно]. Снижение потребления этанола крысами, получавшими один из блокаторов GlyT1, было непродолжительным и могло быть измерено только в течение первой недели после абстиненции (данные о потреблении алкоголя в течение последующих недель после абстиненции не приводятся). Следующий апостериорный тест показал, что лечение крыс наивысшей дозой (10 мг / кг) SSR504734 устраняет рецидивирующее потребление алкоголя (рис.1А). Самая высокая доза (30 мг / кг) лечения A-1246399 не устраняла полностью ADE. Потребление этанола было увеличено в первый день после абстиненции по сравнению с исходным уровнем потребления (рис. 1С). Ежедневное введение любой дозы RO4993850 полностью устраняет ADE, а самая высокая доза (30 мг / кг) RO4993850 снижает потребление алкоголя ниже исходного уровня (рис. 1E). Потребление воды крысами, получавшими SSR504734-, A-1246399- и RO4993850, было увеличено / имело тенденцию к увеличению в дни лечения по сравнению с таковым у животных, получавших носитель [группа обработки фактора × дневной эффект взаимодействия: P = 0.29; F (18,162) = 2,7, P <0,001 и P = 0,06 для групп лечения SSR504734, A-1246399 и RO4993850 соответственно], демонстрируя, что все виды лечения избирательно влияли на потребление алкоголя (рис. 1, B, D, и F).
Рис. 1.Потребление общего этанола, рассчитанного в граммах чистого спирта на килограмм веса тела в день (A, C и E), и воды, рассчитанного в миллилитрах на килограмм веса тела в день (B, D и F), в (A и B) носителе, 1 мг / кг SSR504734, 3 мг / кг SSR504734 и 10 мг / кг SSR504734 ( n = 8 на условия обработки), (C и D) носитель, 3 мг / кг A-1246399, 10 мг / кг A-1246399 и 30 мг / кг A-1246399 ( n = 7-8 на условия лечения) и (E и F) носитель, 3 мг / кг RO4993850, 10 мг / кг RO4993850 и 30 мг / кг RO4993850 ( n = 8 на условия лечения) до и после периода депривации в течение 3 недель.Измерения потребления этанола за последнюю неделю приведены в качестве базового уровня потребления алкоголя (BL). Животные получали в общей сложности пять, каждые 12 часов, i.p. инъекции SSR504734 и A-1246399 и три пероральных введения RO4993850 один раз в день (вертикальные стрелки). Бутылки с этанолом были повторно введены после второго введения лекарства. Данные представлены в виде средних значений ± S.E.M. * Значимые отличия от контрольной группы носителя, P <0,05.
Анализ данных о двигательной активности выявил общее снижение активности в домашней клетке во всех группах животных, что, вероятно, было вызвано алкогольной интоксикацией в течение первых дней после абстиненции [факторный день: F (6,168) = 38.7, P <0,0001; F (6,162) = 69,7, P <0,0001 и F (6,168) = 48,2, P <0,0001 для групп обработки SSR504734, A-1246399 и RO4993850 соответственно] (рис. 2). Двусторонний дисперсионный анализ ANOVA с повторными измерениями показал, что обработка SSR504734 не оказывала значительного влияния на двигательную активность животных (эффект взаимодействия группы факторов и группы обработки × день: P = 0,19 и P = 0,14, соответственно) (рис. . 2А). Однако самая высокая доза A-1246399 значительно снижала активность животных в домашней клетке [группа обработки фактора × эффект взаимодействия в день: F (6,162) = 5.1, P <0,0001] (фиг. 2B). RO4993850 также снижает двигательную активность; однако это изменение активности было значимым только после первого введения лекарственного средства [группа обработки фактора и группа лечения × дневной эффект взаимодействия: F (3,28) = 3,3, P <0,05 и P = 0,08, соответственно] (рис. . 2С).
Рис. 2.Двигательная активность в (A) носителе, 1 мг / кг SSR504734, 3 мг / кг SSR504734 и 10 мг / кг SSR504734 ( n = 8 на условия лечения), (B) носитель, 3 мг / кг A-1246399, 10 мг / кг A-1246399 и 30 мг / кг A-1246399 ( n = 7-8 на условия лечения) и (C) носитель, 3 мг / кг группы животных, получавших RO4993850, 10 мг / кг RO4993850 и 30 мг / кг RO4993850 ( n = 8 на условия лечения).. » Стрелка указывает на повторное воздействие растворов этанола на животных. Данные представлены в виде средних значений ± S.E.M. * Значимые отличия от контрольной группы носителя, P <0,05.
Обработка крыс наивысшей дозой SSR504734 или A-1246399 вызвала небольшую (~ 1%), но значительную потерю массы тела [группа обработки фактора: F (3,28) = 3,5, P <0,05 и F. (3,27) = 27,1, P <0,0001]. Масса тела животного существенно не изменилась во время лечения RO4993850, что указывает на отсутствие влияния на прием пищи и / или метаболизм (группа обработки фактора: P = 0.19).
SSR504734 и A-1246399 Сокращенное восстановление поведения, связанного с поиском алкоголя.
Крысы достигли 90 ± 6 реакций рычага, связанных с этанолом (состояние S + / CS +) и 22 ± 2 ответов рычага, связанных с водой (состояние S– / CS–) к концу фазы кондиционирования. Давление на рычаг постепенно уменьшалось в течение пяти сеансов угасания до 9,3 ± 0,6 и 7,1 ± 0,4 для ранее усиленного этанолом и усиленного водой рычага, соответственно.
Трехфакторный дисперсионный анализ ANOVA с повторными измерениями показал, что количество нажатий на рычаг увеличивалось во время теста восстановления, вызванного этанолом, по сравнению с таковым во время последних сеансов угасания [факторный сеанс: F (1,28) = 14.0, P <0,001 и F (1,30) = 11,6, P <0,01 для групп обработки SSR504734 и A-1246399 соответственно]. В основном это было вызвано повышенной реакцией на активный рычаг в группе крыс, обработанных транспортным средством. Значительное снижение реакции рычага во время теста восстановления S + / CS + наблюдалось у 10 мг / кг крыс, получавших SSR504734 [группа обработки фактора × взаимодействие сеанса: F (1,28) = 4,6, P <0,05] (рис. 3А). Дальнейший апостериорный анализ показал, что реакция на активный рычаг в группе, получавшей SSR504734, во время теста восстановления S + / CS + не отличалась от последних сеансов исчезновения.Реакции неактивного рычага существенно не различались между группами крыс, получавших носитель и SSR504734 (фиг. 3B). Введение 30 мг / кг A-1246399 имело аналогичный эффект на вызванное сигналом восстановление поведения, связанного с поиском этанола. У крыс, получавших A-1246399, реакция на активный рычаг во время теста восстановления не отличалась от реакции, наблюдаемой во время сеансов угасания [группа обработки фактора × взаимодействие сеанса: F (1,30) = 9,0, P <0,01] (Рис. 4A). Реакция на неактивный рычаг существенно не изменилась (рис.4Б).
Рис. 3.Влияние носителя и 10 мг / кг SSR504734 ( n = 8 на условия лечения) на этанол (A и B) и воду (C и D), вызванное сигналом восстановления. Данные представлены как среднее количество нажатий на активные (A и C) и неактивные (B и D) рычаги во время последних сеансов угасания и как количество ответов после предъявления стимулов, ранее сопряженных либо с этанолом, либо с водой. Данные представлены в виде средних значений ± S.E.M. * Значительные различия в ответах рычага угасания; + Значительные отличия от контрольной группы носителя, P <0.05.
Рис. 4.Влияние носителя и 30 мг / кг A-1246399 ( n = от 8 до 9 на условия лечения) на этанол (A и B) и воду (C и D), вызванные меткой. восстановление. Данные представлены как среднее количество нажатий на активные (A и C) и неактивные (B и D) рычаги во время последних сеансов угасания и как количество ответов после предъявления стимулов, ранее сопряженных либо с этанолом, либо с водой. Данные представлены в виде средних значений ± S.E.M. * указывает на существенные различия в ответах рычага угасания; + указывает на значительные отличия от контрольной группы носителя, P <0.05.
Интересно, что животные также увеличили количество ответов во время теста S– / CS– по сравнению с последними сеансами угасания [факторный сеанс: F (1,28) = 7,0, P <0,05 и F (1,30 ) = 11,2, P <0,01 для групп обработки SSR504734 и A-1246399 соответственно]. Это увеличение не было избирательным по отношению к рычагу (активный против неактивного), что указывает на то, что оно могло быть вызвано общим возбуждением животного [фактор сеанса × взаимодействие рычага: P = 0.29 и P = 0,12 для групп обработки SSR504734 и A-1246399 соответственно]. Оба вида лечения значительно снижали реакцию на связанный с водой рычаг [факторная группа лечения × взаимодействие сеанса: F (1,28) = 8,2, P <0,01 и F (1,30) = 8,9, P <0,01 для SSR504734 и группы обработки A-1246399, соответственно]; снижение реакции на неактивный рычаг не достигло статистической значимости (рис. 3, C и D; рис. 4, C и D).
SSR504734 и A-1246399 Сокращенное восстановление кокаинового поведения, вызванное стимулом.
Крысы достигли 660 ± 184 ударов носом, связанных с кокаином, и 51 ± 12 неактивных укусов носом к концу фазы кондиционирования. Количество тыкающих носом постепенно уменьшалось в течение 17 сеансов вымирания до 42 ± 4 ранее усиленных кокаином тычков в нос и до 30 ± 6 неусиленных толчков в нос.
Трехфакторный дисперсионный анализ с повторными измерениями показал, что количество тыкающих носом увеличилось во время теста восстановления, вызванного кокаиновым сигналом, по сравнению с таковым во время последних сеансов исчезновения [факторный сеанс: F (1,26) = 33.3, P <0,0001 и F (1,22) = 19,0, P <0,001 для групп обработки SSR504734 и A-1246399 соответственно]. В основном это было вызвано повышенной реакцией на активное протыкание носа в группе крыс, получавших носитель. Введение 10 мг / кг SSR504734 отменило поиск кокаина. Количество активных тыков носом во время тестирования восстановления у животных, получавших SSR504734, не отличалось от фазы исчезновения [факторная группа лечения × взаимодействие сеанса: F (1,26) = 12.1, P <0,01] (фиг. 5A). Точно так же активные тычки носом во время теста восстановления были отменены лечением A-1246399 [факторная группа лечения × взаимодействие сеанса: F (1,22) = 16,2, P <0,001] (фиг. 6A). Неактивные тычки в нос не уменьшались ни обработкой SSR504734, ни обработкой A-1246399 по сравнению с группой, обработанной носителем (фиг. 5B; фиг. 6B).
Рис. 5.Влияние носителя и 10 мг / кг SSR504734 ( n = 7–8 на условия лечения) на восстановление, вызванное кокаиновым сигналом.Данные представлены как среднее количество активных (A) и неактивных (B) тыкающих носом в течение последних сеансов угасания и как количество тыковок после предъявления стимулов, ранее сопряженных с кокаином. Данные представлены в виде средних значений ± S.E.M. * Значительные отличия от вымирания носа-тыкают; + Значимые отличия от контрольной группы носителя, P <0,05.
Рис. 6.Влияние носителя и 30 мг / кг A-1246399 ( n = 6–7 на условия лечения) на восстановление, вызванное кокаиновым сигналом.Данные представлены как среднее количество активных (A) и неактивных (B) тыкающих носом в течение последних сеансов угасания и как количество тыковок после предъявления стимулов, ранее сопряженных с кокаином. Данные представлены в виде средних значений ± S.E.M. * Значительные отличия от вымирания носа-тыкают; + Значимые отличия от контрольной группы носителя, P <0,05.
Обсуждение
Наше исследование продемонстрировало, что три различных блокатора GlyT1 снижали или даже отменяли рецидивирующее потребление алкоголя у самцов крыс в течение первых дней после абстиненции в условиях свободного выбора из четырех бутылочек.Повторное введение SSR504734 или RO4993850 отменило повторное употребление алкоголя, и значительное снижение было измерено после лечения A-1246399. Чтобы компенсировать снижение потребления этанола, потребление воды увеличивалось у крыс, получавших лекарственное средство, по сравнению с крысами, получавшими носитель, что демонстрирует селективность тестируемых соединений в отношении потребления алкоголя. Блокада GlyT1 вызвала некоторую гиполокацию в случае лечения A-1246399 и RO4993850, но никаких других побочных эффектов не наблюдалось.Вызванное сигналом восстановление поведения, связанного с поиском алкоголя, было отменено введением блокаторов GlyT1 SSR504734 и A-1246399. Оба соединения также снижали вызванное сигналом возобновление поиска воды и поиска кокаина, предполагая более общее участие глицинергической системы в ассоциации стимула и вознаграждения. Неусиленная реакция не претерпевала значительных изменений во время всех трех тестов восстановления, что исключает возможность того, что снижение ответа во время тестов восстановления было вызвано неселективным влиянием лечения на поведение нажатия рычага.
В нашем более раннем исследовании мы показали, что повторное лечение блокатором GlyT1 Org-25935 уменьшало вероятность рецидива алкоголя у хронически пьющих крыс, что могло быть результатом отмены вызванных алкоголем изменений глицинергической и глутаматергической передачи сигналов. Следовательно, были обнаружены множественные изменения в профилях экспрессии генов, связанных с глицинергической и глутаматергической передачей сигналов. Введение блокатора Glyt1 обращало измененные уровни экспрессии этих генов к уровням у крыс, не употреблявших алкоголь.Поскольку и GlyRs, и NMDARs являются первичными мишенями для алкоголя, было высказано предположение, что поведенческие эффекты Org-25935, вероятно, опосредуются через любой из этих рецепторов (Vengeliene et al., 2010). В настоящем исследовании мы использовали три блокатора GlyT1 с несвязанными основными химическими структурами, предоставленные тремя разными поставщиками. Во всех трех экспериментах лечение привело к значительному снижению потребления этанола во время рецидивирующей ситуации, таким образом подтверждая, что глицинергическая система действительно вовлечена в рецидивное употребление алкоголя.
В нашем исследовании блокада GlyT1 уменьшала не только алкоголь, но и реакцию воды, вызванную сигналом. Это может указывать на более общий эффект блокады GlyT1 на обучение стимулом-вознаграждением, и действительно, вызванное сигналом восстановление поведения, связанного с поиском кокаина, также было уменьшено SSR504734 и A-1246399. Также было показано, что лечение SSR504734 снижает потребность в никотине (Cervo et al., 2013). Однако исследование Cervo et al. (2013) продемонстрировали, что на восстановление поиска сахарозы, вызванное сигналом, не повлияло введение SSR504734, предполагая, что блокада GlyT1 избирательна в отношении стимулов, связанных с лекарствами.В нашем исследовании этанол и кондиционирование воды проводились на одном и том же животном; следовательно, у этих животных мог иметь место некоторый уровень генерализации реплик. Это могло повлиять на воздействие лечения на поведение животных во время теста на восстановление воды. Опять же, мы не знаем о вкладе GlyR и / или NMDAR в действие блокаторов GlyT1. Однако критическая роль NMDAR в опосредовании реакции поиска наркотиков, вызванной сигналом, была обозначена в многочисленных исследованиях, посвященных другим наркотикам, вызывающим злоупотребление, в первую очередь кокаину (Bäckström and Hyytiä, 2006, 2007; Feltenstein and See, 2007; Engblom et al., 2008; Mameli et al., 2009), что предполагает вклад NMDAR в эффекты блокады GlyT1. Известно, что высвобождение глутамата повышается во время восстановления поведения, связанного с поиском наркотиков, у животных, обученных самостоятельно принимать алкоголь (Gass et al., 2011), кокаин (McFarland et al., 2003) или героин (LaLumiere and Каливас, 2008). Таким образом, предполагается, что снижение глутаматергической активности во время воздействия условной реплики может снизить склонность к поиску лекарств.Однако ни конкурентные антагонисты NMDAR, ни блокаторы каналов NMDAR не оказались эффективными при восстановлении, вызванном алкоголем или кокаином (Bespalov et al., 2000; Bäckström and Hyytiä, 2004, 2006; Bachteler et al., 2005). Блокирование коагонистического сайта связывания глицина NMDAR, по-видимому, является единственным эффективным способом нацеливания на этот рецептор для снижения поведения, вызванного поиском лекарств, вызванного сигналом (Bäckström and Hyytiä, 2004, 2006). Было показано, что повышенные уровни внеклеточного глицина усиливают функцию NMDAR, но также вызывают интернализацию рецепторов AMPA и NMDA и вызывают долгосрочное подавление возбуждающих постсинаптических токов (Nong et al., 2003; Чен и др., 2011; Alberati et al., 2012), которые, в свою очередь, могли быть ответственны за снижение продуктивности животных во время тестов восстановления, вызванных сигналом. Накопление внеклеточного глицина ведет к тонической активации GlyRs, которая является частью гомеостатического механизма преодоления гипервозбудимости нейронов (Zhang et al., 2008). На сегодняшний день высвобождение глицина не измерялось во время тестирования восстановления, вызванного сигналом; однако было показано, что GlyR модулируют активность мезокортиколимбической дофаминергической системы (Jonsson et al., 2014; Clarke et al., 2015), который играет ключевую роль в ответах на поиск наркотиков (Ito et al., 2000). Существенная роль GlyRs в связанном с алкоголем поведении также была продемонстрирована с использованием мышей с мутантной субъединицей GlyR (Blednov et al., 2015).
Все три протестированных препарата имели очень низкий профиль побочных эффектов. Некоторая гиполокация наблюдалась во время тестирования на наркотики в модели ADE; тем не менее, общее угнетающее действие на центральную нервную систему можно исключить, так как не было затронуто реакция неактивного рычага в тесте восстановления.Гиполокация, вызванная лекарственным лечением, во время тестирования ADE, вероятно, была связана с активацией ингибирующего GlyR. Этот эффект отсутствовал в случае SSR504734 и кратковременный в случае RO4993850. Потеря массы тела, вызванная медикаментозным лечением, была очень незначительной или отсутствовала. Эти результаты показывают, что риск серьезных побочных эффектов, вызванных блокадой GlyT1, очень низок. В согласии с этим, введение блокаторов GlyT1 здоровым добровольцам и пациентам, страдающим алкоголем, продемонстрировало, что эти соединения обычно хорошо переносятся (de Bejczy et al., 2014; Hofmann et al., 2016).
В заключение, наши результаты показали, что блокада GlyT1 нарушает стремление к алкоголю и рецидивирующее поведение у крыс. Это лечение также повлияло на реакцию, вызванную кокаином. В более ранних исследованиях сообщалось, что восстановление поиска никотина, вызванное сигналом, также уменьшалось за счет блокады GlyT1 (Cervo et al., 2013). Механизм такого общего эффекта блокады GlyT1 на ответную реакцию, вызванную лекарственными сигналами, в настоящее время неясен. Мы предлагаем GlyT1 в качестве новой фармакологической мишени для предотвращения рецидивов.Однако необходимы дальнейшие исследования, чтобы лучше понять механизм действия блокаторов GlyT1.
Благодарности
Мы хотели бы поблагодарить Сабрину Кох и Елену Бюхлер за отличную техническую помощь.
Авторский вклад
Участвовал в разработке исследования: Vengeliene, Alberati, Behl, Bespalov, Spanagel.
Проведенные эксперименты: Венгелиене, Россманит, Такахаши.
Внесены новые соединения : Альберати, Бел, Беспалов.
Проведенный анализ данных: Vengeliene.
Написал или участвовал в написании рукописи: Vengeliene, Spanagel.
Сноски
- Получено 28 августа 2017 г.
- Принято 2 января 2018 г.
Финансовую поддержку для этой работы оказал Bundesministerium für Bildung und Forschung [e: Med program, FKZ: 01ZX1311A ( et al., 2013)]. Р.С. раскрыл, что получил исследовательский грант от Ф.Hoffmann-La Roche Ltd об изучении влияния RO4993850 на эффект лишения алкоголя. Все остальные авторы не сообщили о финансовых интересах или потенциальных конфликтах интересов.
https // doi.org / 10.1124 / jpet.117.244822.
Сокращения
- A-1246399
- N- (2 — (((7S, 8R) -7- (азетидин-1-ил) -8-бензил-5,6,7,8-тетрагидронафталин- 2-ил) окси) этил) -1-метил-1H-имидазол-4-сульфонамид
- ADE
- эффект лишения спирта
- AMPA
- α -амино-3-гидрокси-5-метил-4-изоксазолпропионовый кислота
- ANOVA
- дисперсионный анализ
- CS
- условный стимул
- FR
- фиксированное соотношение
- GlyR
- рецептор глицина
- GlyT1
- переносчик глицина408 -аспартатные рецепторы
- Org-25935
- 2 — ([(1 R , 2 S ) -6-метокси-1-фенил-1,2,3,4-тетрагидронафталин-2-ил] метил- метиламино) уксусная кислота
- RO4993850
- [5-метилсульфонил-2 — [(1S) -2,2,2-трифтор-1-метилэтокси] фенил] — [3- (трифторметил) -5,7-дигидропирроло [3,4-b] пиридин -6-ил] метанон
- S
- стимул
- SSR504734
- 2-хлор- N — [( S ) -фенил — [(2S) -пиперидин-2-ил] метил] -3- (трифторметил) бензамид
- © Американское общество фармакологии и экспериментальной терапии, 2018 г.
- Принята к печати 19 сентября 2014 г.
A.N. и К. внес равный вклад в эту работу.
Эта работа была полностью или частично поддержана Национальным институтом здравоохранения Национального института злоупотребления алкоголем и алкоголизмом [гранты R01-AA13378, R01-AA13992, K01-AA17243 и R01-AA022448], Американским фондом Фармацевтическое образование и Школа фармацевтики Университета Южной Калифорнии.
Авторы заявляют об отсутствии конфликта интересов.
↵Данная работа выполнена в рамках частичного выполнения требований к соисканию ученой степени доктора философии. степень в области молекулярной фармакологии и токсикологии (A.N.) и степень магистра фармацевтических наук (K.H.M.), Университет Южной Калифорнии.
Naito A, Muchhala K, Asatryan L, Trudell JR, Homanics GE, Davies DL и Alkana RL (2014) Мутации в петле 2 субъединиц рецептора глицина и GABAA создают сверхчувствительные рецепторы этанола (USER) с потенциальным применением как инструменты картирования мозга. Исследовательская конференция Гордона, Алкоголь и нервная система ; 16–21 февраля 2014 г .; Галвестон, Техас.
Naito A, Muchhala K, Asatryan L, Trudell JR, Homanics GE, Davies DL и Alkana RL (2014) Мутации петли 2 субъединиц рецептора глицина и GABAA создают сверхчувствительные рецепторы этанола (USER) с потенциальным применением в картирование мозга. 37-е Ежегодное собрание Общества исследователей алкоголизма и 17-й Конгресс ISBRA ; 21–25 июня 2014 г .; Бельвью, Вашингтон.
Naito A, Muchhala K, Asatryan L, Homanics GE, Trudell JR, Perkins DI, Davies DL и Alkana RL (2013) Разработка сверхчувствительных рецепторов этанола (ПОЛЬЗОВАТЕЛИ) в качестве новых инструментов для исследования алкоголя. Ежегодное собрание Общества неврологии ; 9–13 ноября 2013 г .; Сан-Диего, Калифорния.
Эта работа ранее была представлена на следующих встречах в виде рефератов: Muchhala K, Naito A, Asatryan L, Homanics GE, Trudell JR, Perkins DI, Davies DL и Alkana RL (2013) Разработка сверхчувствительного этанола рецепторы (ПОЛЬЗОВАТЕЛИ) как новые инструменты для исследования алкоголя. Движущиеся мишени: нейродегенеративные заболевания ; 23 августа 2013 г .; Лос Анджелес, Калифорния.
Naito A, Muchhala K, Asatryan L, Trudell JR, Homanics GE, Davies DL, Alkana RL и Perkins DI (2013) Мутации в петле 2 рецептора GABAA создают сверхчувствительные рецепторы этанола (USER), чувствительные к микромолярным концентрациям этанола. 36-е ежегодное собрание Общества исследователей алкоголизма, алкоголя: Clin Exp Res 37: P199; 22–26 июня 2013 г .; Орландо, Флорида.
Naito A, Muchhala K, Asatryan L, Davies DL, Perkins DI, and Alkana RL (2013) Ночные рецепторы: развитие сверхчувствительных рецепторов этанола (USER) в рецепторах глицина для выяснения молекулярного механизма алкогольная зависимость. USC Graduate Research Symposium ; 2 апреля 2013 г .; Лос Анджелес, Калифорния.
Alkana RL, Naito A, Muchhala K, Asatryan L, Li K, Homanics GE, Trudell JR, Davies DL и Perkins DI (2012) Разработка мутантных GlyR петли 2, чувствительных к микромолярным концентрациям этанола. Ежегодное собрание Общества неврологии ; 13–17 октября 2012 г .; Новый Орлеан, Луизиана.
Alkana RL, Trudell JR, Asatryan L, Naito A, Muchhala K, Li K, Davies DL и Perkins DI (2012) Функциональное понимание механизмов действия этанола в GlyR и GABAAR, выявленных путем манипулирования петлей внеклеточного домена 2. Международное общество биомедицинских исследований алкоголизма. Международный Конгресс . 9–12 сентября 2012 г .; Саппоро, Япония.
Alkana RL, Perkins DP, Li K, Homanics GE, Naito A, Trudell JR и Davies DL (2012) Последовательность петли 2 глубоко влияет на чувствительность альфа-1-глициновых рецепторов (GlyR) к этанолу. Ежегодное собрание исследовательского общества по алкоголизму ; 23–27 июня 2012 г .; Сан-Франциско, Калифорния.
dx.doi.org/10.1124/mol.114.093773.
Дополнительные материалы к этой статье доступны на сайте molpharm.aspetjournals.org.
Переносчики глицина как новые терапевтические мишени при шизофрении, алкогольной зависимости и боли
Avila, A. et al. Активация α2 субъединицы глицинового рецептора способствует миграции корковых интернейронов. Cell Rep. 4 , 738–750 (2013). Эта статья раскрывает новую биологическую роль GlyRα2 в миграции корковых интернейронов во время эмбрионального развития.
Артикул CAS PubMed PubMed Central Google ученый
Волоскер, Регуляция рецептора H. NMDA с помощью D-серина: новые открытия и перспективы. Мол. Neurobiol. 36 , 152–164 (2007).
Артикул CAS PubMed Google ученый
Эйленбург, В., Армсен, В., Бец, Х. и Гомеза, Дж. Транспортеры глицина: основные регуляторы нейротрансмиссии. Trends Biochem. Sci. 30 , 325–333 (2005).
Артикул CAS PubMed Google ученый
Харви, Р. Дж., Топф, М., Харви, К. и Рис, М. И. Генетика гиперэкплексии: больше, чем удивление! Trends Genet. 24 , 439–447 (2008).
Артикул CAS PubMed Google ученый
Supplisson, S.И Roux, M.J. Почему переносчики глицина имеют разную стехиометрию. FEBS Lett. 529 , 93–101 (2002).
Артикул CAS PubMed Google ученый
Обри, К. Р. и др. Транспортеры GlyT2 и VIAAT взаимодействуют для определения везикулярного глицинергического фенотипа. J. Neurosci. 27 , 6273–6281 (2007). Это элегантное исследование демонстрирует, почему функциональные свойства GlyT2 подходят для опосредования высокой цитозольной концентрации глицина, которая необходима для эффективной везикулярной нагрузки с помощью везикулярного ингибиторного переносчика аминокислот в тормозных синапсах.
Артикул CAS PubMed PubMed Central Google ученый
Gomeza, J. et al. Инактивация гена транспортера глицина 1 раскрывает жизненно важную роль поглощения глицина глицина в глицинергическом ингибировании. Нейрон 40 , 785–796 (2003). Это революционное исследование, демонстрирующее, что тяжелый фенотип у мышей с GlyT1-нокаутом, напоминающий глициновую энцефалопатию, связан с гиперактивацией глициновых рецепторов в тормозных глицинергических синапсах.
Артикул CAS PubMed Google ученый
Gomeza, J. et al. Делеция мышиного переносчика глицина 2 приводит к фенотипу гиперэкплексии и постнатальной летальности. Нейрон 40 , 797–806 (2003). Эта важная статья демонстрирует, что GlyT2 жизненно важен для эффективной загрузки медиатора синаптических пузырьков в глицинергических нервных окончаниях, и что мыши с нокаутом GlyT2 являются моделью болезни испуга у человека.
Артикул CAS PubMed Google ученый
Eulenburg, V., Retiounskaia, M., Papadopoulos, T., Gomeza, J. & Betz, H. Функция глиального транспортера глицина 1 важна для раннего постнатального выживания, но не обязательна для взрослых мышей. Glia 58 , 1066–1073 (2010).
Артикул PubMed Google ученый
Руссо, Ф., Обри, К. Р. и Супплиссон, С. Переносчик глицина GlyT2 контролирует динамику наполнения синаптических пузырьков в тормозных нейронах спинного мозга. J. Neurosci. 28 , 9755–9768 (2008).
Артикул CAS PubMed PubMed Central Google ученый
Yee, B. K. et al. Нарушение транспортера глицина 1, ограниченного нейронами переднего мозга, связано с прокогнитивным и антипсихотическим фенотипическим профилем. J. Neurosci. 26 , 3169–3181 (2006). Это первое сообщение о мутантных мышах, лишенных нейрона GlyT1 переднего мозга, ясно демонстрирующее наличие устойчивых к шизофрении фенотипов.
Артикул CAS PubMed PubMed Central Google ученый
Singer, P. et al. Измененные мнемонические функции и устойчивость к антагонизму рецептора N -метил-D-аспартата путем условного нокаута передним мозгом транспортера глицина 1. Неврология 161 , 635–654 (2009).
Артикул CAS PubMed PubMed Central Google ученый
Rees, M. I. et al. Мутации в гене, кодирующем GlyT2 ( SLC6A5 ), определяют пресинаптический компонент вздрагивания у человека. Nature Genet. 38 , 801–806 (2006). Этот отчет демонстрирует, что мутации в человеческом гене GLYT2 вызывают пугающую болезнь с сопутствующими опасными для жизни эпизодами неонатального апноэ.
Артикул CAS PubMed Google ученый
Carta, E. et al. Мутации в гене GlyT2 ( SLC6A5 ) являются второй основной причиной болезни испуга. J. Biol. Chem. 287 , 28975–28985 (2012).
Артикул CAS PubMed PubMed Central Google ученый
Кубелос, Б., Хименес, К. и Зафра, Ф.Локализация переносчика глицина GLYT1 в глутаматергических синапсах головного мозга крысы. Cereb. Cortex 15 , 448–459 (2005).
Артикул PubMed Google ученый
Кубелос, Б., Гонсалес-Гонсалес, И. М., Гименес, С. и Зафра, Ф. Каркасный белок PSD-95 взаимодействует с переносчиком глицина GLYT1 и нарушает его интернализацию. J. Neurochem. 95 , 1047–1058 (2005).
Артикул CAS PubMed Google ученый
Zafra, F. et al. Переносчики глицина по-разному экспрессируются среди клеток ЦНС. J. Neurosci. 15 , 3952–3969 (1995).
Артикул CAS PubMed Google ученый
Койл, Дж. Т., Балу, Д., Бенниворт, М., Басу, А. и Роземан, А. Помимо рецептора дофамина: новые терапевтические мишени для лечения шизофрении. Dialogues Clin. Neurosci. 12 , 359–382 (2010).
PubMed PubMed Central Google ученый
Джавитт, Д. К. Глутамат и шизофрения: фенциклидин, N -метил-D-аспартатные рецепторы и взаимодействия дофамин-глутамат. Внутр. Rev. Neurobiol. 78 , 69–108 (2007). В этой статье прослеживается история возникновения глутаматной гипотезы шизофрении вплоть до недавних попыток увеличить терапию глицином, включая ингибирование обратного захвата глицина.
Артикул CAS PubMed Google ученый
Койл, Дж. Т. рецептор NMDA и шизофрения: краткая история. Schizophr Bull. 38 , 920–926 (2012).
Артикул PubMed PubMed Central Google ученый
Марино, М. Дж., Кнутсен, Л. Дж. И Уильямс, М. Новые возможности для открытия антипсихотических препаратов в постгеномную эпоху. J. Med. Chem. 51 , 1077–1107 (2008).
Артикул CAS PubMed Google ученый
Perry, K. W. et al. Нейрохимические и поведенческие профили селективных ингибиторов GlyT1 ALX5407 и LY2365109 указывают на преимущественное действие в каудальных, а не в корковых областях мозга. Нейрофармакология 55 , 743–754 (2008).
Артикул CAS PubMed Google ученый
Даниш, В.И Парсонс, C.G. Глицин и N -метил-D-аспартат рецепторы: физиологическое значение и возможные терапевтические применения. Pharmacol. Ред. 50 , 597–664 (1998).
CAS PubMed Google ученый
Furukawa, H. & Gouaux, E. Механизмы активации, ингибирования и специфичности: кристаллические структуры лиганд-связывающего ядра рецептора NMDA NR1. EMBO J. 22 , 2873–2885 (2003).
Артикул CAS PubMed PubMed Central Google ученый
Kristensen, A. S. et al. Транспортеры нейромедиаторов SLC6: структура, функция и регуляция. Pharmacol. Ред. 63 , 585–640 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Cioffi, C. L., Liu, S. & Wolf, M. A.Глава 2 — Последние разработки ингибиторов транспортера глицина-1. Annu. Rep. Med. Chem. 45 , 19–35 (2010).
CAS Google ученый
Möhler, H. et al. Переносчик глицина 1 как потенциальная терапевтическая мишень для симптомов, связанных с шизофренией: данные на генетически модифицированных моделях мышей и фармакологическое ингибирование. Biochem. Pharmacol. 81 , 1065–1077 (2011). В этом всестороннем обзоре сравниваются различные модели мышей с нарушением GlyT1. Он также исследует возможность нацеливания на субпопуляции GlyT1 в определенных областях мозга и / или типах клеток.
Артикул CAS PubMed Google ученый
Tsai, G. et al. Нокаут гена переносчика глицина 1: характеристика поведенческого фенотипа. Proc. Natl Acad. Sci. США 101 , 8485–8490 (2004).
Артикул CAS PubMed Google ученый
Martina, M. et al. Снижение экспрессии транспортера глицина типа 1 приводит к серьезным изменениям глутаматергической нейротрансмиссии нейронов гиппокампа CA1 у мышей. J. Physiol. 563 , 777–793 (2005).
Артикул CAS PubMed PubMed Central Google ученый
Балу Д.T. & Coyle, J. T. Состав постсинаптического рецептора глутамата изменен в генетических моделях гипо- и гиперфункции рецептора NMDA у мышей. Brain Res. 1392 , 1–7 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Блэк, М. Д. и др. Прокогнитивная и антипсихотическая эффективность ингибиторов транспорта 1 глицина (GlyT1) в моделях острой и нервной шизофрении: исследования латентного ингибирования на крысах. Psychopharmacol. (Берл.) 202 , 385–396 (2009).
Артикул CAS Google ученый
Boulay, D., Bergis, O., Avenet, P. & Griebel, G. Ингибитор транспортера глицина-1 SSR103800 проявляет селективный и специфический профиль, подобный антипсихотикам, у нормальных и трансгенных мышей. Нейропсихофармакология 35 , 416–427 (2010).
Артикул CAS PubMed Google ученый
Вайнер, И.Нейронные субстраты скрытого торможения: модель переключения. Psychol. Бык. 108 , 442–461 (1990).
Артикул CAS PubMed Google ученый
Мозер, П. К., Хичкок, Дж. М., Листер, С. и Моран, П. М. Фармакология скрытого ингибирования как модель шизофрении на животных. Brain Res. Brain Res. Ред. 33 , 275–307 (2000).
Артикул CAS PubMed Google ученый
Йи, Б.K. et al. Усиление скрытого ингибирования за счет нарушения переносчика глицина 1 опосредуется антидофаминергическим механизмом в прилежащем ядре. Программа № 571.10 / GG15. Планировщик встреч по неврологии, 2010 г. [онлайн], (Сан-Диего, Калифорния; Общество неврологии, 2010).
Google ученый
Грей, Дж. А., Фелдон, Дж., Роулинз, Дж. Н. П., Смит, А. Д. и Хемсли, Д. Р. Нейропсихология шизофрении. Behav.Brain Sci. 14 , 1–19 (1991).
Артикул Google ученый
Harsing, L.G. Jr et al. Ингибиторы транспортера глицина-1 NFPS и Org 24461: фармакологическое исследование. Pharmacol. Biochem. Behav. 74 , 811–825 (2003).
Артикул CAS PubMed Google ученый
Alberati, D. et al. Ингибитор обратного захвата глицина RG1678: фармакологическая характеристика исследуемого агента для лечения шизофрении. Нейрофармакология 62 , 1152–1161 (2012).
Артикул CAS PubMed Google ученый
Depoortere, R. et al. Нейрохимические, электрофизиологические и фармакологические профили селективного ингибитора транспортера глицина-1 SSR504734, потенциально нового типа антипсихотика. Нейропсихофармакология 30 , 1963–1985 (2005).
Артикул CAS PubMed Google ученый
Зингер П., Фелдон, Дж. И Йи, Б. К. Взаимодействие между ингибитором транспортера глицина 1 (GlyT1) SSR504734 и психоактивными препаратами при моторном поведении мышей. Eur. Neuropsychopharmacol. 19 , 571–580 (2009).
Артикул CAS PubMed Google ученый
Leonetti, M. et al. 2-хлор- N — [( S ) -фенил [(2 S ) -пиперидин-2-ил] метил] -3-трифторметилбензамид, моногидрохлорид, ингибитор переносчика глицина типа 1 , увеличивает вызванное высвобождение дофамина в прилежащем ядре крысы in vivo посредством усиленной глутаматергической нейротрансмиссии. Неврология 137 , 555–564 (2006).
Артикул CAS PubMed Google ученый
Лидо, Х. Х., Эриксон, М., Марстон, Х. и Содерпалм, Б. Роль акумбальных рецепторов глицина в модуляции высвобождения дофамина ингибитором транспортера глицина-1 Org25935. Фронт. Психиатрия 2 , 8 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Барч, Д.М. и Картер, С. С. Амфетамин улучшает когнитивные функции у пациентов с шизофренией, принимающих лекарства, и у здоровых добровольцев. Schizophr. Res. 77 , 43–58 (2005).
Артикул PubMed Google ученый
Моландер, А. и Содерпалм, Б. Глициновые рецепторы регулируют высвобождение дофамина в прилежащем ядре крысы. Alcohol Clin. Exp. Res. 29 , 17–26 (2005).
Артикул CAS PubMed Google ученый
Сингх С.П. и Сингх В. Мета-анализ эффективности дополнительных модуляторов рецепторов NMDA при хронической шизофрении. Препараты для ЦНС 25 , 859–885 (2011).
Артикул CAS PubMed Google ученый
Сингер П., Фелдон Дж. И Йи Б. К. Ингибитор транспортера глицина 1 SSR504734 улучшает производительность рабочей памяти в задаче непрерывного отсроченного чередования у мышей C57BL / 6. Psychopharmacol.(Берл.) 202 , 371–384 (2009).
Артикул CAS Google ученый
Робертс Б. М. и др. Ингибирование переносчика глицина устраняет вызванный кетамином дефицит рабочей памяти. Нейроотчет 21 , 390–394 (2010).
Артикул CAS PubMed Google ученый
Boulay, D. et al. Характеристика SSR103800, селективного ингибитора транспортера глицина-1 в моделях, прогнозирующих терапевтическую активность при шизофрении. Pharmacol. Biochem. Behav. 91 , 47–58 (2008).
Артикул CAS PubMed Google ученый
Манахан-Воган, Д., Вильдфорстер, В. и Томсен, С. Спасение гиппокампа LTP и дефицита обучения в модели психоза на крысах путем ингибирования транспортера глицина-1 (GlyT1). Eur. J. Neurosci. 28 , 1342–1350 (2008).
Артикул PubMed Google ученый
Игартуа, И., Solis, J. M. & Bustamante, J. Долгосрочная синаптическая потенциация, индуцированная глицином, опосредуется переносчиком глицина GLYT1. Нейрофармакология 52 , 1586–1595 (2007).
Артикул CAS PubMed Google ученый
Martina, M. et al. Блокада переносчика глицина типа 1 изменяет опосредованные рецептором NMDA ответы и LTP в пирамидных клетках гиппокампа CA1 путем изменения уровней внеклеточного глицина. J. Physiol. 557 , 489–500 (2004).
Артикул CAS PubMed PubMed Central Google ученый
Dubroqua, S. et al. Неповрежденная рабочая память в отсутствие транспортера глицина нейронов переднего мозга 1. Behav. Brain Res. 230 , 208–214 (2012).
Артикул CAS PubMed PubMed Central Google ученый
Зингер П., Boison, D., Mohler, H., Feldon, J. & Yee, B.K. Улучшенная память распознавания после делеции транспортера 1 глицина в нейронах переднего мозга. Behav. Neurosci. 121 , 815–825 (2007).
Артикул PubMed Google ученый
Dubroqua, S., Boison, D., Feldon, J., Mohler, H. & Yee, BK Изучение половой и циркадной зависимости обучающегося фенотипа у мышей с делецией транспортера глицина 1 в двух павловских условиях парадигмы. Neurobiol. Учиться. Mem. 96 , 218–229 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Шимазаки, Т., Каку, А. и Чаки, С. D-серин и ингибитор транспортера глицина-1 улучшают социальную память у крыс. Psychopharmacol. (Берл.) 209 , 263–270 (2010).
Артикул CAS Google ученый
Зингер П., Бойсон, Д., Молер, Х., Фелдон, Дж. И Йи, Б. К. Делеция транспортера глицина 1 (GlyT1) в нейронах переднего мозга способствует обратному обучению: повышенная когнитивная адаптивность? Behav. Neurosci. 123 , 1012–1027 (2009).
Артикул CAS PubMed PubMed Central Google ученый
Nong, Y. et al. Связывание глицина вызывает интернализацию рецептора NMDA. Nature 422 , 302–307 (2003). Это важная статья, демонстрирующая, что стимуляция сайта глицина-B на рецепторах NMDA может стимулировать эти рецепторы для последующей интернализации через клатрин-зависимый эндоцитоз, что подчеркивает возможность того, что глицин и D -серин, которые связываются с сайт глицина-B может также подавлять активность рецептора NMDA.
Артикул CAS PubMed Google ученый
Никифорюк, А.и другие. Блокада транспортера глицина 1 SSR-504734 способствует когнитивной гибкости в зависимости от глицина / рецептора NMDA. Нейрофармакология 61 , 262–267 (2011).
Артикул CAS PubMed Google ученый
Сингер П., Бойсон Д., Молер Х., Фелдон Дж. И Йи Б. К. Модуляция сенсомоторного стробирования при предымпульсном ингибировании с помощью условной делеции транспортера глицина 1 мозга у мышей. Eur. Neuropsychopharmacol. 21 , 401–413 (2011).
Артикул CAS PubMed Google ученый
Kopec, K. et al. Ингибиторы переносчика глицина (GlyT1) с уменьшенным временем пребывания увеличивают предымпульсное торможение, не вызывая гиперлокомоции у мышей DBA / 2. Biochem. Pharmacol. 80 , 1407–1417 (2010).
Артикул CAS PubMed Google ученый
Липина Т., Labrie, V., Weiner, I. & Roder, J. Модуляторы сайта глицина на рецепторах NMDA, D-серин и ALX 5407, проявляют положительные эффекты, аналогичные клозапину, на мышиных моделях шизофрении. Psychopharmacol. (Берл.) 179 , 54–67 (2005).
Артикул CAS Google ученый
Yang, SY, Hong, CJ, Huang, YH & Tsai, SJ Влияние ингибитора транспортера глицина I, N -метилглицина (саркозина), на индуцированные кетамином изменения в сенсомоторном гейтинге и региональном мозге. Экспрессия Fos у крыс. Neurosci. Lett. 469 , 127–130 (2010).
Артикул CAS PubMed Google ученый
Le Pen, G. et al. Дефицит предимпульсного торможения рефлекса испуга у новорожденных крыс с поражением вентрального гиппокампа: обращение глицином и ингибитором переносчика глицина. Biol. Психиатрия 54 , 1162–1170 (2003).
Артикул CAS PubMed Google ученый
Буллич, С.и другие. Биораспределение и дозиметрия излучения лиганда переносчика глицина-1 11 C-GSK931145, определенные на ПЭТ всего тела приматов и человека. Мол. Представь. Биол. 13 , 776–784 (2011).
Артикул Google ученый
Hamill, T. G. et al. Синтез и доклиническая оценка на макаках-резусах [ 18 F] MK-6577 и [ 11 C] CMPyPB переносчик глицина 1 позитронно-эмиссионной томографии радиоиндикаторов. Synapse 65 , 261–270 (2011).
Артикул CAS PubMed Google ученый
Borroni, E. et al. Доклиническая характеристика [ 11 C] R05013853 как нового радиоактивного индикатора для визуализации транспортера глицина типа 1 с помощью позитронно-эмиссионной томографии. Neuroimage 75 , 291–300 (2013).
Артикул CAS PubMed Google ученый
Мезлер, М.и другие. Ингибиторы GlyT1 влияют на транспорт глицина через дискретные сайты связывания. Мол. Pharmacol. 74 , 1705–1715 (2008).
Артикул CAS PubMed Google ученый
Papouin, T. et al. Синаптические и внесинаптические рецепторы NMDA управляются разными эндогенными коагонистами. Ячейка 150 , 633–646 (2012).
Артикул CAS PubMed Google ученый
Лафрен Т.И Левин, Р. Комментарий Управления по санитарному надзору за качеством пищевых продуктов и медикаментов по методологическим вопросам в исследованиях негативных симптомов. Schizophr Bull. 37 , 255–256 (2011).
Артикул PubMed PubMed Central Google ученый
Цай Г. Э. и Лин П. Ю. Стратегии усиления нейротрансмиссии, опосредованной рецептором N -метил-D-аспартата, при шизофрении, критический обзор и метаанализ. Curr.Pharm. Des. 16 , 522–537 (2010).
Артикул CAS PubMed Google ученый
Лин, К. Х., Лейн, Х. Ю. и Цай, Г. Е. Передача сигналов глутамата в патофизиологии и терапии шизофрении. Pharmacol. Biochem. Behav. 100 , 665–677 (2012).
Артикул CAS PubMed Google ученый
Джавитт, Д.C. Ингибиторы транспорта глицина и лечение шизофрении. Biol. Психиатрия 63 , 6–8 (2008).
Артикул CAS PubMed Google ученый
Buchanan, R. W. et al. Когнитивные и негативные симптомы в исследовании шизофрении (CONSIST): эффективность глутаматергических агентов при негативных симптомах и когнитивных нарушениях. Am. J. Psychiatry 164 , 1593–1602 (2007).
Артикул PubMed Google ученый
Heresco-Levy, U. et al. Эффективность высоких доз глицина при лечении стойких негативных симптомов шизофрении. Arch. Общая психиатрия 56 , 29–36 (1999).
Артикул CAS PubMed Google ученый
Lane, H.Y., Chang, Y.C., Liu, Y.C., Chiu, C.C. & Tsai, G.E. Дополнительное лечение саркозином или D-серином при обострении шизофрении: рандомизированное двойное слепое плацебо-контролируемое исследование. Arch. Общая психиатрия 62 , 1196–1204 (2005).
Артикул CAS PubMed Google ученый
Lane, H.Y. et al. Ингибитор переносчика глицина I, N -метилглицин (саркозин), добавленный к клозапину для лечения шизофрении. Biol.Психиатрия 60 , 645–649 (2006).
Артикул CAS PubMed Google ученый
Lane, H. Y. et al. Рандомизированное двойное слепое плацебо-контролируемое сравнительное исследование саркозина ( N -метилглицин) и дополнительного лечения шизофрении D-серином. Внутр. J. Neuropsychopharmacol. 13 , 451–460 (2010).
Артикул CAS PubMed Google ученый
Цай, Г., Lane, H.Y., Yang, P., Chong, M.Y. & Lange, N. Ингибитор переносчика глицина I, N -метилглицин (саркозин), добавленный к антипсихотическим средствам для лечения шизофрении. Biol. Психиатрия 55 , 452–456 (2004).
Артикул CAS PubMed Google ученый
Lane, H. Y. et al. Лечение острой шизофрении саркозином ( N -метилглицин): рандомизированное двойное слепое исследование. Biol. Психиатрия 63 , 9–12 (2008).
Артикул CAS PubMed Google ученый
Джавитт, Д. К., Дункан, Л., Балла, А. и Сершен, Х. Ингибирование системы А-опосредованного транспорта глицина в кортикальных синаптосомах терапевтическими концентрациями клозапина: влияние на механизмы действия. Мол. Психиатрия 10 , 275–287 (2005).
Артикул CAS PubMed Google ученый
Leucht, S.и другие. Сравнительная эффективность и переносимость 15 антипсихотических препаратов при шизофрении: метаанализ нескольких методов лечения. Ланцет 382 , 951–962 (2013).
Артикул CAS Google ученый
van Berckel, B. N. et al. Эффективность и переносимость D-циклосерина у больных шизофренией, не употребляющих наркотики. Biol. Психиатрия 40 , 1298–1300 (1996).
Артикул CAS PubMed Google ученый
Вудс, С.W. et al. Лечение глицином синдрома риска психоза: отчет о двух пилотных исследованиях. Eur. Neuropsychopharmacol. 23 , 931–940 (2013).
Артикул CAS PubMed PubMed Central Google ученый
Pinard, E. et al. Селективные ингибиторы GlyT1: открытие [4- (3-фтор-5-трифторметилпиридин-2-ил) пиперазин-1-ил] [5-метансульфонил-2 — (( S ) -2,2,2-трифтор- 1-метилэтокси) фенил] метанон (RG1678), новое многообещающее лекарство для лечения шизофрении. J. Med. Chem. 53 , 4603–4614 (2010).
Артикул CAS PubMed Google ученый
Umbricht, D. et al. Ингибитор переносчика глицина типа 1 (GLYT1) RG1678: положительные результаты экспериментального исследования по лечению негативных симптомов шизофрении. Нейропсихофармакология 35 , S320 – S321 (2010).
Google ученый
Мартин-Факлам, М.и другие. Зависимость переносчика глицина типа 1 от битопертина: исследование с использованием позитронно-эмиссионной томографии у здоровых добровольцев. Нейропсихофармакология 38 , 504–512 (2013).
Артикул CAS PubMed Google ученый
Моландер, А., Лидо, Х. Х., Лоф, Э., Эриксон, М. и Содерпалм, Б. Ингибитор обратного захвата глицина Org25935 снижает потребление этанола и снижает его предпочтение у самцов крыс линии Вистар. Алкоголь Алкоголь. 42 , 11–18 (2007).
Артикул CAS PubMed Google ученый
Сёдерпалм, Б. и Эриксон, М. Нейросхемы, участвующие в развитии алкогольной зависимости: дофаминовая система и точки доступа к ней. Curr. Верхний. Behav. Neurosci. 13 , 127–161 (2013). Это исследование предлагает модель алкогольной зависимости, которая связывает действие глицина и алкоголя с механизмом вознаграждения, и дает обоснование для ингибирования обратного захвата переносчика глицина как средства снижения потребления алкоголя и рецидивов.
Артикул CAS PubMed Google ученый
Uslaner, J. M. et al. Ингибирование транспортера глицина 1 ослабляет вызванное никотином, но не пищевым стимулом, восстановление ответа, ранее связанного с сахарозой. Behav. Brain Res. 207 , 37–43 (2010).
Артикул CAS PubMed Google ученый
Ахат-Мендес, К., Nic Dhonnchadha, B. A., Platt, D. M., Kantak, K. M. & Spealman, R. D. Ингибирование переносчика глицина-1, предшествующее обучению угасанию, подавляет повторное приобретение поиска кокаина. Нейропсихофармакология 37 , 2837–2845 (2012).
Артикул CAS PubMed PubMed Central Google ученый
Nic Dhonnchadha, B.A. et al. Ингибирование переносчика глицина-1 способствует исчезновению кокаинового сигнала и ослабляет повторное приобретение кокаинового поведения. Зависимость от наркотиков и алкоголя. 122 , 119–126 (2012).
Артикул CAS PubMed Google ученый
Венгелиене, В., Леонарди-Эссманн, Ф., Зоммер, У. Х., Марстон, Х. М. и Спанагель, Р. Блокада переносчика глицина-1 приводит к стойкому сокращению рецидивов употребления алкоголя у крыс. Biol. Психиатрия 68 , 704–711 (2010).
Артикул CAS PubMed Google ученый
Лидё, Х.Х., Стомберг, Р., Фагерберг, А., Эриксон, М. и Содерпалм, Б. Ингибитор обратного захвата глицина Org 25935 взаимодействует с базальным и индуцированным этанолом высвобождением дофамина в прилежащем ядре крысы. Alcohol Clin. Exp. Res. 33 , 1151–1157 (2009).
Артикул CAS PubMed Google ученый
Szegedi, A. et al. Оценка ингибитора переносчика глицина Org 25935 для профилактики рецидивов у пациентов с алкогольной зависимостью: рандомизированное двойное слепое многопрофильное плацебо-контролируемое исследование. Нейропсихофармакология 38 , S314 – S446 (2012).
Артикул Google ученый
Моландер, А., Лоф, Э., Стомберг, Р., Эриксон, М. и Содерпалм, Б. Вовлечение прилежащих рецепторов глицина в регуляцию добровольного потребления этанола у крыс. Alcohol Clin. Exp. Res. 29 , 38–45 (2005).
Артикул CAS PubMed Google ученый
Эггерс, Э.Д. и Бергер, А. Дж. Механизмы модуляции каналов нативных рецепторов глицина этанолом. J. Neurophysiol. 91 , 2685–2695 (2004).
Артикул CAS PubMed Google ученый
Спанагель Р. и Венгелиене В. Новые стратегии фармакологического лечения для предотвращения рецидивов. Curr. Верхний. Behav. Neurosci. 13 , 583–609 (2013).
Артикул CAS PubMed Google ученый
Wee, S.И Кооб, Г. Ф. Роль опиоидной системы динорфин-каппа в усиливающих эффектах злоупотребления наркотиками. Psychopharmacol. (Берл.) 210 , 121–135 (2010).
Артикул CAS Google ученый
Манн К., Бладстром А., Торуп Л., Гуаль А. и ван ден Бринк В. Расширение возможностей лечения алкогольной зависимости: рандомизированное контролируемое исследование налмефена по мере необходимости. Biol. Психиатрия 73 , 706–713 (2013).
Артикул CAS PubMed PubMed Central Google ученый
Spanagel, R. et al. Комплексная сеть исследования генома для изучения генетики алкогольной зависимости. Наркоман. Биол. 15 , 369–379 (2010).
Артикул CAS PubMed Google ученый
Лидо, Х. Х., Марстон, Х., Эриксон, М. и Содерпалм, Б.Ингибитор обратного захвата глицина Org24598 и акампросат снижают потребление этанола крысами; развитие толерантности к акампросату, но не к Org24598. Наркоман. Биол. 17 , 897–907 (2012).
Артикул CAS PubMed Google ученый
Vengeliene, V., Bachteler, D., Danysz, W. & Spanagel, R. Роль рецептора NMDA в рецидиве алкоголя: исследование фармакологического картирования с использованием эффекта алкогольной депривации. Нейрофармакология 48 , 822–829 (2005).
Артикул CAS PubMed Google ученый
Гасс, Дж. Т. и Олив, М. Ф. Глутаматергические субстраты наркомании и алкоголизма. Biochem. Pharmacol. 75 , 218–265 (2008).
Артикул CAS PubMed Google ученый
Сесак, С. Р. и Грейс, А.A. Сеть вознаграждения кортико-базальных ганглиев: микросхема. Нейропсихофармакология 35 , 27–47 (2010).
Артикул PubMed Google ученый
Engblom, D. et al. Рецепторы глутамата на дофаминовых нейронах контролируют постоянство поиска кокаина. Нейрон 59 , 497–508 (2008).
Артикул CAS PubMed Google ученый
Синдруп, С.Х., Отто, М., Финнеруп, Н. Б. и Дженсен, Т. С. Антидепрессанты в лечении невропатической боли. Bas. Clin. Pharmacol. Toxicol. 96 , 399–409 (2005).
Артикул CAS Google ученый
Núñez, E., Lopez-Corcuera, B., Vazquez, J., Gimenez, C. и Aragon, C. Дифференциальные эффекты трициклического антидепрессанта амоксапина на захват глицина, опосредованные рекомбинантными переносчиками глицина GLYT1 и GLYT2 . Br. J. Pharmacol. 129 , 200–206 (2000).
Артикул PubMed PubMed Central Google ученый
Tanabe, M., Takasu, K., Yamaguchi, S., Kodama, D. & Ono, H. Ингибиторы переносчиков глицина как потенциальная терапевтическая стратегия при хронической боли с нарушением памяти. Анестезиология 108 , 929–937 (2008).
Артикул CAS PubMed Google ученый
Морита, К.и другие. Спинальное антиаллодиническое действие ингибиторов транспортера глицина в моделях нейропатической боли у мышей. J. Pharmacol. Exp. Ther. 326 , 633–645 (2008).
Артикул CAS Google ученый
Дохи, Т., Морита, К., Китайма, Т., Мотояма, Н. и Мориока, Н. Ингибиторы переносчиков глицина как новая стратегия открытия лекарств от невропатической боли. Pharmacol. Ther. 123 , 54–79 (2009). В этом обзоре исследуются доказательства, подтверждающие роль ингибирования обратного захвата глицина в контроле над болью, и описываются соответствующие нейронные цепи обработки боли, которые мы представили в этой статье.
Артикул CAS PubMed Google ученый
Yoshikawa, S., Oguchi, T., Funahashi, Y., de Groat, W. C. & Yoshimura, N. Ингибитор транспортера глицина типа 2 (GlyT2) улучшает гиперактивность мочевого пузыря и ноцицептивное поведение у крыс. Eur. Урол. 62 , 704–712 (2012).
Артикул CAS PubMed PubMed Central Google ученый
Cheng, W. et al. Интрацеребровентрикулярная или интратекальная инъекция глицина вызывает анальгезию при тепловой ноцицепции и химической ноцицепции через рецепторы глицина. Eur. J. Pharmacol. 614 , 44–49 (2009).
Артикул CAS PubMed Google ученый
Уоллес, М.S. et al. Рандомизированное двойное слепое плацебо-контролируемое исследование антагониста глицина при нейропатической боли. Неврология 59 , 1694–1700 (2002).
Артикул CAS PubMed Google ученый
Бейер, К., Комисарук, Б. Р., Лопес-Колом, А. М. и Каба, М. Введение AP5, антагониста глутамата, демонстрирует обезболивающее действие глицина у крыс. Pharmacol. Biochem. Behav. 42 , 229–232 (1992).
Артикул CAS PubMed Google ученый
Сентено, М. В., Муцо, А., Миллекамп, М. и Апкариан, А. В. Префронтальная кора и спинной мозг, опосредованная антинейропатией и анальгезией, индуцированной саркозином, ингибитором транспортера глицина-Т1. Боль 145 , 176–183 (2009).
Артикул CAS PubMed PubMed Central Google ученый
Кодама, Д., Оно, Х. и Танабе, М. Повышенное поглощение глицина в гиппокампе и когнитивная дисфункция после повреждения периферических нервов. Боль 152 , 809–817 (2011).
Артикул CAS PubMed Google ученый
Werdehausen, R. et al. Метаболиты лидокаина ингибируют переносчик глицина 1: новый механизм обезболивающего действия системного лидокаина? Анестезиология 116 , 147–158 (2012).
Артикул CAS PubMed Google ученый
Hermanns, H. et al. Дифференциальные эффекты спинно применяемых ингибиторов транспортера глицина на ноцицепцию на крысиной модели нейропатической боли. Neurosci. Lett. 445 , 214–219 (2008).
Артикул CAS PubMed Google ученый
Хараниши, Ю., Хара, К., Терада, Т., Накамура, С. и Сата, Т. Антиноцицептивный эффект интратекального введения ингибитора транспортера глицина-2 ALX1393 на модели острой боли у крыс. Anesth. Анальг. 110 , 615–621 (2010).
Артикул CAS PubMed Google ученый
Nishikawa, Y., Sasaki, A. & Kuraishi, Y. Блокада транспортера глицина GlyT2, но не GlyT1, улучшает динамическую и статическую механическую аллодинию у мышей с герпетической или постгерпетической болью. J. Pharmacol. Sci. 112 , 352–360 (2010).
Артикул CAS PubMed Google ученый
Мантс, А.Г. и др. Интратекальный глицин при боли и дистонии при сложном региональном болевом синдроме. Боль 146 , 199–204 (2009).
Артикул CAS PubMed Google ученый
Вазири Р. Глицинотерапия шизофрении: некоторые предостережения. Biol. Психиатрия 39 , 155–156 (1996).
Артикул CAS PubMed Google ученый
Harsing, L.G. Jr et al. Переносчик глицина типа 1 и его ингибиторы. Curr. Med. Chem. 13 , 1017–1044 (2006).
Артикул CAS PubMed Google ученый
Янг, К. Р. и Свенссон, К. А. Аллостерическая модуляция рецептора NMDA через повышение уровня глицина и D-серина в головном мозге: терапевтические возможности при шизофрении. Pharmacol. Ther. 120 , 317–332 (2008).
Артикул CAS PubMed Google ученый
Уэллет, Д., Сазерленд, С., Ван, Т., Гриффини, П. и Мурти, В. Первое исследование на людях селективного ингибитора GlyT1 GSK1018921: взаимосвязь между воздействием и головокружением . Clin. Pharmacol. Ther. 90 , 597–604 (2011).
Артикул CAS PubMed Google ученый
Аткинсон, Б.N. et al. ALX 5407: мощный селективный ингибитор транспортера глицина hGlyT1. Мол. Pharmacol. 60 , 1414–1420 (2001).
Артикул CAS PubMed Google ученый
Howard, A. et al. Переносчик глицина GLYT1 необходим для опосредованной глицином защиты эпителиальных клеток кишечника человека от окислительного повреждения. J. Physiol. 588 , 995–1009 (2010).
Артикул CAS PubMed PubMed Central Google ученый
Джавитт, Д.C. Является ли сайт глицина наполовину насыщенным или наполовину ненасыщенным? Эффекты глутаматергических препаратов у больных шизофренией. Curr. Opin. Психиатрия 19 , 151–157 (2006).
Артикул PubMed Google ученый
Френч, Дж. А. и Фот, Э. Рациональная политерапия. Эпилепсия 50 (Приложение 8), 63–68 (2009).
Артикул CAS PubMed Google ученый
Тамминга, К.A. Когда полипрагмазия является преимуществом? Am. J. Психиатрия 168 , 663 (2011).
Артикул PubMed Google ученый
Рицнер, М.С. (ред.) Полифармация в практике психиатрии, Том II: Использование полипрагмазии в «реальном мире» (Springer, 2013).
Забронировать Google ученый
D’Souza, D. C. et al. Ингибитор переносчика глицина ослабляет психотомиметические эффекты кетамина у здоровых мужчин: предварительные данные. Нейропсихофармакология 37 , 1036–1046 (2012).
Артикул CAS Google ученый
Pinard, E. et al. Открытие бензоилпиперазинов как нового класса сильнодействующих и селективных ингибиторов GlyT1. Bioorg. Med. Chem. Lett. 18 , 5134–5139 (2008).
Артикул CAS Google ученый
Грей, Дж.А. и Николл Р. А. Мышление вне синапса: глицин на внесинаптических рецепторах NMDA. Cell 150 , 455–456 (2012).
Артикул CAS Google ученый
Mothet, J. P. et al. D-серин является эндогенным лигандом для сайта глицина N рецептора -метил-D-аспартата. Proc. Natl Acad. Sci. США 97 , 4926–4931 (2000).
Артикул CAS Google ученый
Розенберг, Д.и другие. Высвобождение нейронального D-серина и глицина через транспортер ASC-1 регулирует синаптическую активность, зависимую от рецептора NMDA. J. Neurosci. 33 , 3533–3544 (2013).
Артикул CAS PubMed PubMed Central Google ученый
Marchetti, M. et al. Связывание АТФ с сериновой рацемазой человека кооперативно и модулируется глицином. FEBS J. http://dx.doi.org/10.1111/febs.12510 (2013).
Musante, V., Summa, M., Cunha, RA, Raiteri, M. & Pittaluga, A. Взаимодействие пресинаптического транспортера глицина GlyT1 и рецептора NMDA: актуальность для активации ауторецептора NMDA в присутствии Mg 2+ ион. J. Neurochem. 117 , 516–527 (2011).
Артикул CAS Google ученый
Duguid, I.C. & Smart, T.G. in Biology of the NMDA Receptor (ed.Донген, А. М. В.) 313–328 (CRC Press, 2009).
Google ученый
Gelfin, E. et al. Адъювантная терапия D-серином облегчает поведенческие и двигательные симптомы при болезни Паркинсона. Внутр. J. Neuropsychopharmacol. 15 , 543–549 (2012).
Артикул CAS Google ученый
Шнайдер, Дж. С., Тинкер, Дж. П., Ван Велсон, М. и Джардиньер, М.Влияние частичного агониста глицина D-циклосерина на когнитивные функции у обезьян, получающих хронические низкие дозы MPTP. Brain Res. 860 , 190–194 (2000).
Артикул CAS Google ученый
Jimenez, E. et al. P2Y пуринергическая регуляция транспортеров нейротрансмиттеров глицина. J. Biol. Chem. 286 , 10712–10724 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Баркесс, Г.и другие. Хроматин-связывающий белок HMGN3 стимулирует ацетилирование гистонов и транскрипцию гена Glyt1 . Biochem. J. 442 , 495–505 (2012).
Артикул CAS PubMed PubMed Central Google ученый
Verissimo, C. et al. Некетотическая гиперглицинемия: причина энцефалопатии у детей. J. Child Neurol. 28 , 251–254 (2013).
Артикул PubMed Google ученый
Эпплгарт, Д.А. и Тоун, Дж. Р. Глициновая энцефалопатия (некетотическая гиперглицинемия): комментарии и предположения. Am. J. Med. Genet. A 140A , 186–188 (2006).
Артикул Google ученый
Conter, C. et al. Генетическая гетерогенность гена GLDC у 28 неродственных пациентов с глициновой энцефалопатией. J. Inherit Metab. Дис. 29 , 135–142 (2006).
Артикул CAS PubMed Google ученый
Мэр, Ф.Jr et al. Атипичная некетотическая гиперглицинемия с нарушением системы транспорта глицина в нервной ткани. Neurochem. Патол. 2 , 233–249 (1984).
PubMed Google ученый
Jursky, F. & Nelson, N. Экспрессия развития транспортеров глицина GLYT1 и GLYT2 в мозге мышей. J. Neurochem. 67 , 336–344 (1996).
Артикул CAS PubMed Google ученый
Шианг Р.и другие. Мутации в α1-субъединице ингибирующего рецептора глицина вызывают доминантное неврологическое расстройство — гиперэкплексию. Nature Genet. 5 , 351–358 (1993).
Артикул CAS PubMed Google ученый
Chung, S. K. et al. Патофизиологические механизмы доминантных и рецессивных мутаций GLRA1 при гиперэкплексии. J. Neurosci. 30 , 9612–9620 (2010).
Артикул CAS PubMed PubMed Central Google ученый
Chung, S. K. et al. GLRB — третий главный ген, влияющий на гиперэкплексию. Гум. Мол. Genet. 22 , 927–940 (2013).
Артикул CAS Google ученый
Джеймс, В. М. и др. Новые миссенс-мутации в гене бета-субъединицы глицинового рецептора (, GLRB ) при болезни испуга. Neurobiol. Дис. 52 , 137–149 (2013).
Артикул CAS PubMed PubMed Central Google ученый
Gimenez, C. et al. Новая доминантная мутация гиперэкплексии Y705C изменяет транспорт и биохимические свойства пресинаптического транспортера глицина GlyT2. J. Biol. Chem. 287 , 28986–29002 (2012).
Артикул CAS PubMed PubMed Central Google ученый
Шарлье, К.и другие. Высокоэффективное картирование ассоциаций на основе SNP и управление рецессивными дефектами домашнего скота. Nature Genet. 40 , 449–454 (2008).
Артикул CAS PubMed PubMed Central Google ученый
Gill, J. L. et al. Болезнь испуга у ирландских волкодавов, связанная с микроделецией в гене транспортера глицина GlyT2 . Neurobiol. Дис. 43 , 184–189 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Боме И. и Лудденс Х. Тормозные нейронные цепи как мишень противоэпилептических препаратов. Curr. Med. Chem. 8 , 1257–1274 (2001).
Артикул CAS PubMed PubMed Central Google ученый
Зайлер Н. и Сархан С. Синергетические противосудорожные эффекты агониста ГАМК и глицина. Gen. Pharmacol. 15 , 367–369 (1984).
Артикул CAS PubMed PubMed Central Google ученый
Halsey, M. J., Little, H. J. и Wardley-Smith, B. Системно вводимый глицин защищает мышей от стрихниновых судорог, но не от поведенческих эффектов высокого давления. J. Physiol. 408 , 431–441 (1989).
Артикул CAS PubMed PubMed Central Google ученый
Петерсон, С.L. Глицин усиливает противосудорожное действие диазепама и фенобарбитала при возбужденных амигдалоидных припадках у крыс. Нейрофармакология 25 , 1359–1363 (1986).
Артикул CAS PubMed PubMed Central Google ученый
Ларсон, А. и Бейтц, А. Дж. Глицин потенцирует судороги, вызванные стрихнином: роль рецепторов NMDA. J. Neurosci. 8 , 3822–3826 (1988).
Артикул CAS PubMed PubMed Central Google ученый
Семьянов А., Уокер М. К., Куллманн Д. М. и Сильвер Р. А. Тонически активные рецепторы ГАМК: модуляция усиления и поддержание тона. Trends Neurosci. 27 , 262–269 (2004).
Артикул CAS PubMed PubMed Central Google ученый
Эйхлер, С.A. et al. Сплайс-специфическая роль рецептора глицина α3 в гиппокампе. Eur. J. Neurosci. 30 , 1077–1091 (2009).
Артикул PubMed PubMed Central Google ученый
Chen, R.Q. et al. Роль рецепторов глицина в индуцированной глицином LTD в пирамидных нейронах CA1 гиппокампа. Нейропсихофармакология 36 , 1948–1958 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Чжан Л.H., Gong, N., Fei, D., Xu, L. & Xu, T. L. Поглощение глицина регулирует активность сети гиппокампа посредством тонического ингибирования, опосредованного глициновыми рецепторами. Нейропсихофармакология 33 , 701–711 (2008). В этой статье показано, что ингибирование GlyT1 в гиппокампе способствует индукции зависимого от рецептора NMDA LTP и одновременно усиливает ингибирование нейронов через рецепторы глицина, тем самым указывая на то, что GlyT1 может эффективно настраивать как возбуждающую, так и ингибирующую активности в гиппокампе посредством регуляции торговля глицином.
Артикул CAS PubMed PubMed Central Google ученый
Socała, K., Nieoczym, D., Rundfeldt, C. & Wlaz, P. Эффекты саркозина, ингибитора переносчика глицина типа 1, на двух моделях судорог у мышей. Pharmacol. Реп. 62 , 392–397 (2010).
Артикул PubMed PubMed Central Google ученый
Калиничев, М.и другие. Ингибиторы переносчика глицина 1 (GlyT1) проявляют противосудорожные свойства в тесте на максимальный порог электрошока на крысах (MEST). Brain Res. 1331 , 105–113 (2010).
Артикул CAS PubMed PubMed Central Google ученый
Петерсон, С.Л. Усиление противосудорожного препарата глицином при максимальных приступах электрошока имитируется D-серином и антагонистом 7-хлорокинуреновой кислоты. Eur. J. Pharmacol. 199 , 341–348 (1991).
Артикул CAS PubMed PubMed Central Google ученый
Петерсон, С.Л. и Шваде, Н.Д. Противосудорожная активность D-циклосерина специфична для тонических судорог. Epilepsy Res. 15 , 141–148 (1993).
Артикул CAS PubMed PubMed Central Google ученый
Wlaź, P., Баран, Х. и Лошер, В. Влияние частичного агониста глициновых / NMDA рецепторов, D-циклосерина, на порог судорожных припадков и некоторые фармакодинамические эффекты MK-801 у мышей. Eur. J. Pharmacol. 257 , 217–225 (1994).
Артикул PubMed PubMed Central Google ученый
Xue, J. G. et al. Активация рецептора NMDA усиливает ингибирующую ГАМКергическую передачу на пирамидные нейроны гиппокампа через пресинаптические и постсинаптические механизмы. J. Neurophysiol. 105 , 2897–2906 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Перри, Т. Л. и Хансен, С. Аминокислотные нарушения в эпилептогенных очагах. Неврология 31 , 872–876 (1981).
Артикул CAS PubMed PubMed Central Google ученый
Фостер, А.К. и Кемп, Дж. А. Нейробиология. Глицин поддерживает возбуждение. Nature 338 , 377–378 (1989).
Артикул CAS PubMed PubMed Central Google ученый
Краучер, М. Дж. И Брэдфорд, Х. Ф. 7-хлорокинуреновая кислота, нечувствительный к стрихнину антагонист глициновых рецепторов, ингибирует киндлинг лимбических припадков. Neurosci. Lett. 118 , 29–32 (1990).
Артикул CAS PubMed PubMed Central Google ученый
Бристоу, Л.J. et al. Противосудорожное средство и поведенческий профиль L-701 324, мощного перорально активного антагониста в модулирующем сайте глицина на рецепторном комплексе N -метил-D-аспартат. J. Pharmacol. Exp. Ther. 279 , 492–501 (1996).
CAS PubMed PubMed Central Google ученый
Stelzer, A., Slater, N. T. & ten Bruggencate, G. Активация рецепторов NMDA блокирует ГАМКергическое ингибирование в модели in vitro эпилепсии. Nature 326 , 698–701 (1987).
Артикул CAS PubMed Google ученый
Endele, S. et al. Мутации в GRIN2A и GRIN2B , кодирующих регуляторные субъединицы рецепторов NMDA, вызывают вариабельные фенотипы развития нервной системы. Nature Genet. 42 , 1021–1026 (2010).
Артикул CAS PubMed Google ученый
Карвилл, Г.L. et al. GRIN2A Мутации вызывают расстройства эпилепсии-афазического спектра. Nature Genet. 45 , 1073–1076 (2013).
Артикул CAS PubMed Google ученый
Lemke, J. R. et al. Мутации в GRIN2A вызывают идиопатическую фокальную эпилепсию с роландическими шипами. Nature Genet. 45 , 1067–1072 (2013).
Артикул CAS PubMed Google ученый
Леска, Г.и другие. GRIN2A. мутации при приобретенной эпилептической афазии и родственных детских фокальных эпилепсиях и энцефалопатиях с речевой и языковой дисфункцией. Nature Genet. 45 , 1061–1066 (2013).
Артикул CAS PubMed Google ученый
Мюллер, Э., Ле-Коррон, Х. и Лежандр, П. Внесинаптические и постсинаптические рецепторы в глицинергической и ГАМКергической нейротрансмиссии: разделение труда? Фронт.Мол. Neurosci. 1 , 3 (2008).
Артикул CAS PubMed PubMed Central Google ученый
Li, Y. & Xu, T. L. Зависящее от состояния перекрестное ингибирование между анионными ГАМК и ионотропными рецепторами глицина в нейронах СА1 гиппокампа крысы. Нейроотчет 13 , 223–226 (2002).
Артикул PubMed Google ученый
Бреустедт, Дж., Schmitz, D., Heinemann, U. & Schmieden, V. Характеристика ингибирующего рецептора глицина на нейронах энторинальной коры. Eur. J. Neurosci. 19 , 1987–1991 (2004).
Артикул PubMed Google ученый
Броди, М. Дж. И Силлс, Г. Дж. Комбинирование противоэпилептических препаратов — рациональная политерапия? Изъятие 20 , 369–375 (2011).
Артикул Google ученый
Гринберг, В.M. et al. Дополнительный глицин в лечении обсессивно-компульсивного расстройства у взрослых. J. Psychiatr. Res. 43 , 664–670 (2009).
Артикул PubMed Google ученый
Ву, П. Л., Тан, Х. С., Лейн, Х. Ю., Цай, К. А. и Цай, Г. Е. Саркозиновая терапия при обсессивно-компульсивном расстройстве: проспективное открытое исследование. J. Clin. Psychopharmacol 31 , 369–374 (2011).
Артикул CAS PubMed Google ученый
Карлссон, М. Л. О роли коркового глутамата в обсессивно-компульсивном расстройстве и синдроме дефицита внимания с гиперактивностью, двух феноменологически противоположных состояниях. Acta Psychiatr. Сканд. 102 , 401–413 (2000).
Артикул CAS PubMed Google ученый
Ву, К., Ханна, Г. Л., Розенберг, Д. Р. и Арнольд, П. Д. Роль передачи сигналов глутамата в патогенезе и лечении обсессивно-компульсивного расстройства. Pharmacol. Biochem. Behav. 100 , 726–735 (2012).
Артикул CAS PubMed Google ученый
Kushner, M. G. et al. Терапия с усиленным воздействием D-циклосерина при обсессивно-компульсивном расстройстве. Biol. Психиатрия 62 , 835–838 (2007).
Артикул CAS PubMed Google ученый
Wilhelm, S. et al. Расширение поведенческой терапии с помощью D-циклосерина при обсессивно-компульсивном расстройстве. Am. J. Psychiatry 165 , 335–341 (2008).
Артикул PubMed Google ученый
Ressler, K. J. et al. Когнитивные усилители в качестве дополнения к психотерапии: использование D-циклосерина у фобических людей для облегчения угасания страха. Arch. Общая психиатрия 61 , 1136–1144 (2004).
Артикул PubMed Google ученый
Дэвис М., Ресслер К., Ротбаум Б. О. и Ричардсон Р. Влияние D-циклосерина на вымирание: перевод из доклинической в клиническую работу. Biol. Психиатрия 60 , 369–375 (2006).
Артикул CAS PubMed Google ученый
Отто, М.W. et al. Эффективность D-циклосерина для усиления ответа на когнитивно-поведенческую терапию панического расстройства. Biol. Психиатрия 67 , 365–370 (2010).
Артикул CAS PubMed Google ученый
Guastella, A. J. et al. Рандомизированное контролируемое исследование D-циклосерина, улучшающего экспозиционную терапию при социальном тревожном расстройстве. Biol. Психиатрия 63 , 544–549 (2008).
Артикул CAS PubMed Google ученый
Hofmann, S.G. et al. Расширение экспозиционной терапии D-циклосерином при социальном тревожном расстройстве. Arch. Общая психиатрия 63 , 298–304 (2006).
Артикул CAS PubMed Google ученый
Наций, К. Р. и др. Оценка ингибитора переносчика глицина Org 25935 в качестве дополнения к когнитивно-поведенческой терапии панического расстройства: многоцентровое рандомизированное двойное слепое плацебо-контролируемое исследование. J. Clin. Психиатрия 73 , 647–653 (2012).
Артикул CAS PubMed Google ученый
Huang, C.C. et al. Ингибирование транспортера глицина-1 как новый механизм лечения депрессии. Biol. Психиатрия http://dx.doi.org/10.1016/j.biopsych.2013.02.020 (2013).
Heresco-Levy, U. et al. Рандомизированное дополнительное исследование высоких доз D-циклосерина для лечения устойчивой депрессии. Внутр. J. Neuropsychopharmacol. 16 , 501–506 (2013).
Артикул CAS PubMed Google ученый
Burgdorf, J. et al. GLYX-13, функциональный частичный агонист участка глицина рецептора NMDA, вызывает эффекты, подобные антидепрессантам, без побочных эффектов, подобных кетамину. Нейропсихофармакология 38 , 729–742 (2013).
Артикул CAS PubMed PubMed Central Google ученый
Катена-Делл’оссо, М., Fagiolini, A., Rotella, F., Baroni, S. & Marazziti, D. Глутаматная система как цель для разработки новых антидепрессантов. CNS Spectr. 18 , 188–198 (2013).
Артикул PubMed Google ученый
Perez-Siles, G. et al. Молекулярные основы дифференциального взаимодействия с литием переносчиков глицина GLYT1 и GLYT2. J. Neurochem. 118 , 195–204 (2011).
Артикул CAS PubMed Google ученый
Полешак, Э.и другие. Сложное взаимодействие между глициновыми / NMDA рецепторами и серотонинергическими / норадренергическими антидепрессантами в тесте принудительного плавания на мышах. J. Neural Transm. 118 , 1535–1546 (2011).
Артикул CAS PubMed PubMed Central Google ученый
Vanhoof, G. et al. Гетерозиготы-переносчики глицина 1 демонстрируют снижение тревожности в новых условиях. Программа № 504.13. 2004 Планировщик встреч по неврологии (Сан-Диего, Калифорния; Общество неврологии, 2004).
Google ученый
Dubroqua, S. et al. Воздействие нарушения нейронального транспортера глицина 1 в переднем мозге в стареющем мозге: доказательства возрастных фенотипов в обучении по Павлову. Behav. Neurosci. 124 , 839–850 (2010).
Артикул PubMed PubMed Central Google ученый
Якобс, Б. Л., ван Прааг, Х.И Гейдж, Ф. Х. Нейрогенез мозга взрослых и психиатрия: новая теория депрессии. Мол. Психиатрия 5 , 262–269 (2000).
Артикул CAS PubMed Google ученый
Harvey, R.J. et al. GlyR α3: важная мишень для сенсибилизации боли, опосредованной PGE2 в позвоночнике. Наука 304 , 884–887 (2004). Это ключевое исследование раскрывает новую биологическую роль GlyRα3 в сенсибилизации центральной воспалительной боли.
Артикул CAS PubMed Google ученый
Rácz, I., Schütz, B., Abo-Salem, O. M. & Zimmer, A. Висцеральная, воспалительная и невропатическая боль у мышей с дефицитом глицинового рецептора α3. Нейроотчет 16 , 2025–2028 (2005).
Артикул PubMed Google ученый
Hösl, K. et al. Спинальные рецепторы простагландина E подтипа EP2 и субъединица α3 глицинового рецептора, которые опосредуют центральную воспалительную гипералгезию, не вызывают боли после повреждения периферического нерва или инъекции формалина. Боль 126 , 46–53 (2006).
Артикул CAS PubMed Google ученый
Харви, В. Л., Кейли, А., Мюллер, У. С., Харви, Р. Дж. И Диккенсон, А. Х. Селективная роль рецепторов глицина субъединицы α3 в воспалительной боли. Фронт. Мол. Neurosci. 2 , 14 (2009).
Артикул CAS PubMed PubMed Central Google ученый
Морита, К., Китайма, Т., Мориока, Н. и Дохи, Т. Глицинергическое опосредование тактильной аллодинии, индуцированной фактором активации тромбоцитов (PAF), посредством передачи сигналов глутамат-NO-циклический GMP в спинном мозге мышей. Боль 138 , 525–536 (2008).
Артикул CAS PubMed Google ученый
Xiong, W. et al. Каннабиноидное усиление рецепторов глицина способствует анальгезии, вызванной каннабисом. Nature Chem.Биол. 7 , 296–303 (2011). Это ключевое исследование картирует сайт связывания каннабиноидов на рецепторах глицина и демонстрирует, что каннабиноид-индуцированная анальгезия отсутствует у мышей, лишенных GlyRα3, но не у мышей с каннабиноидным рецептором 1 (CB1R) — и мышей с нокаутом CB2R.