РОЛЬ ОЖИРЕНИЯ В ЭТИОПАТОГЕНЕЗЕ РАКА ЭНДОМЕТРИЯ
И.И. Фролова
Госпиталь для ветеранов войн № 2 г. Москвы (тел.: 8 499 940 97 31)
В обзоре литературы представлен взгляд на ожирение как на один из основных факторов риска рака эндометрия первого патогенетического варианта.
Ключевые слова: рак эндометрия, ожирение, индекс массы тела, андрогены, эстрогены, ароматазы, лептин.
Рак эндометрия (РЭ) является самой распространенной онкогинекологической патологией в экономически развитых странах и занимает 7-е место среди причин смертности от злокачественных новообразований в Западной Европе [20]. В Европейском Союзе ежегодно заболевают 81500 женщин. В Российской Федерации в 2009 году у 18523 женщин диагностирован РЭ, стандартизованная заболеваемость составляет 15,53 на 100 000 женского населения. Многие годы РЭ занимает второе место среди онкологической патологии женщин, уступая лишь раку молочной железы, и первое место среди опухолей женской половой сферы [5].
Средний возраст возникновения РЭ равен 60 годам, но 90% женщин заболевает в возрасте старше 50 лет [31]. Отмечается тенденция роста заболеваемости среди женщин репродуктивного и перименопаузального возраста: до 40% от общего числа [5].
Разные факторы риска, гистопатологические варианты и молекулярные механизмы развития опухоли приводят к двум патогенетическим вариантам РЭ. Традиционный или гормонозависимый патогенетический вариант характеризуется возникновением и развитием опухоли на фоне хронической гиперэстрогении из предшествующей гиперплазии эндометрия. Альтернативный или гормононезависимый патогенетический вариант – развитие опухолей эндометрия при отсутствии гиперплазии и хронической гиперэстрогении, по течению и прогнозу он наиболее агрессивный.
Одним из факторов риска РЭ первого патогенетического варианта считается ожирение, более 40% случаев РЭ развиваются на его фоне. Ожирение повышает риск РЭ в любом возрасте, вне зависимости от применения КОК, курения, длительности менопаузы и наличия сахарного диабета [36, 37]. Умеренно повышают риск РЭ избыточный вес в пубертатном периоде и в молодом возрасте. Но в зрелом возрасте высокий индекс массы тела (ИМТ) уже достоверно повышает риск РЭ [40].
Умеренно повышают риск РЭ избыточный вес в пубертатном периоде и в молодом возрасте. Но в зрелом возрасте высокий индекс массы тела (ИМТ) уже достоверно повышает риск РЭ [40].
Ожирение – хроническое заболевание, характеризующееся избыточным накоплением жира в организме, при котором ИМТ составляет 30 кг/м2 и выше.
Ожирение рассматривается как ведущая среди потенциально устранимых причин смертности, уступая только табакокурению. К концу ХХ века по данным ВОЗ избыточную массу тела имело около 30% жителей нашей планеты, а это в абсолютных цифрах около 1,7 млрд. человек. Заболеваемость ожирением на планете за последнее десятилетие увеличилась на 75%, во всех регионах мира прогнозируется сохранение этой тенденции [33].
Несмотря на отсутствие широкомасштабных эпидемиологических исследований, в настоящее время в России не менее трети трудоспособного населения имеют избыточную массу тела, и четверть страдают ожирением. По данным эпидемиологов, у жителей мегаполисов ожирение встречается чаще, чем в сельской местности [4]. Ожирение поражает женщин на 20% чаще мужчин, и в 2 раза чаще у женщин регистрируется III степень. Пик развития ожирения приходится на возраст 45-55 лет. В 3-4 раза чаще ожирение встречается в возрасте 40-60 лет, чем в возрасте 15-20 лет [6].
По данным эпидемиологов, у жителей мегаполисов ожирение встречается чаще, чем в сельской местности [4]. Ожирение поражает женщин на 20% чаще мужчин, и в 2 раза чаще у женщин регистрируется III степень. Пик развития ожирения приходится на возраст 45-55 лет. В 3-4 раза чаще ожирение встречается в возрасте 40-60 лет, чем в возрасте 15-20 лет [6].
К сожалению, не существует единой патогенетической классификации ожирения. Информативным показателем степени ожирения считается ИМТ, который рассчитывается делением показателя массы тела в килограммах на квадрат показателя роста в метрах. По классификации массы тела по ИМТ, утверждённой ВОЗ в 1999 г., нормальным считается ИМТ в пределах 18,5-24,9, когда риск сопутствующих заболеваний для популяции в пределах среднего. ИМТ менее 18,5 говорит о дефиците массы тела, риск, соответственно, низкий. Предожирение и повышенный риск сопутствующей патологии вероятен при ИМТ 25,0-29,9. Ожирение I степени (ИМТ 30,0-34,9) уже существенно повышает риск сопряжённой с ним патологии.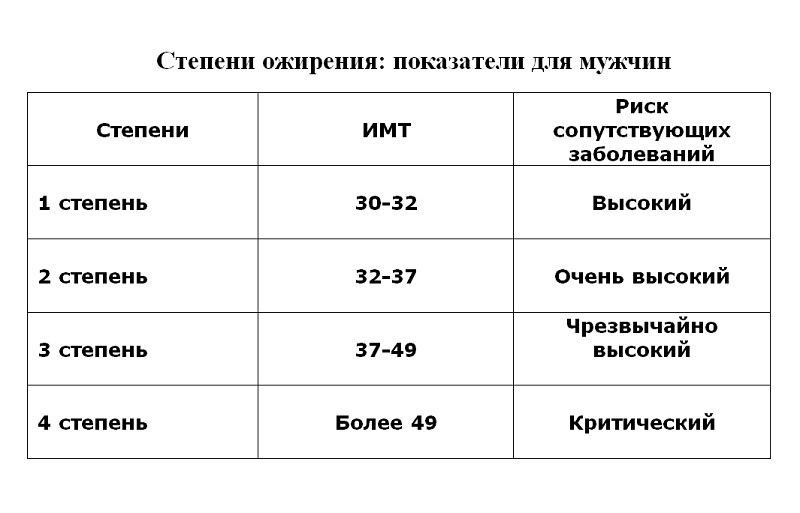
Кроме самого факта наличия ожирения и его степени, для оценки риска ассоциированных с ожирением заболеваний существенное значение имеет характер распределения жира. Оценка типа распределения жира проводится по результатам измерения окружности талии (ОТ) и окружности бедер (ОБ), а также соотношения этих показателей (ОТБ). Выделяют андроидный и гиноидный типы ожирения.
Для андроидного (мужского, висцерального, абдоминального) типа характерно отложение жира в верхних отделах туловища, особенно в области живота. При андроидном типе ОТБ превышает 0,85 для женщин и 0,95 для мужчин.
Гиноидное (женское, ягодичное) ожирение характеризуется преимущественным отложением жира в нижних отделах туловища. При нём ОТБ распределения жира менее 0,85 для женщин и менее 0,95 для мужчин. Нарушения углеводного обмена, сердечно-сосудистые заболевания регистрируются значительно чаще при андроидном типе ожирения. Характер распределения жировой клетчатки влияет и на степень риска РЭ: при андроидном типе распределения жировой клетчатки риск развития РЭ увеличивается в 6 раз. Риск РЭ даже при нормальном весе в 3,2 раза выше при андроидном типе [17].
Характер распределения жировой клетчатки влияет и на степень риска РЭ: при андроидном типе распределения жировой клетчатки риск развития РЭ увеличивается в 6 раз. Риск РЭ даже при нормальном весе в 3,2 раза выше при андроидном типе [17].
У женщин старшего возраста избыточная прибавка в весе ассоциирована с отсутствием или малым числом родов, ранним возрастом первых родов, патологической прибавкой веса во время беременности и коротким периодом грудного вскармливания [23].
В перименопаузе 60% женщин отмечает быстрое нарастание массы тела [27]. По данным специалистов Питсбургского университета, здоровые женщины в течение первых 3-х лет постменопаузы прибавляют в весе в среднем 2,5 кг, 20% набирают 5 кг и более. К 8-му году постменопаузы прибавка в весе в среднем на всю когорту составляет около 6-ти кг [39]. Увеличение массы тела на каждые 5 килограмм повышает риск РЭ в 1,2 раза; прибавка веса на 30 и более килограмм увеличивает риск РЭ в 3-4 раза. Кроме того, снижение собственного веса не снижает риска РЭ до величин, характерных для женщин никогда не страдавших ожирением [36].
Кроме того, снижение собственного веса не снижает риска РЭ до величин, характерных для женщин никогда не страдавших ожирением [36].
У женщин с ожирением в 4 раза чаще отмечаются различные нарушения менструального цикла, по сравнению с женщинами нормального веса. Ожирением страдают 45% пациенток с аменореей [19]. Прямая связь ановуляции с ожирением установлена в исследовании S. Franks [14]. В то же время только у 9-13% женщин с нормальным менструальным циклом зарегистрирован избыток веса [19, 28].
Обращает на себя внимание совпадение возрастных пиков заболеваемости ожирением и РЭ, оба развиваются чаще всего в пре- и постменопаузе. По мнению ряда авторов для развития РЭ возможно большее значение имеет абсолютная масса жировой ткани, нежели относительный вес женщины [1, 8, 12].
ЭТИОПАТОГЕНЕЗ ОЖИРЕНИЯ
Ожирение – результат дисбаланса между получаемой и потребляемой энергией. Три пути расходования энергии организма: затраты на поддержание основных физиологических функций в стандартных условиях – основной обмен; специфическое динамическое действие пищи: дополнительный расход энергии на пищеварение и стимуляцию метаболизма, обусловленный притоком нового субстрата – термогенный эффект и физическая активность – наибольший расход.
Известны такие функции жировой ткани как депонирование энергии и воды в организме; участие в процессах обмена жировых кислот и углеводов, в синтезе жира из углеводов, в регуляции фосфорно-кальциевого обмена (начальные этапы синтеза витамина D), в метаболизме половых стероидов. Сравнительно недавно установлено, что жировая ткань является так же источником синтеза ряда факторов, обладающих эндокринным, паракринным и аутокринным действиями.
Обсуждается непосредственная роль самой жировой ткани в генезе ожирения. Установлено, что преимущественно при ожирении происходит гипертрофия адипоцитов, однако при тяжелых формах заболевания наблюдается также увеличение и количества жировых клеток. Существует «критическая жировая масса», являющаяся триггером для образования новых адипоцитов, и это подтвердил эксперимент.
Установлено, что преимущественно при ожирении происходит гипертрофия адипоцитов, однако при тяжелых формах заболевания наблюдается также увеличение и количества жировых клеток. Существует «критическая жировая масса», являющаяся триггером для образования новых адипоцитов, и это подтвердил эксперимент.
Роль нарушения регуляции энергетического баланса в предрасположенности к развитию ожирения, в частности дисбаланса различных пептидов (лептина, грелина и др.) и нейропептидов (нейропептида Y и др.) также обсуждается. Полипептидный гормон лептин, синтезируемый и секретируемый жировыми клетками, участвует в регуляции энергетического обмена, уменьшая аппетит и повышая расход энергии путем механизма обратной связи между его уровнем и выработкой участвующего в формировании чувства голода гипоталамического нейропептида Y [6]. Ведущая функция лептина – регуляция внутриклеточного гомеостаза жирных кислот, предотвращение развития эктопического отложения жира в тканях, в норме не депонирующих жир, в периоды переедания – липотоксикоза [41].
У подавляющего большинства тучных людей уровень лептина повышен. С увеличением тучности уровень лептина в крови возрастает и снижается при уменьшении количества жировой ткани. Однако при наступлении так называемой «точки липостаза», когда за короткий период времени человек теряет более 30 кг, лептин запускает цикл противодействия снижению веса, и аппетит растёт, а человек вновь набирает массу. Продукция лептина в подкожной жировой клетчатке выше, чем в жировых депо. Лептин циркулирует в связанном с транспортным белком виде, резистентность к нему объясняется аномалиями в структуре лептин-связывающего белка – «лептинорезистентность». Возможной причиной лептинорезистентности является аномалия рецепторов гипоталамуса. Согласно теории, выдвинутой E. Petridou [29], в секреторную фазу менструального цикла лептин ингибирует синтез прогестерона. Показано, что высокие концентрации лептина у женщин с ожирением снижают протекторное действие прогестерона на эндометрий [18]. Установлена корреляция уровня лептина и повышения риска РЭ [29].
Изучена роль серотонина в формировании пищевого поведения: повышение его уровня в гипоталамических структурах в эксперименте вызывает чувство сытости. Подобным действием обладает и дофамин – при повышении активности катехоламинов потребность в еде снижается [6]. Увеличение содержания белка грелина также усиливает чувство голода [6]. Нарушения обмена серотонина и рецепции эндорфинов влекут изменения в психоэмоциональной сфере, что напрямую отражается и на пищевом поведении. В данном контексте употребление углеводистых продуктов является своеобразным допингом, а ожирение имеет сходство с такими болезнями, как алкоголизм и наркомания.
Не вызывает сомнений наличие генетической предрасположенности к ожирению. При избытке массы тела у одного из родителей, ожирение развивается у 40% детей. Часто обсуждается вопрос о доминирующем значении семейного стереотипа питания, однако исследования с раздельно живущими однояйцовыми близнецами подтверждают именно роль наследственного фактора. Наследственный фактор в настоящее время рассматривается как полигенный – открыто множество генов, отвечающих за развитие ожирения [6]. Выявлены мутации в гене липопротеинлипазы у больных с ожирением и сахарным диабетом II типа, изучается роль рецептора, активирующего пролифератор пероксисом PPAR.
Наследственный фактор в настоящее время рассматривается как полигенный – открыто множество генов, отвечающих за развитие ожирения [6]. Выявлены мутации в гене липопротеинлипазы у больных с ожирением и сахарным диабетом II типа, изучается роль рецептора, активирующего пролифератор пероксисом PPAR.
Возрастное повышение активности регуляторных центров гипоталамуса и снижение их чувствительности к стимулирующим или тормозящим влияниям с периферии, играет значимую роль в формировании ожирения. Процесс старения организма сопровождается изменением соотношения между гормонами, способствующими мобилизации жира, и гормонами, активирующими липогенез в пользу преобладания гормонов, способствующих концентрации жира в висцеральных депо [13]. Снижение скорости основного обмена в постменопаузе у женщин выражено в большей степени, чем у мужчин того же возрастного периода. В менопаузу изменяется и соотношение пептидов и нейромедиаторов, участвующих в регуляции центра голода и насыщения: уровень анорексигенных факторов (лептина, дофамина, холецистокинина) значительно ниже, чем у женщин в пременопаузе. В то же время, содержание орексигенных факторов (нейропептида Y, галанина, норадреналина) повышено [7].
В то же время, содержание орексигенных факторов (нейропептида Y, галанина, норадреналина) повышено [7].
Результаты ряда клинических и экспериментальных исследований демонстрируют и участие эстрогенов в регуляции пищевого поведения. Известно, что такие нарушения пищевого поведения, как булимия, анорексия и пищевые эксцессы встречаются преимущественно у женщин [16].
ОЖИРЕНИЕ В ЭТИОПАТОГЕНЕЗЕ РАКА ЭНДОМЕТРИЯ
При ожирении в жировой ткани увеличивается конверсия андрогенов в эстрогены, что повышает риск развития РЭ. Яичники пременопаузальных женщин в норме продуцируют прогестерон. В пременопаузе, особенно у тучных женщин, превалируют ановуляторные циклы, и эстрогены остаются не уравновешенными эндогенным прогестероном, что и ведёт к повышению риска РЭ [22]. В норме в постменопаузе яичники не продуцируют ни эстрогены, ни прогестерон; эстрогены образуются из андростендиола, секретируемого надпочечниками. При посредстве ароматаз происходит конверсия андростендиола в эстрон, а превращение эстрона в эстрадиол происходит в периферической жировой ткани. Поскольку яичники в постменопаузе не вырабатывают прогестерон, концентрация эстрадиола сыворотки крови очень высока. Повышение риска РЭ в постменопаузе в 5-10 раз связывают именно с отсутствием выработки овариального прогестерона [21]. У женщин нормальной массы тела только 1% андростендиона конвертируется в эстрон, в то время как при ожирении конверсия достигает уже 10% [2, 9]. Уровень эстрогенемии у постменопаузальных женщин в большей степени определяется именно массой тела, второстепенны возраст и продолжительность менопаузы [2].
Сопровождающие ожирение нарушения энергетического баланса, повышение ИМТ и снижение уровня белка, связывающего половые стероиды (БСПС), – факторы, повышающие риск развития РЭ. БСПС – гликопротеин, имеющий специфическое сродство к эстрогенам и андрогенам, к снижению уровня БСПС приводит повышение ИМТ [21]. При изменении ИМТ у женщин в пременопаузе при возрастании массы тела более чем на 30 кг, снижается уровень БСПС и соответственно повышается количество свободного эстрадиола в циркулирующей крови. Исследование N. Potischman и соавт. [30] подтверждает ассоциацию ожирения с повышением уровня сывороточного эстрадиола как следствия снижения концентрации БСПС, связывающего эстрогены.
Таким образом, ожирение способствует развитию гиперпластических и неопластических процессов в эндометрии. При этом роль ожирения рассматривается в нескольких ракурсах: как источник эндогенного образования эстрогенов из андрогенов и как комплекс метаболических сдвигов, потенцирующих вероятность развития опухолей [5, 32]. Дисрегуляция эндогенных стероидов и факторов роста является отправной точкой для нарушения баланса между процессами клеточной пролиферации и апоптозом. Хроническое, не уравновешенное эндогенным прогестероном или его синтетическими аналогами, воздействие эстрогенов на эндометрий приводит к повышению митотической активности клеток эндометрия и, в свою очередь, повышению репликации ДНК и накоплению соматических мутаций с формированием гиперплазии и малигнантного фенотипа [10].
Атипическая гиперплазия эндометрия может расцениваться как облигатный предрак только в сочетании с комплексом обменно-эндокринных нарушений [5, 35]. Группа исследователей, изучавших инциденты РЭ агрессивного клинического течения, пришла к выводу, что риск РЭ высокой степени агрессии достоверно выше у женщин с ожирением, родивших менее двух детей, принимающих более 8 лет ЗГТ эстрогенными препаратами, с сопутствующим сахарным диабетом [38]. При сочетании ожирения и сахарного диабета, троекратно увеличивается риск РЭ по сравнению с не страдающими диабетом женщинами [7]. Сахарный диабет повышает риск РЭ только у женщин с ожирением [11]. Установлено, что ожирение способствует манифестации генетически детерминированной гиперинсулинемической инсулин-резистентности [9]. E. Weiderpass и соавт. показали, что только у тучных женщин артериальная гипертония является фактором риска РЭ [37].
ПРОФИЛАКТИКА ОЖИРЕНИЯ КАК ПРОФИЛАКТИКА РАКА ЭНДОМЕТРИЯ
Несомненна роль санитарно-просветительной работы и превентивных мероприятий среди находящихся в группе риска по развитию ожирения детей и взрослых в профилактике ожирения. Развитию и закреплению устойчивой мотивации к предупреждению и лечению избыточного веса не только с эстетической целью, но и с целью профилактики опасного для жизни заболевания будет способствовать внедрение в массы знаний об ожирении как об одном из основных факторов риска развития РЭ.
Учитывая тот факт, что в основе ожирения всегда лежит дисбаланс между количеством потребляемой с пищей энергии и энергозатратами организма, основной целью коррекции питания является создание энергетического дефицита. Исследование A.J. Littman и соавт. [25] подтверждают достоверное снижение риска РЭ у женщин, придерживающихся низкокалорийной диеты с высоким содержанием овощей и фруктов. При возрастании физической активности уменьшается количество висцерального жира и предотвращается снижение мышечной массы при похудании [34]. Расходование энергии важно для поддержания оптимального веса.
«Проект здорового образа жизни» изучал возможности профилактики прибавки веса у 535 здоровых пременопаузальных женщин среднего возраста 47 лет, с ИМТ от 20 до 34 [24]. В основной группе из 275 человек проводилась коррекция образа жизни: снижение общей калорийности пищи, ограничение употребления жиров до 25% суточного рациона, повышение физической активности. 260 женщин контрольной группы не получали подобных рекомендаций. К концу периода наблюдения в 54 недели 35% женщин находились в постменопаузе. Уменьшение употребления жиров, особенно насыщенных, и холестерина с пищей у женщин в пременопаузе способствует статически значимому снижению массы тела, индекса ОТ/ОБ, а также темпов прироста триглицеридов, глюкозы и инсулина. Отмечена также корреляция снижения массы тела с повышением физической активности.
Анализ 13-ти исследований показал достоверную редукцию риска РЭ (на 30-40%) среди физически активных женщин [15]. C.E. Matthews с соавторами показали, что риск РЭ снижается и при интенсивных занятиях спортом (упражнения на тренажерах, бег и т.п.), и при умеренных физических нагрузках (пешеходные прогулки, танцы, садоводство, работа по дому) [26].
Подтверждение прямой связи ожирения с высоким риском РЭ свидетельствует о необходимости включения в государственные и международные программы профилактики РЭ положений, предупреждающих ожирение.
Литература
- Бохман Я.В. Руководство по онкогинекологии. – СПб: «ООО Издательство Фолиант», 2002. – С.195-229.
- Берштейн Л.М. Внегонадная продукция эстрогенов (роль в физиологии и патологии). – СПб.: Наука, 1998. – 172 с.
- Берштейн Л.М. Гормональный канцерогенез. – СПб.: Наука, 2000, – 200 с.
- Мельниченко Г.А., Романцева Т.И. Ожирение: эпидемиология, классификация, патогенез, клиническая симптоматика и диагностика.\ В кн. «Ожирение: этиология, патогенез, клинические аспекты» под ред. Дедова И.И., Мельниченко Г.А. – М.: ООО «Медицинское информационное агентство», 2006. – С.17-43.
- Под ред. Чиссова В.И., Старинского В.В., Петровой Г.В. Злокачественные новообразования в России в 2009 году (заболеваемость и смертность)/ М.: ФГУ «МНИОИ им. П.А. Герцена Минздравсоцразвития России», 2011.- С. 260
- Ожирение (клинические очерки) / Под ред. проф. Барановского А.Ю., проф. Ворохбиной Н.В. – СПб.: Издательство «Диалект», 2007. – 240 с.
- Романцева Т.И. Особенности патогенеза и лечения менопаузального метаболического синдрома./ В кн. «Ожирение: этиология, патогенез, клинические аспекты» под ред. Дедова И.И., Мельниченко Г.А. – М.: ООО «Медицинское информационное агентство», 2006. – С.219-236.
- Сафина Н.С. Оценка липидного обмена и иммуногенетических факторов у больных раком эндометрия. // Автореф. дисс. канд. мед. наук. – СПб. – 1999. – 38 с.
- Чернуха Г.Е. Аденоматозная и железистая гиперплазия эндометрия в репродуктивном возрасте (патогенез, клиника, лечение). // Автореф. дисс. д-ра мед. наук. – М. – 1999. – 42 с.
- Akhmedkhanov A., Zeleniuch-Jacquotte A., Toniolo P. Role of Exogenous and Endogenous Hormones in Endometrial Cancer. // Ann. NY Acad. Sci. – 2001; 943: 296-315.
- Anderson K.E., Anderson E., Mink P.J. et al. Diabetes and endometrial cancer in lowa women’s health study. // Canc. Epidemiol. Biomarc. Prevent. – 2001; 10 (6): 611-6.
- Baanders van Halewyn E.A., Blankenstein M.A., Thijssen J.H. et al. A comparative study of risk factors for hyperplasia and cancer of the endometrium. // Eur. J. Cancer. Prev. – 1996. – 5 (2): 105-112.
- Bjurntorp P. The regulation of adipose tissue distribution in humans. // Int. J. Obes. – 1997; 20: 291-302.
- Franks S., Willis D.S. et al. The effects of ovarian steroids, epidermal growth factor, insulin and insulin-like factor-1 on ovarian stromal growth. // Proceedings of Endocrine Society, 79th Annual Meeting, June 11-14, 1997. – San Diego: Endocrine society Press 1997; 389.
- Friednreich C., Orenstein M. Physical Activity and Cancer Prevention: Etiologic Evidence and Biological Mechanisms. // J. Nutrition. – 2002; 132: 3456-3464.
- Geary N. The effect of estrogen on appetite. // Medscape Woman’s Health J. – 1998; 3: 1-11.
- Elliot E.A., Matanoski G.M., Rosenshein N.B. Body fat patterning in women with endometrial cancer. // Gynecol. Oncol. – 1990; 39: 253.
- Hale G.E., Hughes C.L., Cline J.M. Endometrial Cancer: Hormonal Factors, The Premenopausal “Window of Risk”, and Isoflavones. // J. Clin. Endocr. Metab. – 2002; 87: 3-15.
- Hartz E.J., Borboriak P.N. et al. The association of obesity with infertility and related menstrual abnormalities in women. // Int. J. Obes. Relat. Metab. Disord. – 1979; 3: 57-73.
- Под редакцией Тюляндина С. А., Носова Д. А., Переводчиковой Н. И. Минимальные клинические рекомендации Европейского Общества Медицинской Онкологии (ЕSМО)/ М.: Издательская группа РОНЦ им. Н. Н. Блохина, 2010.— 436 с.
- Key T., Allen N., Verkasalo P., Banks E. Macronutrient Metabolism Group Symposium on “Energy Flux and Cancer.” Energy Balance and Cancer: The Role of Sex Hormones. // Proceed. Nutrit. Societ. – 2001; 60: 81-89.
- Key T.J., Pike M.C. The dose-effect relationship between “unopposed” oestrogens and endometrial: central role in explaining and predicting endometrial cancer risk. // Br. J. Cancer. – 1988; 57 (2): 20-24.
- Kirchengast S., Gruber D., Sator M. et al. Postmenopausal weight status, body composition and fat distribution in relation to parameters of menstrual and reproductive history.// Maturitas. – 1999; 33: 117-26.
- Kuller L.H., Simkin-Silwerman L.R., Wing R.R. et al. Women’s Healthy Lifestyle Project: a randomized clinical trial. Results at 54 months. / Circ. Am. Heart Assoc. – 2001; 2 (9): 32-7.
- Littman A.J., Berestford S.A., White E. The association of dietary fat and plant foods with endometrial cancer (United Stated).// Cancer Caus. Control. – 2001; 12 (8): 691-702.
- Matthews C.E., Xu W.X., Zheng W. et al. Physical Activity and Risk of Endometrial Cancer: A Report from the Shanghai Endometrial Cancer Study. // Canc. Epidemiol. Biomarc. Prevent. – 2005; 14: 779-85.
- Milewicz A., Bidzinska B., Sidorovicz A. Perimenopausal obesity. // Gynecol. Endocrinol. – 1996; 10: 285-91.
- Norman R.J., Clark A.M. Lifestyle factors in the aetiology and management of polycystic ovary syndrome. In: Polycystic ovary syndrome. Ed. By Gabor T. Kovacs. // Cambridge University Press. – 2000; 98-116.
- Petridou E., Belechri M., Dessypris N. et al. Leptin and Body mass Index in Relation to Endometrial Cancer Risk. // Ann. Nutrit. Metab. – 2002; 46: 147-51.
- Potischman N., Hoover R., Brinton L. et al. Case Control Study of Endogenous Steroid Hormones and Endometrial Cancer. // J. Nat. Cancer Inst. – 1996; 88: 1127-1135.
- Purdie D., Green A. Epidemiology of Endometrial Cancer. // Best Pract. Res. Clin. Obstet. Gynecol. – 2001; 15: 341-354.
- Qie M., Peng Z., Wang S. et al. Evaluation on ISGP classification of endometrial hyperplasia in clinical application. // Hua Hsi I Ko Ta Hsuch Pao. – 1997; 28 (1): 95-7.
- Seidell J.S. The Worldwide epidemic of obesity. / In: Progress in obesity research. 8-th International congress on obesity. B. Guy-Grand, G. Aithhaud, London: John Libbey & Company Ltd. 1999; 661-8.
- Shaw K., Gennat H., O’Rourke P., Del Mar C. Exercise for overweight or obesity. //Cochrane Database Syst. Rev. – 2006. – 18; (4): 321-3.
- Tabata T., Yamawaki T. at al. Clinical value of dilatation and curettage for abnormal uterine bleeding. // Arch. Gynecol. Obstet. – 2001; 264 (4): 174-6.
- Trentham-Dietz A., Nichols H.B., Hampton J.M., Newcomb P.A. Weight change and risk of endometrial cancer. // Int. J. Epidemiol. – 2006; 35 (1): 166-8.
- Weiderpass E., Persson I., Adami H.O. et al. Body size in different periods of life, diabetis mellitus, hypertension, and of postmenopausal endometrial cancer (Sweden). // Cancer Causes Control. – 2000; 11 (2): 185-92.
- Weiss J.M., Saltzman B.S., Doherty J.A. et al. Risk factors for the incidence of endometrial cancer according to the aggressiveness of disease. // Am. J. Epidemiol. – 2006; 164 (1): 56-62.
- Wing R., Matthews K., Kuller L. et al. Weight gain at time of menopause. // Ann. Int. Med. – 1991; 151: 97-102.
- Xu W.H., Xiang Y.B., Zhang X. et al. Weight history and risk of endometrial cancer among Chinese women. // Int. J. Epidemiol. – 2005; 35(1): 166-8.
- Unger R.H. Lipotoxic Diseases. // Annu. Rev. Med. – 2002; 53: 319-36.
ИЗБЫТОЧНАЯ МАССА ТЕЛА — Новоаганская районная больница
Избыточная масса тела, излишнее накопление жира в
организме и отложение его в подкожной клетчатке, сальнике и других
тканях и органах (главным образом вследствие нерационального питания и
малой двигательной активности). Является предстадией ожирения, в основе
которого лежит сформировавшееся преобладание жира из питательных
веществ, процессами его распада. Проблема избыточной массы тела и
ожирения стала серьезной угрозой для многих людей, приобрела социальный
характер. Прошло время, когда полнота считалась показателем благополучия
и здоровья. Полнеть — это, прежде всего, означает стареть, и не только
внешне. У людей с избыточным весом нарушается нормальная деятельность
сердечно-сосудистой системы, чаще развивается сахарный диабет,
атеросклероз, гипертоническая и желчно-каменная болезнь. Избыточная
масса тела влияет на продолжительность жизни, укорачивая ее на 10-15
лет.
Масса тела человека зависит в первую очередь от типа
телосложения, пола, возраста, служит относительным показателем
физического развития человека и состояния его здоровья. Для того, чтобы
сохранить фигуру красивой, а здоровье крепким, надо постоянно следить за
массой тела. Самый простой и доступный метод определения должной массы
тела является росто-весовой показатель, который соответствует величине
роста в сантиметрах минус 100. Масса тела считается избыточной, если она
превышает этот предел на 7% и более. При превышении массы тела над
пределом на 10-29% — ожирение 1 степени; на 30-49% — ожирение 2степени;
на 50-99% — ожирение 3 степени; свыше 100% — ожирение 4степени. При 1 и 2
степени ожирения трудоспособность и жизненная активность человека не
нарушены. Вообще провести четкую границу между здоровым и «упитанным»
человеком и больным начальной степенью ожирения во многих случаях
трудно. Недаром французские исследователи полушутливо выделяют такие 3
степени ожирения:
1 степень — когда окружающие завидуют;
2 степень — когда они смеются;
3 степень — когда они сочувствуют больному.
При
продолжающемся переедании и малоподвижном образе жизни одна стадия
незаметно переходит в другую. Выделяют две формы избыточной массы тела.
Первичную, при которой накопление жира в организме не связано с
какими-либо заболеваниями, а является следствием внешних воздействий.
Врачи издавна называли эту форму тучности «ожирением от переедания» или
«ожирением от лени». Она наиболее распространена и составляет более 70%
случаев ожирения. Значительно реже встречается вторичная форма нарушения
жирового обмена, связанная главным образом с эндокринными
расстройствами. Однако и при этой форме в той или иной степени имеет
место избыточное потребление пищи. Чаще всего избыточная масса тела
обусловлена неправильным и нерациональным питанием и образом жизни.
Ведущим в развитии избыточной массы тела и ожирения (около 60% первичных
случаев) является диетический дисбаланс, обусловленный избыточной
калорийностью пищи — перееданием энергоемких продуктов (сливочное масло,
сало, жирное мясо, мучные блюда, сахар, сладости, картофель),
содержащих повышенное количество жиров и углеводов, и недостаточной
физической активностью.
Установлено, что с возрастом пищевой центр
становится менее чувствительным к изменениям содержания глюкозы в крови и
для подавления чувства голода требуется более значительное количество
глюкозы в крови, чем в молодом возрасте, а значит и большее потребление
нищи. Этот фактор играет определенную роль в повышении аппетита и
тучности у людей пожилого возраста. Следует помнить о том, что чувство
голода у тучных людей проявляется гораздо сильнее, чем у людей с
нормальной массой тела, что стимулирует переедание.
Бичом
современного человека является малоподвижность. Первостепенная роль в
борьбе с тучностью принадлежит не врачу, а человеку, если он желает
иметь нормальную массу тела, а значит и здоровье. Для того, чтобы
ограничить себя в пище, следить за калорийностью продуктов рациона,
систематически заниматься физкультурой, необходимы сила воли,
собранность и настойчивость.
Ожирение
В течение длительного
времени ожирение не относилось к разряду смертельно опасных заболеваний.
Более того, избыточная масса тела нередко связывались с представлением о
здоровье. Конец ХХ века охарактеризовался большими успехами в развитии
диагностических методик, как биохимических, так и инструментальных,
которые позволили выявить зависимость развития и прогрессирования многих
тяжелых заболеваний от ожирения.
Ожирение, являясь заболеванием,
которое само по себе, значительно повышает риск преждевременной
смертности, одновременно увеличивает вероятность развития целой группы
заболеваний, каждое из которых во много раз повышает летальность среди
населения. Основными заболеваниями-спутниками ожирения являются
артериальная гипертензия, сахарный диабет, ишемическая болезнь сердца,
остеоартроз, дислипидемия.
Риск развития вышеперечисленных
сопутствующих заболеваний у лиц с ожирением, особенно с крайними его
степенями возрастает в 4-5 раз, что приводит к повышению летальности у
данной группы пациентов в 5-6 раз.
По причине развития ожирение
условно делится на гормональное (первично нарушение функции эндокринной
системы, приводящее к накоплению жировой ткани в организме) и
алиментарно-конституциональное. Причиной развития второго типа ожирения
является избыточное потребление энергии с пищей и недостаточный ее
расход в процессе жизнедеятельности. Практически во всех случаях
ожирение носит смешанный характер, так как при развитии избыточного веса
неизбежно наступает нарушение функции эндокринной и системы, и наоборот
нарушения работы гипоталамо-гипофизарной системы, надпочечников,
щитовидной железы приводят нарушению работы центра голода-насыщения.
Лечение ожирения преследует 3 основные задачи:
Достижение идеальной массы тела, что у больных особенно с крайними степенями ожирения не всегда выполнимо.
Остановка прогрессирующего увеличения массы тела и предупреждение тем самым развития сопутствующих заболеваний.
Снижение
избыточной массы тела минимум на 10%, что уже приводит к статистически
достоверному улучшению течения сопутствующих заболеваний, и снижает риск
их развития.
Объективно оценить, насколько далеко зашел процесс,
помогает индекс массы тела – это величина, которая показывает степень
избыточного веса. Его рассчитывают путем деления веса в килограммах на
квадрат роста в метрах. При нормальной массе ИМТ составляет 18,5 — 24,5,
об избыточном весе говорят, когда это цифры 25,0 — 29,9. Смело можете
ставить себе диагноз ожирения I степени, если ИМТ= 30,0 — 34,9, ожирения
II степени, если 35,0 — 39,9, и III степени при ИМТ >40,0.
Без
сомнения, очень важно знать, почему возникает ожирение, и как
предотвратить его появление, но большинство из нас спохватывается, когда
проблема уже налицо и пришла пора приступить непосредственно к лечению
ожирения. И если многим это становится понятно, только глядя в зеркало,
которое отражает далеко не те формы, которые вам хотелось бы иметь, то
врачи не устают подчеркивать, что без полноценного лечения ожирения вы
обрекаете себя на развитие тяжелых хронических болезней и, в конечном
счете, уменьшение продолжительности жизни, которая, согласитесь, может
быть такой прекрасной.
По данным последних лет ясно, что
консервативное лечение ожирения оказывается успешным, только если
проводится комплексно – специальные диеты, лекарства и регулярные
физические упражнения. Но и в случае хорошего результата всегда остается
вероятность того, что вес вернется к исходному уровню, если вы не
выдержите и дадите себе поблажку, ибо сразу нужно принять за правило,
что лечение ожирения – это пожизненный процесс.
Если говорить о
питании, то на страницах сайта в изобилии представлены разнообразнейшие
диеты, которые можно выбрать на свой вкус. Хотя лечение ожирения
подразумевает больше переход к рациональному сбалансированному питанию, о
чем вы тоже можете узнать, оставаясь у нас в гостях. Сейчас же
напомним, что рациональное питание в идеале должно быть гипокалорийным,
учитывающим ваши личные пищевые пристрастия, чтобы поглощение пищи не
стало для вас мукой, а также возраст, пол и физическую активность.
Большой
интерес вызывают, конечно, лекарства для лечения ожирения, которые,
правда, нужно принимать очень длительно и если немедикаментозное лечение
ожирения не приносит плодов. Они способствуют вполне определенному
снижению веса, темп которого выравнивается обычно к шестому месяцу
лечения, но после года лечения эффективность их, по данным последних
исследований, начинает падать. Что же может нам предложить современная
фармакология для лечения ожирения? Такие известные препараты, как
сибутрамин и фентермин обладают способностью снижать аппетит. При их
приеме быстро возникает чувство насыщения, что заставляет вас спокойно
отказываться от поглощения дополнительной порции пищи. Другие средства –
орлистат или ксеникал, не дают всасываться жирам, поступающим с пищей,
связываясь в желудке с разнообразными липазами, благодаря которым жиры и
расщепляются, в результате чего до 30% жиров выводится со стулом. Но и
этого недостаточно, если лишний вес составляет 45 и более килограмм,
остается последнее средство – бариатрическая хирургия.
Результаты
оперативного лечения ожирения впечатляют – по данным американских
врачей, пациенты за 10 лет теряют от 20 до 80 кг и поддерживают вес на
достигнутом уровне, что хорошо сказывается и на течении сопутствующих
заболеваний – сахарного диабета, артериальной гипертензии, липидных
нарушений обмена и т.д. Поэтому для кого-то лучший вариант лечения
ожирения – бариатрическая хирургия, которая как раз помогает людям с
избыточным весом. И для многих, особенно при наличии серьезных
заболеваний, это единственный способ улучшить здоровье и продлить жизнь,
и сделать это окончательно и бесповоротно.
Перед тем, как принять
решение об операции, нужно узнать, какой у вас индекс массы тела. Если
цифра ИМТ оказывается больше 40, значит пришла пора заняться лечением
ожирения путем проведения бариатрической операции. Если же вы уже
страдаете от таких неприятных вещей, как повышенное давление, сахарный
диабет, проблемы с суставами или варикозное расширение вен, то решаться
на хирургическое вмешательство нужно и при ИМТ, равном 35.
В
арсенале бариатрической хирургии имеется на сегодняшний день несколько
проверенных операций, общий смысл которых – значительно уменьшить объем
поступаемой в желудок пищи, раз уж вы не можете это сделать усилием
воли! Например, бандажирование желудка. Это когда вам на желудок
накладывают силиконовое колечко, из-за чего он становится похож по форме
на песочные часы. А верхний отдел такого желудка способно заполнить и
растянуть даже малое количество пищи, и именно растяжение верхней части
желудка сообщает головному мозгу, что вы насытились и больше есть не
хотите. Сами понимаете, если так происходит постоянно, пищи вы съедаете
не в пример меньше, чем до операции – все гениальное просто! И так
потихоньку, за год-полтора, теряете 50-70 % лишнего веса.
Желудок
еще можно шунтировать – тоже создается «малый желудочек» в верхней части
обычного, объемом где-то 50 мл, к которому еще и подшивается петля
тонкой кишки. В результате половина пищи проходит не обычным путем, а
сразу в кишечник, то есть питательные вещества из нее не всасываются.
Итог очень впечатляющий – 70-100% избыточной массы тела за год-полтора.
Более
щадящая операция для лечения ожирения – введение внутрижелудочного
баллона, как временной меры. Представляет собой силиконовый шар, который
уже в желудке заполняется жидкостью. Постоянно делая вид, что он и есть
пища, баллон имитирует чувство сытости, воздействуя на стенки органа. К
сожалению, уже через 6 месяцев баллон нужно удалить, чтобы не было
осложнений, но за это время можно потерять 10 — 15 килограммов. Но и
получив столь долгожданный результат, вы не должны забывать, что болезнь
никуда не уйдет, если вы возьметесь за ум и не сформируете, наконец,
правильный режим питания и достаточной физической активности, которые
являются важнейшими составляющими здорового образа жизни
Развитие
ожирения у любого человека возникает тогда, когда потребление энергии
превышает ее затраты в течение длительного времени. Очень
незначительное, но постоянное избыточное потребление энергии приводит к
выраженному накоплению жировой ткани в организме.
Ожирение может
постичь даже не предрасположенных к нему людей – слишком велико влияние
современной цивилизации потребления на всех и каждого. Развитие
ожирения стало одной из основных проблем здравоохранения в большинстве
промышленно развитых стран. Болезненное, (морбидное) ожирение
приобретает поистине угрожающие масштабы из за неизбежного развития
тяжелейших сопутствующих заболеваний, находящихся в прямой
причинно-следственной связи с избыточной массой тела.
Нарушение обмена веществ при ожирении
Метаболический
синдром, также известный как синдром инсулинорезистентности или синдром
Х, представляет собой совокупность обменных нарушений у лиц с
определенным конституциональным типом. Метаболический синдром является
фактором риска развития ишемической болезни сердца. Синдром включает
следующие признаки:
внутрибрюшное ожирение
инсулинорезистентность (увеличение уровня глюкозы и инсулина в крови натощак)
сахарный диабет 2 типа
дислипидемия
(повышенный уровень триглицеридов в крови, низкая концентрация в
сыворотке крови липопротеинов высокой плотности (ЛПВП)
артериальная гипертония.
В
последнее время были определены и другие метаболические (обменные)
нарушения (при внутрибрюшном ожирении), которые могут служить фактором
риска развития ишемической болезни сердца. Ожирение само по себе не
является необходимым условием развития метаболического синдрома. Имеется
в виду метаболическое «ожирение» с преимущественно внутрибрюшным
накоплением жировой ткани даже у людей с нормальным весом.
Существует
гипотеза, что основным механизмом развития метаболического синдрома
является инсулинорезистентность. Однако в исследовании Framingham
Offspring Study с участием пациентов, не страдавших диабетом, было
выявлено, что инсулинорезистентность может быть не единственным
фактором, приводящим к развитию метаболического синдрома, и существуют
еще и другие независимые физиологические процессы, участвующие в
формировании данной патологии. Хотя абдоминальное (внутрибрюшное)
ожирение часто сочетается с инсулинорезистентностью, до конца не ясно,
накопление ли внутриорганной жировой ткани (в области сальника и
брыжейки кишечника) или подкожной жировой клетчатки в области живота
связано с развитием инсулинорезистентности.
К тому же объем одного
депо тесно связан с объемом другого, в связи с чем, сложно определить
роль каждого в изменении чувствительности к инсулину. Кроме того, не
известно, накопление висцеральной или подкожной абдоминальной жировой
ткани участвует в развитии метаболического синдрома или это вообще
характерно для людей с повышенным риском развития обменных осложнений
ожирения.
Сахарный диабет 2 типа
Вероятно, увеличение
распространения сахарного диабета 2 типа на 25% в мире за последние 20
лет связано со значительным распространением ожирения. Увеличение
Индекса массы тела (ИМТ), абдоминальный (внутрибрюшной) тип накопления
жировой ткани — серьезные факторы риска развития сахарного диабета 2
типа.
Данные национального института здоровья свидетельствуют, что
более 2/3 мужчин и женщин в США, страдающих диабетом 2 типа, имеют ИМТ
> 27 кг/м2.
Более того, риск диабета возрастает линейно в
зависимости от величины ИМТ. Среди людей, имеющих ИМТ 25,0—29,9 кг/м2
(избыточная масса тела), 30,0—34,9 кг/м2 (ожирение I степени) и > 35
кг/м2 (ожирение II/III степени), распространенность сахарного диабета 2
типа была 2%, 8% и 13% соответственно.
Данные Nurses Health Study
показали, что риск развития диабета начинает возрастать даже у женщин с
нормальным весом, когда ИМТ превышает 22 кг/м2. С увеличением
внутрибрюшной жировой массы, окружности талии и соотношения окружности
талии к окружности бедра риск диабета увеличивается при любом значении
ИМТ.
Прирост массы тела в молодом возрасте также увеличивает риск
развития диабета. Так у мужчин и женщин 35 — 60 лет, которые в возрасте
18–20 лет прибавили в весе от 5 до 10 кг, риск развития диабета был в 3
раза выше, чем у тех, у которых вес изменялся в пределах 2 кг.
Дислипидемия
Ожирение,
особенно абдоминальное (внутрибрюшное), ассоциировано с повышением
уровня триглицеридов, низкой концентрацией холестерина высокой плотности
и увеличением концентрации холестерина низкой плотности, который
является основным фактором способствующим формированию атеросклероза.
Большинство
данных указывает на то, что при избыточной массе тела и ожирении
увеличивается концентрация в сыворотке общего холестерина и холестерина
низкой плотности, однако связанные с цифрами ИМТ различия в
концентрациях общего холестерина и холестерина низкой плотности более
выражены у людей с меньшим весом, и с возрастом они уменьшаются.
Данные
Национального института здоровья США показали, что с увеличением цифр
ИМТ у мужчин прогрессивно увеличивается концентрация холестерина в крови
(содержание общего холестерина > 240 мг/дл или 6,21 ммоль/л), в то
время как у женщин распространенность увеличенного уровня холестерина
была самой высокой при ИМТ 25 кг/м2 и 27 кг/м2, и не увеличивалась с
дальнейшим ростом цифр ИМТ.
Патологическое изменение концентрации
липидов (жиров) в сыворотке при ожирении является клинически важным
показателем, так как связано с увеличением риска развития ишемической
болезни сердца.
Риск осложнений сердечно-сосудистых заболеваний при ожирении
Ишемическая
болезнь сердца (ИБС) Пациенты, страдающие преимущественно абдоминальной
(внутрибрюшной) формой ожирения, и те, у кого отмечался рост массы тела
в молодом возрасте, подвержены большему риску возникновения ишемической
болезни сердца (ИБС).
Риск ИБС начинает расти уже при «нормальном»
индексом массы тела (ИМТ) (23 кг/м2 у мужчин и 22 кг/м2 у женщин). А
наличие абдоминального ожирения увеличивает риск ИБС при любом значении
ИМТ. Действительно, в исследовании Nurse Health Study было выявлено, что
у женщин с более низким ИМТ, но большим значением отношения окружности
талии к окружности бедра, риск возникновения инфаркта миокарда (в том
числе с летальным исходом) был выше, чем у женщин с большим ИМТ, но
меньшим значением отношения окружности талии к окружности бедра.
Прибавка
в весе 5 кг и более после 18 лет также увеличивает риск возникновения
инфаркта миокарда. Большую роль в увеличении частоты ИБС при ожирении
играют такие факторы риска, как артериальная гипертония, дислипидемия,
изменение толерантности к глюкозе или диабет, а также метаболический
синдром.
В эпидемиологических исследованиях сложнее изучить влияние
самого ожирения на риск ИБС, возможно, потому что необходимо длительное
время для наблюдения больных, выявления роли других факторов риска,
влияющих на изменение массы тела (например, курение), и изучение влияния
типа распределения жировой ткани.
Тем не менее, в нескольких
продолжительных эпидемиологических исследованиях было показано, что
лишний вес и ожирение увеличивают риск ИБС даже после коррекции других
известных факторов риска. В связи с этим, Американская кардиологическая
ассоциация недавно включила ожирение в перечень важнейших факторов риска
ишемической болезни сердца и разработала рекомендации по нормализации
веса.
Цереброваскулярные и тромбоэмболические осложнения
Как у
женщин, так и у мужчин лишний вес и ожирение увеличивают риск
возникновения ишемического инсульта. Риск развития инсульта (в том числе
и с летальным исходом) у больных ожирением прогрессивно растет с
увеличением значений индекса массы тела (ИМТ) и почти в два раза выше,
чем у худощавых людей.
Ожирение, особенно абдоминальное, также
увеличивает риск венозного застоя, тромбоза глубоких вен и тромбоэмболии
(закупорки оторвавшимся тромбом) легочной артерии. Болезнь вен нижних
конечностей может развиваться вследствие увеличения внутрибрюшного
давления и патологии свертывающей системы крови, а также увеличения
выброса медиаторов воспаления (биологически активных веществ
сопровождающих воспаление) при абдоминальном (внутрибрюшном) ожирении.
Недавние
исследования показали, что у лиц пожилого возраста с ИМТ 25 кг/м2 и
более, после перенесенной операции протезирования тазобедренного
сустава, риск последующей госпитализации по поводу тромбоэмболических
осложнений (тромбоз глубоких вен или тромбоэмболия легочной артерии) был
в 2,5 раза больше.
Артериальная гипертония (АГ)
Связь между
значением индекса массы тела (ИМТ) и возникновением артериальной
гипертонии была подтверждена в крупных эпидемиологических исследованиях.
Данные этих исследований продемонстрировали, что (с учетом возраста)
распространенность АГ среди женщин и мужчин, страдающих ожирением, в 2,5
раза выше (38% и 42% соответственно), чем у худых людей (15% как среди
мужчин, так и женщин).
Существенным фактором риска развития
артериальной гипертонии также является абдоминальный (внутрибрюшной) тип
ожирения, в некоторых исследованиях указанный который может быть даже
более важным, чем ИМТ. В работе Framingham Study было обнаружено, что АД
увеличивается на 6,5 мм рт. ст. на каждые 10% увеличения массы тела.
Осложнение гастроэнтерологических заболеваний при ожирении
Гастроэзофагеальная рефлюксная болезнь (ГЭРБ)
Заключается
в развитии воспаления в нижней трети пищевода вследствие недостаточно
плотного смыкания круговой мышцы (сфинктера) расположенного между
пищеводом и желудком. Во многих крупных эпидемиологических исследованиях
было обнаружено, что симптомы ГЭРБ и у худых людей, и у пациентов,
страдающих ожирением, практически одинаковы.
Однако, пока не ясно,
действительно ли ожирение вызывает развитие рефлюкса (заброса кислого
желудочного содержимого в пищевод) или просто часто сочетается с ГЭРБ.
Ввиду наличия противоречивых данных, неизвестно, существует ли
взаимосвязь между ожирением и известными факторами, предрасполагающими к
развитию ГЭРБ (ослабление тонуса нижнего пищеводного сфинктера и
изменение рН в кислую сторону в нижей части пищевода).
Тем ни менее
многие исследования подтверждают влияние ожирения на появление симптомов
пищеводного рефлюкса. Хотя повышение внутрибрюшного давления, вызванное
увеличенными размерами живота, у людей, страдающих ожирением, может
предрасполагать к возникновению рефлюкса, неизвестно, проводились ли
какие-либо контролируемые рандомизированные (статистически достоверные)
исследования по оценке влияния снижения веса на уменьшение симптомов
рефлюкса. Тот факт, что даже умеренное снижение веса приводит к
значительному улучшению состояния пациентов, говорит скорее всего за то,
что благоприятный клинический эффект достигается больше благодаря
диете, нежели собственно снижению массы тела.
Исследования с
участием пациентов с ожирением первой степени и симптомами ГЭРБ
показали, что снижение веса не привело к улучшению состояния или
изменению показателей пищеводной рН при 24–часовом контроле.
Характерно,
что симптомы ГЭРБ всегда исчезают сразу после хирургического
вмешательства по поводу ожирения, еще до того, как существенно снизится
масса тела. Это говорит о том, что исключение самого заброса кислого
содержимого или желчи в пищевод, а не снижение веса, улучшает состояние
пациентов. Выраженный гастроэзофагеальный рефлюкс может появиться после
вертикальной гастропластики. Хотя в других исследованиях показано, что
вертикальная бандажированная гастропластика и бандажирование желудка,
при которых увеличивается сопротивление прохождению пищи через
сформированное сужение (см. хирургические методы лечения), не изменяет
тонус нижнего пищеводного сфинктера и не увеличивает число эпизодов
рефлюкса.
Болезни желчного пузыря
Ожирение является серьезным фактором риска возникновения заболеваний желчного пузыря, особенно у женщин.
Риск
появления камней в желчном пузыре увеличивается с ростом индекса массы
тела (ИМТ). Данные исследования Nurses Health Study продемонстрировали,
что женщины, страдающие ожирением (ИМТ > 30 кг/м2) имеют вдвое
больший риск (а женщины с тяжелой формой ожирения и ИМТ > 45 кг/м2 — в
7 раз больший риск) появления камней в желчном пузыре по сравнению с
худощавыми женщинами (ИМТ < 24 кг/м2). Ежегодная заболеваемость
составляет 1% среди женщин с ИМТ > 30 кг/м2 и 2% среди женщин с ИМТ
> 45 кг/м2.
У мужчин, страдающих ожирением, риск развития желчекаменной болезни ниже, чем у женщин.
Вероятность
образования камней в желчном пузыре растет и при снижении веса в связи с
увеличением концентрации холестерина в желчи, образованием кристаллов
холестерина и снижением сократительной функции желчного пузыря.
Образование
«новых» камней происходит примерно у 25-35% людей с ожирением, которые
быстро сбросили вес благодаря применению низко–жировой или очень
низкокалорийной диеты, а также хирургического вмешательства на желудке.
Риск образования камней в желчном пузыре увеличивается, когда скорость
снижения веса достигает 1,5 кг (~ 1,5% массы тела) в неделю.
Содержание
жиров в пище также влияет на образование камней в желчном пузыре. Так,
поступление с пищей 4 г жиров слабо стимулирует опорожнение желчного
пузыря, в то время как поступление 10 г жиров способствует его
максимальной сократительной активности. Обнаружено, что увеличение
потребления жиров при очень низкокалорийной диете (< 600 ккал в день)
предохраняет от образования камней.
Риск образования камней во
время диеты становится значительно ниже (0–17%) у тех пациентов, которые
придерживаются низкокалорийной, а не очень низкокалорийной и
низко–жировой, диеты (> 800 ккал в день), содержащей 15–30 г жиров в
день. Для уменьшения риска образования камней в желчном пузыре не
обязательно увеличивать содержание жиров в пище во время диеты.
Риск
образование камней, как при использовании очень низкокалорийной диеты,
так и после хирургического вмешательства можно значительно снизить,
добавив к лечению урсодезоксихолиевую кислоту. Назначение
урсодезоксихолиевой кислоты в дозе 600 мг в день обеспечивает наилучшую
профилактику образования камней в желчном пузыре у тех пациентов,
которые хотели бы достичь быстрого снижения веса.
Панкреатит
Хотя
было бы логичным предположить, что пациенты, страдающие ожирением,
имеют более высокий риск развития панкреатита в связи с наличием камней в
желчном пузыре, изучению этого вопроса было посвящено лишь небольшое
число исследований.
Однако статистически достоверно установлено, что
панкреатит любого происхождения у пациентов с ожирением имеет более
плохой прогноз, чем у пациентов с нормальным весом.
Большое число
исследований свидетельствует о том, что у пациентов, имеющих лишний вес
или ожирение, высок риск развития местных и общих осложнений
панкреатита. Увеличен риск развития самой тяжелой стадии этого
заболевания — панкреонекроза.
Болезни печени
Ожирение приводит к
нарушению работы печени, что проявляется в увеличении ее размеров,
увеличением биохимических печеночных показателей и изменениями на
клеточном уровне (крупноячеистый стеатоз, жировой гепатоз, фиброз и
цирроз).
Хотя эта патология представлена в обзорах, как ряд частных
случаев, вероятнее всего, такого рода нарушения можно отнести к
признакам заболевания, известного как неалкогольная жировая дистрофия
печени. В связи с малочисленностью данных, широта распространения данной
патологии у пациентов с ожирением не известна.
Наиболее характерным
признаком является увеличение печеночных ферментов
(аланинаминотрансферазы — АЛТ и аспартатаминотрансферазы — АСТ). Но
обычно эти цифры не превышают двукратного значения верхней границы
нормы. Кроме того, уровень роста печеночных ферментов не соответствует
тяжести гистологических изменений. Диета сама по себе может быть
причиной временного увеличения концентрации печеночных ферментов в
течение первых 6 недель снижения веса.
Ретроспективный анализ
образцов печеночной ткани, взятых на исследование у пациентов, имевших
лишний вес или страдавших ожирением, показал, что у 30% пациентов имелся
фиброз печеночной соединительной ткани, причем у трети из них (10% из
всей группы) был обнаружен скрытый цирроз печени. Кроме того, ожирением
страдают многие пациенты, у которых обнаруживаются признаки жирового
перерождения печени неалкогольного происхождения. От 40% до 100%
пациентов с неалкогольным стеатогепатитом (НСГ), по совокупным данным
нескольких исследований, имеют ожирение.
По данным аутопсии
(посмертного вскрытия) у пациентов, страдавших ожирением, в ~75% случаев
встречается стеатоз, в ~20% — стеатогепатит и в ~2% — цирроз печени.
Хотя
клинические, лабораторные и клеточные признаки неалкогольного жирового
поражения печени уже определены, природа и патогенез этого заболевания
до сих пор не совсем понятны. У многих заболевание протекает
бессимптомно, или больные жалуются на повышенную утомляемость,
недомогание или чувство дискомфорта в области живота.
Увеличение
размеров печени отмечается у 75% больных. Отношение АСТ/АЛТ у больных
обычно меньше единицы, в отличие от значения этого показателя у больных
алкогольным стеатогепатитом.
При наблюдении пациентов в течение 1–7
лет было отмечено прогрессирование болезни печени у 40% больных, а у 10%
развился цирроз. Кроме того, у большинства пациентов с простым
стеатозом, болезнь принимала доброкачественное течение, в то время как
стеатогепатит, фиброз и цирроз часто приводили к развитию осложнений и
более тяжелому течению заболевания. Хотя, в конечном счете, лишь у
небольшого числа больных неалкогольным жировым поражением печени
развивается цирроз, в странах с широким распространением ожирения эта
патология становится одной из основных причин цирроза печени.
К тому
же ожирение увеличивает риск развития фиброза и цирроза у больных с
алкогольным поражением печени и гепатитом С. Не до конца понятно, почему
у людей, страдающих ожирением, развивается неалкогольное жировое
перерождение печени. Есть основания полагать, что развитие этой
патологии часто ассоциировано с внутрибрюшным ожирением (определяемым по
окружности талии), инсулинорезистентностью (увеличение уровня глюкозы
натощак и инсулина в крови), диабетом, повышенным содержанием
триглицеридов в крови, низкой концентрацией в сыворотке липопротеидов
высокой плотности (ЛПВП) и артериальной гипертензией. Существует
гипотеза, что, развитие данного заболевания связано с 2 и более
повреждающими воздействиями на печень.
В первую очередь это стеатоз,
причиной которого чаще всего является изменение липидного обмена
вследствие ожирения, а именно увеличение расщепления триглицеридов
жировой ткани, что в свою очередь увеличивает поступление свободных
жирных кислот в печень.
Во–вторых, перекисное окисление липидов в
печени и высвобождение цитокинов может оказывать прямое повреждающее
действие на клетки печени и способствовать развитию воспаления и
фиброза.
Хотя снижение веса является типичной рекомендацией для
пациентов с ожирением и неалкогольным жировым поражением печени, пока не
известно, воздействует ли такая терапия на природу заболевания.
Постепенное снижение веса на 10% и более может корректировать уровень
печеночных ферментов и способствовать уменьшению размеров печени,
содержания жира в печеночной ткани и ослаблению проявлений
стеатогепатита. Но быстрое снижение веса при терапии с помощью очень
низкокалорийной диеты или голодания может спровоцировать воспаление.
Ожирение увеличивает риск развития рака
Прослеживается
взаимосвязь наличия лишнего веса и ожирения с увеличением риска
развития рака пищевода, желчного пузыря, поджелудочной железы, молочной
железы, почек, матки, шейки матки и предстательной железы.
Во многих
эпидемиологических исследованиях была выявлена строгая взаимосвязь
между индексом массы тела (ИМТ) и развитием рака толстой кишки и у
мужчин, и у женщин. Эта взаимосвязь еще усиливается в отношении рака
дистальных частей кишечника (толстой кишки) и в отношении больных с
указанием на развитие подобного рака у родственников.
Ожирение и лишний вес в возрасте 18 лет увеличивает риск смертности от рака молочной железы и эндометрия.
Однако
у женщин с ожирением в постменопаузе риск рака молочной железы
перестает расти, что, видимо, связано с защитным действием увеличения
веса у женщин в этом возрасте.
Однако сложно определить, увеличение
риска развития рака связано с ожирением как таковым или еще и с
употреблением высококалорийной и содержащей много жиров пищи, поскольку
оба эти фактора часто сочетаются.
Поражение суставов при ожирении
Подагра
Подагра
— заболевание, характеризующееся поражением суставов вследствие
отложения кристаллов мочевой кислоты. С помощью продолжительных и
перекрестных исследований было установлено, что ожирение влияет на
развитие повышенного содержания мочевой кислоты в крови (гиперурикемия) и
подагры.
Более того, гиперурикемия развивается у больных с
абдоминальным типом ожирения и метаболическим синдромом, а наличие
инсулинорезистентности снижает выведение мочевой кислоты почками.
Остеоартрит
Лишний
вес и ожирение увеличивают риск развития остеоартрита (воспления)
суставов, несущих повышенную нагрузку, чаще коленных, поскольку коленные
суставы в период активности человека испытывают большую нагрузку, чем
тазобедренные.
Чаще остеоартриты возникают у женщин, даже если они
имеют незначительный избыток массы тела. Исследования среди пар
близнецов показали, что если один из близнецов страдает клинически
выраженным или асимптомным остеоартритом нижних конечностей, то его вес,
как правило, на 3–5 кг выше, чем у другого близнеца.
Данные
исследований также показали, что ожирение участвует в патогенезе
(развитии) остеоартрита. В некоторых научных работах с ожирением
связывается и возникновение остеоартрита суставов рук, что позволяет
предположить и существование других механизмов возникновения заболевания
суставов при ожирении, помимо увеличения нагрузки
Психические расстройства при ожирении
У
примерно 20-30% пациентов с ожирением, которым не удалось снизить свой
вес в различных клиниках, развивается депрессия и другие психические
нарушения.
Однако явного доказательства, что у больных с ожирением
чаще, чем у людей с нормальным весом, возникают психические или
эмоциональные нарушения (при произвольной выборке в популяции) [253], не
существует. У женщин такие нарушения возникают чаще, чем у мужчин.
Возможно, это связано с давлением общественного мнения, предписывающего
женщине быть стройной.
Некоторые поведенческие отклонения могут
способствовать возникновению ожирения. Булемия, т.е. употребление
большого количества пищи за короткий промежуток времени, обычно
сопровождается ощущением потери контроля над собой и чувством вины. Этот
диагноз можно поставить в случае повторения таких эпизодов, по меньшей
мере, дважды в месяц в течение 6 и более месяцев, и при этом переедание
не сопровождается последующим употреблением слабительных с целью
предотвращения увеличения массы тела.
Около 10-15% людей, имеющих
ожирение, страдают булемией, в то время как в популяции этот показатель
равен 2%. Пациенты с данным заболеванием, которые включаются в программу
по снижению веса, обычно имеют больший вес и хуже поддаются терапии,
чем те пациенты с ожирением, у кого эта патология отсутствует.
Фармакологическая
и поведенческая коррекция этого нарушения может уменьшить количество
эпизодов переедания, но, как ни странно, часто не приводит к снижению
веса.
Другое отклонение, так называемый синдром ночного приема пищи,
также связано с ожирением, но встречается реже, чем булемия.
Употребление большего количества пищи в вечернее время приводит к более
позднему отходу ко сну, частым ночным пробуждениям, обычно
сопровождающимся приемом пищи, и утренней анорексии.
На
сегодняшний день существуют реальные достижения в лечении ожирения,
которые эффективно применяются и касаются они в основном новых
лекарственных препаратов и хирургических методов борьбы с лишним весом.
Базовые методы лечения ожирения – диета и физическая нагрузка уже имеют большую историю.
Диетотерапия
является основным методом снижения избыточного веса. Это связано с тем,
что для большинства людей с лишним весом проще уменьшить количество
потребляемой пищи, чем увеличить физическую активность.
Большинство
диет основано либо на снижении количества потребляемых калорий, либо на
изменении состава пищи. Однако снижению веса способствует именно
уменьшение калорийности, а не изменение состава пищи. На сегодняшний
день существует огромное количество диет, многие из которых составлены,
что называется, «с потолка». А между тем, диетическое питание оказывает
значительное влияние на организм, обмен веществ и Ваше самочувствие.
Поэтому мы настоятельно рекомендуем Вам: если Вы все же решили соблюдать
диету, то в первую очередь следует посоветоваться с врачом.
Не
занимайтесь самолечением! Некоторые диеты имеют свои противопоказания,
для выявления которых необходимо обследование. Кроме того, врач может
подобрать Вам ту диету, которая будет максимально эффективна и безопасна
именно для Вас.
Физические упражнения поддерживают здоровье,
предохраняют от заболеваний и, чему все больше свидетельств, замедляют
процесс старения. При выполнении физических упражнений в кору головного
мозга поступает огромное количество сигналов (импульсов) от мышц,
сухожилий, связок, суставов, кожи, глаз, ушей, органов равновесия, а
также сердца, сосудов, легких и других органов, обеспечивающих
двигательный акт. В случае уменьшения числа импульсов, идущих от мышц к
мозгу, нарушается нервная деятельность, работоспособность падает.
Упражнения
полезны в любом возрасте, поскольку обычная дневная активность редко
может обеспечить достаточную физическую нагрузку, любой, желающий
насладиться эффектом физических упражнений, должен предпринять для этого
специальные усилия.
Усвоив одну единственную здоровую привычку, вы
сможете снизить вес, уменьшить риск заболеваемости, улучшить сердечную
деятельность, смягчить проявления депрессии и т.д. Многие думают, что
физические упражнения — это из жизни атлетов. Некоторые беспокоятся, что
их будут критиковать за неправильное выполнение движений. На самом деле
мы боимся того, что может быть для нас настоящим спасением. А
упражнений существует великое множество, нужно выбрать те, что подходят
именно Вам. Нужно просто очень сильно хотеть, стремиться и верить в
себя.
Как люди различного возраста могут достичь рекомендованного уровня физической активности
Возраст и род занятий Формы физической активности
Дети младшего школьного возраста
Ходьба пешком в школу и домой.
Физкультурные
занятия и подвижные игры в школе (на переменах, на уроках
физкультуры и в секциях) 3-4 раза в неделю подвижные игры после школы.
выходные дни; более длительные пешие прогулки, посещение парка или бассейна, прогулки на велосипеде
Подростки Ходьба пешком (или езда на велосипеде) в школу и домой.
По будним дням 3-4 организованных или самостоятельных занятий спортом или другими формами физической активности
В выходные дни; пешие и велосипедные прогулки, плавание, другие спортивные занятия.
Студенты Ежедневная ходьба пешком (или езда на велосипеде) в учебное заведение и домой.
Использование любой возможности физической нагрузки; ходьба по лестнице (вместо лифта), различный ручной труд.
По будним дням 2-3 раза в неделю организованные занятия спортом или физические упражнения, посещение спортзала или бассейна.
В выходные дни; более длительные пешие или велосипедные прогулки, плавание, другие спортивные занятия.
Взрослые,
работающие по найму Ходьба пешком или езда на велосипеде на работу.
Использование любой возможности физической нагрузки: ходьба по лестнице
(вместо лифта), различный ручной труд.
По будним дням 2-3 раза занятия спортом, упражнения в спортзале или посещения бассейна.
В
выходные дни: более длительные пешие или велосипедные прогулки,
плавание, занятия спортом, ремонтные работы по дому, работа в саду.
Взрослые, работающие на дому
Ежедневные прогулки, уход за садом или ремонтные работы по дому.
Использование любой возможности физической нагрузки: ходьба по лестнице
(вместо лифта), различный ручной труд.
По будним дням иногда занятия спортом, упражнения в спортзале или посещение бассейна.
В выходные дни: более длительные пешие или велосипедные прогулки, занятия спортом.
Взрослые,
неработающие Ежедневные прогулки, уход за садом, ремонтные работы по
дому. Использование любой возможности физической нагрузки: ходьба по
лестнице (вместо лифта), различный ручной труд.
В выходные дни: более длительные пешие и велосипедные прогулки, плавание, другие занятия спортом.
Время от времени занятия спортом, упражнении в спортзале или посещение бассейна.
Пенсионеры
Ежедневные пешие или велосипедные прогулки, уход за садом или ремонтные
работы по дому. Использование любой возможности физической нагрузки:
ходьба по лестнице (вместо лифта), различный ручной труд.
По выходным дням: более длительные пешие прогулки, езда на велосипеде или плавание.
Более
новые методы, составляющие, на сей день, последние достижения в лечения
ожирения, не требуют больших временных затрат и усилий воли, они
помогают как бы извне, и уже поэтому гарантируют выраженный и
длительный эффект, и тем существенно повышают качество жизни. Впрочем,
медикаментозные препараты для лечения ожирения известны не первый год.
Это в основном лекарства центрального действия, подавляющие аппетит.
А
вот к последним достижениям в лечении ожирения можно отнести новые
разработки, обладающие выраженным несистемным действием. Они имеют более
высокую эффективность и безопасность, поэтому могут применяться более
длительное время. Действуют они следующим образом — блокируют в желудке и
кишечнике фермент липазу, в связи с чем расщепления жира не происходит,
и он проходит через кишечник, не всасываясь. Вследствие этого большой
процент триглицеридов (содержащих жирные кислоты) и холестерина просто
выводится из организма, что позволяет создать существенный дефицит
калорий, особенно на фоне применения низкокалорийной диеты.
Вместе с
избавлением, и существенным, от лишних килограммов снижается риск и
сердечно-сосудистых заболеваний, нормализуется артериальное давление и
углеводный обмен. Нужно ли говорить, что и самочувствие в результате
такого лечения только улучшается! Все замечательно, но, все же,
высмотрев в аптеке подобный препарат, не забудьте проконсультироваться с
лечащим врачом, так как даже самые лучшие препараты имеют
противопоказания к применению.
Использование новых хирургических
методов, также составляющих одно из последних достижений в лечении
ожирения, гарантирует более существенный и быстрый, и самое главное,
самый стабильный результат, когда ожирение имеет тяжелую стадию. В
настоящее время все методы хирургического лечения ожирения основаны на
двух эффектах. Это или резкое уменьшение объема функционирующего
желудка, или исключение из процесса пищеварения значительного участка
тонкой кишки, что не позволяет полноценно всасываться большому
количеству питательных веществ.
И в том, и в другом случае результат
основан на создании значительного дефицита калорий, чего не удается
добиться большинству людей применением различных диет. Здесь же вы
неуклонно теряете лишние килограммы, так как энергетический дефицит
существует помимо любых ваших усилий. Выраженный эффект бариатрических
операций поддерживается и тем, что наряду с потерей килограммов,
уменьшаются и проявления большинства сопутствующих заболеваний.
Но
последнее достижение лечений ожирения – временная установка
внутрижелудочного баллона, которая позволила применять эффективное
хирургическое вмешательство даже при невысоких степенях ожирения, тогда
как раньше операции проводились лишь при очень сильном ожирении и полном
отсутствии эффекта от консервативной терапии. Суть манипуляции
заключается в том, что в желудок заводится зонд, нижний конец которого
соединен с плотно упакованным силиконовым шаром. При введении через зонд
физиологического раствора шар расправляется до объема 400-800 мл,
заполняя значительную часть полости желудка. Шар устанавливается на срок
до 6 мес., и затем удаляется. При необходимости процедуру через
определенное время можно повторить.
Также к международным стандартам
хирургического лечения ожирения относится операция наложения
желудочного бандажа, который изменяет форму желудка в виде «песочных
часов» для создания «малого желудочка» с очень небольшим объемом. Пища,
попадая в малый «верхний» отдел желудка, благодаря наличию узкого
перехода, созданного внешним бандажом, задерживается в нем, воздействуя
при этом на рецепторы, находящиеся в большом количестве в области дна,
которые активизируют центр насыщения в коре головного мозга. Находясь в
«малом желудочке» в течение длительного времени, пища помогает пациенту
долго не испытывать чувства голода.
Реже, из-за своего более
сильного эффекта, применяется шунтирование желудка, когда создается
небольшой резервуар в верхней части желудка, способный вместить лишь
совсем маленькое количество (до 30 куб. см) твердой пищи. Далее, с
помощью специального подшивания петли тонкой кишки, создается обходной
путь, по которому съеденная пища минует большую часть желудка и часть
тонкого кишечника. В результате происходит эффективное снижение веса
вследствие уменьшения количества принимаемой пищи и невозможности ее
полноценного всасывания.
Такое вмешательство требует тщательного
наблюдения лечащим врачом, так как в некоторых случаях, вследствие
нарушений всасывания микроэлементов, возможно развитие различных
состояний, связанных с недостаточностью витаминов (в частности B12),
железа, кальция. В таких случаях необходимо принимать пищевые
минеральные и витаминные комплексы, проводить медикаментозную коррекцию
указанных состояний. Поэтому такая операция проводится, как правило, при
очень значительных превышениях веса, когда риск от осложнений,
связанных с ожирением, намного превышает риск операции.
Что и
говорить, хирургия вещь очень серьезная, и выполнение любых операций у
больных ожирением требует самой высокой квалификации хирурга,
современного оснащения клиники, обученного медицинского персонала.
Многое зависит и от самого пациента, который для хорошего результата
должен внести свой вклад четким соблюдением рекомендаций врача после
операции. Только тесное взаимодействие хирурга и пациента позволяет
достичь желаемого результата.
Просто и понятно о хирургии снижения веса.
Как похудеть? Что представляет собой избыточный вес
Как
похудеть? Можно сколько угодно уходить от проблемы веса, считать ее
косметическим дефектом, оправдывать конституциональными особенностями
организма, прятать за шутками вроде: «хорошего человека должно быть
много», от этого ожирение не перестанет быть тяжелейшим заболеванием,
имеющим тенденцию ко всё большему распространению. Этот недуг может
постичь даже не предрасположенных к нему людей — слишком велико влияние
современной цивилизации потребления на всех и каждого. Ожирение
отравляет и значительно укорачивает жизнь многих и многих людей. Для
эффективной борьбы с избыточным весом, прежде всего, нужно четко
уяснить, что это болезнь которую необходимо лечить. Необходимо признать
существование проблемы, перестать стесняться и или наоборот бравировать
своими большими размерами.
Как похудеть? Что такое бариатрическая хирургия?
Раздел хирургии, занимающийся лечением людей с избыточным весом, называется «бариатрическая хирургия».
Если
вы много лет пытались похудеть, испробовали все диеты, лекарства и
другие консервативные способы, если избыточный вес причиняет вам
серьезные проблемы, в том числе проблемы со здоровьем, то можно подумать
о самом сильном и стабильном способе похудения — о хирургическом
лечении.
Почему вообще возникает необходимость делать операции для
снижения веса? Как мы знаем, у многих людей избыточный вес вызывает
крайне серьезные медицинские проблемы. Он сам по себе становится
болезнью. В этом случае снижать вес нужно обязательно, чтобы улучшить
здоровье и продлить жизнь. Никакие другие способы, кроме хирургического,
не дают такого сильного, и главное, стабильного снижения веса.
Известно, что только 1% людей сохраняет сниженный вес после похудения за
счет диеты или таблеток, остальные набирают прежний или еще больший
вес. Только хирургия дает возможность решить эту проблему окончательно.
Поэтому все больше и больше людей в мире делают выбор в пользу
хирургического лечения.
Чтобы ответить на вопрос: «а нужно ли мне
делать операцию по снижению веса?», нужно обязательно определить ваш
Индекс Массы Тела.
Индекс Массы Тела это величина, характеризующая
степень избыточного веса, определяется путем деления веса в килограммах,
на квадрат роста в метрах.
Если ИМТ больше 40, тогда вам можно и нужно делать такую операцию.
В
случае, если у вас имеются такие болезни, как повышенное давление,
сахарный диабет, проблемы с суставами, или варикозное расширение вен,
тогда операцию необходимо делать уже при ИМТ равном 35.
Как похудеть? Бандажирование желудка
Цель
операции заключается в резком уменьшении объема съедаемой пищи, и
соответственно, в снижении количества потребляемых калорий. Эта цель
достигается путем наложения специального силиконового кольца (бандажа)
на верхнюю часть желудка, сразу под пищеводом. Наложенное кольцо придает
желудку форму песочных часов. Совсем небольшое количество пищи, попадая
в малую часть желудка и задерживаясь в ней, вызывает растяжение
желудочных стенок, которое обычно бывает только при переполненном
желудке. Специальные нервы верхней части желудка сигнализируют в мозг:
«Я сыт!!!»
Через год-полтора после этой операции можно потерять от 50 до 70% избыточной массы тела.
Как похудеть? Шунтирование желудка
Операция,
которая сочетает в себе уменьшение желудка и снижение всасывания
питательных веществ — это операция желудочного шунтирования. Суть ее
заключается в том, что создается «малый желудочек» объемом 50 мл путем
пересечения желудка в верхней части, а затем к этому «малому желудочку»
подшивается петля тонкой кишки. При этом пища идет по короткому пути, и
половина -для восстановления проходимости желудочно-кишечного тракта.
Кроме того, кишечник перестраивается таким образом, что пищеварительные
соки поджелудочной железы и желчь встречаются с пищей лишь после
прохождения полутора метров тонкой кишки.
Таким образом, благодаря
уменьшению желудка достигается эффект малого количества съедаемой пищи, а
ее «холостой» транспорт по тонкой кишке приводит к уменьшению
всасывания питательных веществ.
Через год-полтора после этой операции можно потерять от 70 до 100% избыточной массы тела.
Как похудеть? Что такое лапароскопия?
Лапрароскопия
или лапароскопическая хирургия — это способ выполнять хирургические
операции без разреза живота. Вместо разреза хирург делает несколько
проколов, через которые вводит специальные инструменты. Происходящее
внутри пациента хирургическая бригада контролирует посредством
видеосистемы соединенной с оптическим прибором, который также
вставляется в живот через прокол. Благодаря лапароскопической технике
операция становится «малоивазивной», то есть не причиняющей значительной
травмы тканям и органам пациента. Представляете насколько это важно для
людей страдающих от избыточного веса? Молниеносное развитие
бариатрической хирургии в последнее десятилетие связано исключительно с
появлением лапароскопического метода оперирования.
Все, даже самые сложные бариатрические операции мы выполняем Лапароскопически.
Как похудеть? Лечение внутрижелудочным баллоном
В
качестве временной меры для снижения веса мы применяем внутрижелудочный
баллон. Это устройство представляет собой силиконовый шар, заполняемый
жидкостью в объеме ? литра. Заполняя часть внутрижелудочного
пространства, а также, воздействуя на нервные окончания, баллон
имитирует чувство сытости.
Благодаря установке желудочного баллона
пациент может потерять 15 — 20кг. Однако необходимо помнить, что не все
хорошо переносят присутствие крупного инородного тела в желудке. Кроме
того, через 6 месяцев после установки баллона его необходимо удалять во
избежание развития осложнений. При этом, если на место старого не
устанавливается новый баллон, потерянные килограммы могут вернуться
вновь с избытком.
Наши советы:
ешьте три раза в день, включая завтрак;
избегайте
пищи, в которой много сахара и жира, и наоборот, вводите в рацион
крахмалосодержащую пищу — хлеб из пшеничной муки грубого помола,
картофель, рис;
уменьшите порции мясных блюд, даже постное мясо дает много калорий;
переходите на обезжиренные молочные продукты и продукты пониженной жирности;
ешьте медленно, отводите еде полчаса, от добавок откажитесь;
если
вы чувствуете голод в промежутках между завтраком и обедом, обедом и
ужином, установите время для легкой закуски утром и во второй половине
дня;
чтобы закуска действительно была легкой, малокалорийной,
наметьте заранее, что будете есть; идеальны для закусок фрукты и овощи;
ешьте медленно, пусть ваши вторые завтраки и обеды длятся не менее 10
минут;
будьте готовы к тому, что вес будет уменьшаться медленно;
лучше похудеть за год на 5кг, чем сбросить те же килограммы за месяц, а
через два их снова набрать;
ведите «дневник питания и физических нагрузок».
Мезенхимальные дистрофии — это… Что такое Мезенхимальные дистрофии?
Мезенхима́льные дистрофи́и (стромально-сосудистые дистрофии) — нарушения метаболизма, развивающиеся в строме органов.
Классификация
В зависимости от типа веществ, обмен которых нарушен, выделяют три группы мезенхимальных дистрофий:
- Мезенхимальные диспротеинозы (нарушения обмена белков)
- Мезенхимальные липодистрофии (нарушения обмена липидов)
- Мезенхимальные углеводные дистрофии.
К мезенхимальным диспротеинозам относятся (1) мукоидное набухание, (2) фибриноидные изменения, (3) внеклеточный гиалиноз и (4) амилоидоз.
Мезенхимальные липодистрофии
Мезенхимальные липодистрофии (мезенхимальные липидозы) включают (1) нарушения обмена ацилглицеролов (нейтральных жиров), а также (2) холестерола и его эфиров (холестеридов). К ацилглицеролам относятся три класса соединений — моно-, ди- и триглицериды. Нарушения метаболизма нейтральных жиров (ацилглицерозы) носят локальный (местный) и генерализованный (распространённый) характер, сопровождаются накоплением (кумулятивные дистрофии) или истощением (резорбтивные дистрофии) тканевых липидов. Нарушения обмена холестерола и холестеридов (холестерозы) проявляются как недостаточностью этих веществ в организме (болезнь Бе́ссена—Ко́рнцвейга), так и избыточной концентрацией их в некоторых тканях (атеросклероз, ксантоматоз, липоидная дуга роговицы). В качестве синонима для обозначения холестерозов иногда используется термин холестериновый диатез.
I. Ацилглицерозы
- Генерализованные кумулятивные ацилглицерозы (ожирение, наследственные гиперацилглицеролемии)
- Генерализованные резорбтивные ацилглицерозы (кахексия, синдром гиперпродукции липотропина)
- Локальные кумулятивные ацилглицерозы (липоматозы, вакатное ожирение)
- Локальные резорбтивные ацилглицерозы (очаговые некрозы жировой ткани, регионарные липоатрофии).
II. Холестерозы (холестериновый диатез).
- Резорбтивный холестероз (болезнь Бе́ссена—Ко́рнцвейга)
- Кумулятивные холестерозы [атеросклероз, ксантоматоз, липоидная дуга роговицы].
Среди генерализованных кумулятивных ацилглицерозов основное значение имеет ожирение, но к данной группе мезенхимальных липодистрофий относятся также наследственные заболевания, сопровождающиеся развитием гиперацилглицеролемии. Классическим примером наследственной гиперацилглицеролемии является болезнь Бюргера—Грютца (гиперлипидемия Ia типа), при которой отсутствует фермент липопротеин-липаза, вследствие чего нейтральный жир хиломикронов не расщепляется и эти частицы длительно остаются в кровотоке (гиперхиломикронемия). Помимо хиломикронов пи наследственных гиперацилглицеролемиях в крови увеличивается содержание липопротеинов очень низкой плотности — частиц, также богатых нейтральными жирами.
Ожирение
Ожирение (общее ожирение, тучность, adipósitas) — патологический процесс, сопровождающийся существенным увеличением в организме массы белой жировой ткани. Общее количество ацилглицеролов при этом также значительно увеличивается. Минимальный показатель превышения «стандартной» массы тела при ожирении составляет 20% у мужчин и 25% у женщин (именно при таком избытке массы тела отмечается статистически значимое увеличение вторичной заболеваемости и смертности). Ожирение является одним из наиболее распространённых заболеваний в развитых странах мира. Процесс чаще развивается после 40 лет, преимущественно у женщин. Ожирение нередко сопровождается развитием паренхиматозной липодистрофии печени (жирового гепатоза).
При наличии у грудного ребёнка избытка массы тела за счёт избыточного развития жировой ткани термин «ожирение» не используется: по традиции такое состояние обозначают понятием паратрофи́я.
Классификация
Формы ожирения классифицируют следующим образом:
I. Этиологический принцип
- Первичное (алиментарно-конституциональное) ожирение
- Вторичное (симптоматическое) ожирение [наследственные и приобретённые варианты].
Формы вторичного приобретённого ожирения:
- Эндокринное ожирение
- Церебральное (гипоталамическое) ожирение
- Дуоденальное (дуоденопривное) ожирение [при удалении двенадцатиперстной кишки]
- Гипоксическое ожирение (первичный пиквикский синдром)
- Ятрогенное ожирение [например, длительное применение β-адреноблокаторов угнетает липолиз].
II. Выраженность процесса
- I степень (превышение массы тела над «стандартной» не более чем на 29%)
- II степень (30—49%)
- III степень (50—99%)
- IV степень (100% и более).
III. Преимущественная локализация отложений жира
- Симметричный (универсальный) тип
- Асимметричные типы (верхний, средний, нижний).
IV. Микроморфологические особенности белой жировой ткани
- Гипертрофическое ожирение
- Гиперпластическое ожирение
- Смешанный вариант ожирения.
V. Морфогенез ожирения
- Динамическая фаза (масса тела нарастает)
- Статическая фаза (масса тела относительно постоянна).
Симметричный и асимметричный типы ожирения
По внешним проявлениям выделяют симметричный и асимметричные типы ожирения. При симметричном типе белая жировая ткань относительно равномерно распределена по разным частям тела. При верхнем типе жировая клетчатка выражена в области лица, затылка, шеи, плечевого пояса и молочных желёз. При среднем типе отмечается развитие подкожной клетчатки передней брюшной стенки («жировой фартук»), при нижнем типе — в области ягодиц, бёдер и голеней. Верхний тип соответствует андроидному, или «яблочному» (характерному для мужчин), нижний — гиноидному, или «грушевидному» (типичному для женщин), ожирению. Это распределение вызвано различной чувствительностью липоцитов к половым гормонам: липоциты верхней половины тела более чувствительны к андрогенам, поэтому преобладание андрогенов над эстрогенами сопровождается развитием верхнего типа ожирения. Андроидный тип ожирения чаще сочетается с нарушенной толерантностью к глюкозе или с диабетом, гиперлипидемией, артериальной гипертензией, гиперандрогенией и гирсутизмом у женщин.
Гиперпластическое и гипертрофическое ожирение
По микроморфологическим особенностям жировой ткани различают гипертрофическое и гиперпластическое ожирение. Количество клеток белой жировой ткани достигает максимума в пубертатном периоде (у женщин их около 40 млрд., у мужчин вдвое меньше, что связывают с супрессивным действием тестостерона на развитие жировой ткани). Диагностика гипертрофического и гиперпластического ожирения проводится по материалу биопсии жировой клетчатки.
Гипертрофическое ожирение характерно для заболевания, появившегося в зрелом возрасте, оно развивается на фоне персистирующей гипергликемии и вторичной гиперинсулинемии, оно характеризуется увеличением объёма липоцитов за счёт накопления в их цитоплазме ацилглицеролов. Так, при увеличении диаметра клетки в 2 раза количество нейтрального жира в ней возрастает в 8 раз. Общее число жировых клеток при этом типе ожирения существенно не изменяется. Несмотря на уменьшение чувствительности гипертрофированных адипоцитов к инсулину, чувствительность этих клеток к липолитическим гормонам (половые и тиреоидные гормоны) увеличивается, поэтому возможно существенное снижение массы жировой ткани консервативными методами лечения. Коррекция массы тела при гипертрофическом ожирении необходима, т.к. оно сопровождается выраженными метаболическими нарушениями и приводит к тяжёлым изменениям прежде всего со стороны сердечно-сосудистой системы.
Напротив, гиперпластическое ожирение отличается увеличением числа липоцитов, содержание ацилглицеролов в клетке не меняется. Этот тип ожирения не связан с увеличением концентрации глюкозы и инсулина в крови. Метаболические расстройства при гиперпластическом ожирении не выражены, поэтому его течение, как правило, доброкачественное, однако снизить массу гиперплазированной жировой ткани консервативными методами затруднительно. В ряде случаев при ожирении сочетаются признаки гипертрофии и гиперплазии адипоцитов. Гиперпластическое или смешанное ожирение отмечается у лиц с избыточной массой тела с детства.
Этиология и патогенез ожирения
Любая форма ожирения, по существу, является алиментарной, т.е. связана с избыточным пищевым поведением (полифагией, или гиперфагией, крайняя степень выраженности которой называется булимией). Однако когда говорят об этиологии ожирения, имеют в виду причины, приводящие к чрезмерной пищевой активности. В этом отношении выделяют четыре основных этиологических фактора ожирения: (1) недостаточная мобилизация жиров из депо («синдром жадных липоцитов»), (2) торможение липолиза под действием гормонов (недостаточности липолитических и избытка антилиполитических), (3) поражение гипоталамического центра насыщения и (4) хроническая гипоксия. Как правило, перечисленные факторы действуют взаимосвязанно в разных комбинациях, но один из них обычно доминирует.
Недостаточная мобилизация жиров из депо обусловлена прежде всего гиперинсулинемией, возникающей при резистентности к инсулину периферических тканей. Обычно гиперинсулинемия при ожирении вторичная, особенно при первичном варианте ожирения. Она связана с рассредоточением цитолеммальных инсулиновых рецепторов по поверхности резко увеличенной жировой клетки и с влиянием контринсулярных факторов, продуцируемых, в частности, самими липоцитами (прежде всего ФНО-α). Гипертрофия жировых клеток, обусловливая снижение их чувствительности к инсулину, замыкает один из порочных кругов патогенеза ожирения. Инсулин является антилиполитическим гормоном, блокируя активность триглицерид-липазы адипоцитов. Развивающаяся при этом гипогликемия стимулирует пищевое поведение благодаря активации центра голода. Второй порочный круг формируют глюкокортикоиды, продукция которых при ожирении часто повышается.
Предрасполагающие к ожирению факторы. Способствуют развитию ожирения отягощённая наследственность, гиподинамия, обильные порции пищи, преобладание в пище жиров и дефицит белков (недостаток белка ослабляет специфическое динамическое действие пищи), а также никтофагия (употребление пищи преимущественно поздно вечером и ночью). Считается, что обильная белковая и углеводистая пища не относится к факторам, предрасполагающим к ожирению, хотя по отношению к углеводистой пище мнения различны.
Роль тиреоидных гормонов и бурого жира. Переедание далеко не всегда приводит к ожирению. Обычно при этом ускоряется метаболизм за счёт большего образования трийодтиронина, что не позволяет ацилглицеролам активно накапливаться в жировых депо. Кроме того, при избыточном питании отмечено разрастание бурой жировой ткани, клетки которой усиленно окисляют триглицериды, рассеивая избыток энергии с теплом.
Синдром Эрисихтона
В основе ожирения лежит увеличение концентрации в крови жиров (гиперлипидеми́я). Алиментарная гиперлипидемия рядом авторов удачно названа синдромом Эриси́хтона. История этого персонажа древнегреческих мифов, переосмысленная Овидием, любопытна и в медицинском отношении.
Эрисихтон (в буквальном переводе с древнегреческого «защитник земель») — сын фессалийского царя Триопа (поэтому иногда он носит имя Триопей), сам впоследствии ставший царём. Отличался крайним непочтением к богам («…презирая божественность Вышних, На алтарях никогда в их честь не курил фимиама»). Срубив дуб в священной роще, и тем самым погубив любимую нимфу-дриаду Деметры (Цереры), он навлёк на себя гнев богини. Дриада перед смертью прокляла Эрисихтона, а её сестры сообщили Деметре о святотатстве фессалийского царя. Разгневанная Деметра страшно наказала Эрисихтона, упросив богиню голода наделить его вечной неутолимой страстью к еде. Та исполнила волю Деметры, явившись к спящему Эрисихтону:
Сладостный сон между тем Эрисихтона нежил крылами
Мягкими: тянется он к соблазнительно снящимся яствам,
Тщетно работает ртом; изнуряет челюсть о челюсть,
Мнимую пищу глотать обольщённой старается глоткой.
Но не роскошную снедь, а лишь воздух пустой пожирает.
Проснувшись, Эрисихтон начинает с жадностью поглощать пищу, и уже не может остановиться («Алчет всё большего он, чем больше нутро наполняет. …в нём пища любая К новой лишь пище влечёт. Он есть, но утроба пустует.»). Вскоре царь растратил все свои богатства в тщетной попытке усмирить муки голода. У него осталась лишь дочь Мнестра, и её Эрисихтон продал, желая насытиться. Но божественная кара неотвратима — Эрисихтон погибает, пожирая собственное тело:
После того, как алчба состояние всё истощила,
Снова и снова еду доставляя лихому недугу,
Члены свои раздирать, зубами грызть Эрисихтон
Начал: тело питал, убавляяся телом,– несчастный!
(Публий Овидий Назон. Метаморфозы. Перевод С. Шервинского).
Первичное ожирение
Первичное ожирение — ожирение, развивающееся вследствие абсолютной или относительной недостаточности гормона жировой ткани лептина. Поскольку причина первичного ожирения установлена, его не следует называть идиопатическим, как это было принято ранее. Лептин секретируется «сытыми» липоцитами и посредством собственных рецепторов в цитолемме нейронов центра насыщения способствует выработке в нём глюкагоноподобного пептида I, который в свою очередь тормозит продукцию нейропептида Y нейронами центра голода. Нейропептид Y в настоящее время считается основным медиатором чувства голода, именно он стимулирует пищевое поведение.
Выделяют две формы первичного ожирения:
- Абсолютная недостаточность лептина (дефицит самого гормона) — 20% случаев первичного ожирения
- Относительная недостаточность лептина (снижение чувствительности к гормону лептиновых рецепторов центра насыщения) — 80% случаев ожирения. Относительная недостаточность лептина проявляется гиперлептинемией.
Несмотря на установленную причину первичного ожирения, на практике его обычно диагностируют путём исключения других (вторичных) форм болезни.
Эндокринное ожирение
Эндокринное ожирение — ожирение, развивающееся под влиянием гормонов. По современным представлениям, любая форма ожирения может быть отнесена к эндокринным расстройствам: при первичном ожирении прежде всего нарушается обмен лептина, при церебральном — нейропептида Y и глюкагоноподобного пептида I.
Однако по сложившейся традиции различают четыре основные формы вторичного эндокринного ожирения:
- Ожирение при гиперкортицизме (первичном и вторичном)
- Ожирение при гиперинсулинизме
- Ожирение при гипогонадизме
- Ожирение при гипотиреозе.
Глюкокортикоидные гормоны и инсулин относятся к антилиполитическим, половые и тиреоидные гормоны — к липолитическим, поэтому высокое содержание глюкокортикоидов (гиперкортицизм) и инсулина (гиперинсулинизм), а также снижение уровня половых (гипогонадизм) и тиреоидных (гипотиреоз) гормонов способствуют ожирению.
1. Ожирение при гипогонадизме. Гипогонадизм служит причиной ожирения при адипозогенитальной дистрофии. В основе адипозогенитальной дистрофии (синдрома Фрёлиха) лежит приобретённая недостаточность гонадотропинов гипофиза. Их дефицит приводит к недостаточности синтеза и секреции половых гормонов. Заболевание проявляется в препубертатном или пубертатном возрасте двумя основными синдромами, обусловленными гипогонадизмом: (1) гипогенитализмом (недоразвитием половых органов и вторичных половых признаков) и (2) ожирением. Характерно преимущественное отложение жира в области лица («кукольное лицо»), груди, живота и бёдер. Конечности полные. Нередко формируется ожирение гиноидного типа. Болеют в основном мальчики. Болезнь Лоуренса—Муна—Бидля — врождённый вариант адипозогенитальной дистрофии. Периферические формы гипогонадизма также сопровождаются развитием ожирения. При евнухоидизме жир преимущественно откладывается в области плечевого пояса, на груди, в области живота и бёдер. Сходные внешние проявления возникают при ожирении у мужчин в климактерическом периоде. Снижение функции яичников сопровождается накопление жира на плечах («климактерический горбик»), груди, животе, бёдрах.
2. Ожирение при гиперкортицизме. Гиперкортицизм лежит в основе болезни и синдрома Кушинга (Иценко—Кушинга). Жир при этом накапливается преимущественно в области лица («лунообразное лицо»), шеи, верхней половины туловища и живота. В области шейных позвонков формируется жировой «буйволиный горбик». Конечности худые. На коже живота, бёдер, плеч, молочных желёз появляются грубые багрово-красные полосы (стрии — разрывы подкожной жировой клетчатки), иногда с кровоизлияниями по периферии. Примерно у 10% больных ожирения не развивается, но типичное перераспределение жира происходит. Юношеский базофилизм (гипоталамический синдром пубертатного периода, пубертатно-ювенильный диспитуитаризм) также проявляется гиперкортицизмом (вторичный гиперкортицизм). Кроме того происходит усиление продукции соматотропного и гонадотропных гормонов. Гиперфункция аденогипофиза при этом обусловлена гиперпродукцией нейросекреторными клетками гипоталамуса соответствующих рилизинг-факторов. В ткани гипофиза увеличивается число базофильных клеток (отсюда термин базофилизм). Заболевание формируется в период полового созревания и проявляется ожирением, при котором жир в подкожной клетчатке распределяется равномерно, но иногда откладывается преимущественно в нижней части туловища, что у юношей обусловливает некоторую феминизацию внешности. В отличие от ожирения по типу Кушинга, конечности при ювенильном базофилизме остаются полными. Стрии тонкие, розовые, без кровоизлияний.
3. Ожирение при гиперинсулинизме. Гиперпродукция инсулина характерна для сахарного диабета II типа и особенно выражена при опухоли из инсулиноцитов (инсулиноме). При этом развивается ожирение с равномерным распределением жира или по андроидному типу.
4. Ожирение при гипотиреозе. Ожирение при гипотиреозе (микседеме) связано со снижением основного обмена. Избыточная масса тела обусловлена не только ожирением, но и «слизистым отёком» тканей, усиливающим впечатление тучности.
Кроме перечисленных форм, к эндокринному ожирению относят эпифизарное ожирение при синдроме Марбурга—Милку (гиперфункция эпифиза за счёт гиперплазии пинеоцитов или опухоли из них — пинеаломы).
Своеобразным вариантом эндокринного ожирения является дуоденальное (дуоденопривное) ожирение, развивающееся у больных, которым была удалена двенадцатиперстная кишка. Многие гормоны двенадцатиперстной кишки регулируют чувство голода, основным дуоденальным ингибитором голода служит холецистокинин, поэтому при удалении этого органа через некоторое время возникает полифагия и ожирение.
Церебральное ожирение
Церебральное (гипоталамическое) ожирение связано с поражением нейронов центра насыщения (центра сытости). Характер патологического процесса в головном мозге может быть различным — воспаление, опухоль, травма и т.д. При этом сохранный центр голода (центр аппетита) активируется, что определяет избыточное пищевое поведение. Центр насыщения расположен в вентромедиальных зонах гипоталамуса, центр голода — в латеральных. Двустороннее разрушение вентромедиальных зон приводит к выраженному ожирению экспериментальных животных в результате полифагии, повреждения латеральных зон могут вызвать отказ от еды (афагию) и смерть от истощения. И, напротив, хроническая стимуляция вживлёнными электродами вентромедиальных ядер гипоталамуса вызывает афагию, а латеральных — полифагию. Для церебрального ожирения характерно быстрое нарастание массы тела после травмы черепа, энцефалита или при опухоли мозга. Масса тела может увеличиваться даже при нормальном аппетите, хотя обычно отмечается полифагия. Ожирение, как правило, симметричное и достигает высоких степеней.
К церебральному относится также ожирение при болезни Барракера—Симонса (Barraquer—Simons), развивающейся у детей и молодых женщин после церебральной формы ревматизма (малой хореи) и при диффузных болезнях соединительной ткани. Ожирение по типу Барракера—Симонса характеризуется преимущественным отложением жира в области бёдер, ягодиц и передней брюшной стенки. На коже плечевого пояса появляются сине-багровые стрии. Интересно, что жир верхней половины тела атрофируется, начиная с головы («голова мертвеца»), поэтому данное заболевание можно отнести и к локальным резорбтивным ацилглицерозам.
Своеобразным патогенезом отличается церебральное ожирение при синдроме Стюарта—Морганьи—Мореля, при котором развивается гиперостоз костей черепа и облитерация их синусов. Вероятно, вследствие этого в промежуточном мозге больных нарушается кровообращение и происходит избирательное поражение центра насыщения. Болеют в основном девочки. Жир откладывается в области живота (андроидный тип ожирения).
Гипоксическое ожирение
Гипоксическое ожирение (первичный пиквикский синдром) обусловлено хронической гипоксией вследствие нейрогенной гиповентиляции лёгких. Липоциты относятся к производным малодифференцированных клеток фибробластического дифферона, которые способны активно пролиферировать при низком парциальном давлении кислорода в среде. Обычно длительная гипоксия провоцирует разрастание только фиброзной ткани, но в ряде случаев это происходит и с белой жировой тканью. Некоторые авторы полагают, что гипоксическое ожирение имеет церебральный характер.
Первичный пиквикский синдром включает (1) нарушение дыхания, прежде всего во сне, от чего сон становится беспокойным и сопровождается необычайно интенсивным храпом; (2) компенсаторную дневную сонливость (больные засыпают сразу же, оказавшись вне активной деятельности) и (3) ожирение. Синдром назван пиквикским по названию книги Чарлза Диккенса «Записки Пиквикского клуба», один из героев которой (Джо) страдал этим симптомокомплексом. Ожирение при первичном пиквикском синдроме обычно относится к среднему типу. У больных формируется правожелудочковая недостаточность (лёгочное сердце).
От первичного необходимо отличать вторичный пиквикский синдром, развивающийся на фоне уже имеющегося ожирения.
Наследственное ожирение
К наследственным формам ожирения относят любые его случаи, связанные с наследуемым дефектом генов. Часть заболеваний, проявляющихся наследственным ожирением, можно отнести к эндокринному или гипоталамическому типам ожирения. Например, при болезни Лоуренса—Муна—Бидля ожирение носит эндокринный характер. Болезнь Гирке (гликогеноз I типа) сопровождается постоянной гипогликемией из-за отсутствия нарушения гликогенолиза, вследствие чего больные часто и много едят, стараясь подавить приступы булимии. Жир при этом откладывается главным образом в подкожной клетчатке лица. Для болезни Гирке характерен также стеатоз печени. Патогенез ожирения при других наследственных болезнях не ясен (например, при синдромах Прадер—Вилли и Карпентера).
Осложнения
Основным осложнением ожирения является атеросклероз, связанный с гиперлипидемией. Значительно увеличивается риск развития артериальной гипертензии, сахарного диабета, холелитиаза, варикозного расширения вен нижних конечностей, подагры, острого аппендицита, цирроза печени (у мужчин) и ряда злокачественных опухолей. Избыточная масса тела однозначно укорачивает продолжительность жизни. Кроме того, развиваются липоматоз миокарда, периферическая гиперэстрогения, гиповентиляционный синдром (вторичный пиквикский синдром) и перегрузка костей и суставов (прежде всего позвоночника).
Висцеральный липоматоз. Ожирение характеризуется не только увеличением объёма жировой клетчатки, но и разрастанием жировой ткани в строме внутренних органов (висцеральный липоматоз). Наиболее опасен липоматоз миокарда: в участках выраженного липоматоза может произойти разрыв стенки сердца. Висцеральный липоматоз сочетается с андроидным типом ожирения. Липоциты внутренних органов более чувствительны к катехоламинам и выделяют больше ФНО-α (кахектина), обладающего контринсулярным потенциалом. За счёт этого висцеральные липоциты вносят наибольший вклад в развитие резистентности к инсулину при ожирении.
Периферическая гиперэстрогения. У больных ожирением нарушается периферический метаболизм половых гормонов. В жировой ткани происходит ускорение ароматизации андрогенов, в частности, тестостерона и андростендиона в эстрадиол и эстрон соответственно, что приводит к гиперэстрогении, способствующей возникновению метроррагии и доброкачественной дисплазии молочной железы, а у мужчин — гинекомастии, уменьшению роста волос на лице и теле. При этом и у мужчин и у женщин нарушается половая функция.
Регионарные кумулятивные ацилглицерозы
Регионарные кумулятивные ацилглицерозы включают (1) липоматозы и (2) вакатное ожирение.
Липоматозы
Липоматоз. Термин липоматоз применяется в двух значениях: (1) очаговое разрастание жировой ткани без чётких границ (например, липоматоз миокарда при ожирении) и (2) множественные липомы (опухоли белой жировой ткани). В некоторых случаях эти процессы сочетаются. Солитарные липомы и гиберномы (опухоли бурой жировой ткани) также можно отнести к регионарным кумулятивным ацилглицерозам. Липомы обычно представляют собой мягкие, инкапсулированные, не спаянные с окружающими тканями, жёлтые на разрезе и с поверхности узлы диаметром, как правило, от 1 до 10 см. Количество липом при липоматозе у одного пациента может быть различным, иногда превышает 100 узлов.
Распространённые по всему телу (включая и внутренние органы) липомы называют диффузным липоматозом. Различают детскую и взрослую формы диффузного липоматоза. Детская форма обычно проявляется в первые два года жизни.
Симметричные формы липоматоза особенно характерны для мужчин среднего возраста, длительно проживающих в странах средиземноморского региона. У многих из них в анамнезе имеются заболевания печени или хронический алкоголизм.
В практике патологоанатома наибольшее значение имеют болезни (1) Маделунга, (2) Деркума, (3) Рота—Пэйлэрда, (4) Лонуа—Бенсо, (5) Вернея—Потена, (6) Лери, (7) Мараньона, (8) Фере—Лангмида, (9) Пастера—Валлери—Радо—Бламутье, (10) тазовый и (11) стероидный липоматоз, а также (12) ВИЧ-липодистрофия, хотя в целом эти липоматозы редки.
1. Липоматоз Деркума (lipomatosis dolorosa) [Фрэнсис Де́ркум — американский невролог] характеризуется появлением липомоподобных болезненных узлов в подкожной клетчатке верхних конечностей, в коже молочных желёз, спины, передней брюшной стенки, ягодиц и бёдер. Болезненность определяется при пальпации, давлении и травме узла. Она связана с воспалительным процессом в нерве (невритом). Кожа над узлами цианотичная, нередки кровоизлияния. При микроморфологическом исследовании в ткани узлов обнаруживаются многочисленные кровеносные капилляры, в нервных стволиках отмечаются признаки продуктивного воспаления. В некоторых липомах возникает очаговый некроз жировой ткани и формируются типичные липогранулёмы с наличием гигантских многоядерных макрофагов. Заболевание типично для тучных женщин в климактерическом периоде. При липоматозе Деркума развиваются атрофические процессы в эндокринных железах, прежде всего щитовидной (гипотиреоз).
2. При синдроме Маделунга (множественном шейном симметричном липоматозе) [Отто фон Маделунг — немецкий хирург] в области шеи происходит разрастание жировой ткани в виде множественных болезненных узлов. Узлы сдавливают органы шеи, приводя к дисфагии и затруднению дыхания. Вероятно, липоматоз развивается на фоне гипофизарных нарушений.
3. При синдроме Рота—Пэйлэрда очаговые разрастания белой жировой ткани в поясничной области и на конечностях. Узлы располагаются симметрично.
4. Синдром Лонуа—Бенсо (симметричный липоматоз Лонуа—Бенсо) [Launois—Bensaude] характеризуется появлением симметричных липом в области шеи, живота и паховых складок. Особенно характерны узлы на передней и задней поверхностях шеи и в подчелюстной области. Иногда этот синдром в отечественной литературе называют «липоматозом Лонуа—Вансода» или «синдромом Лэньу—Бенсо».
5. Липоматоз Вернея—Потена отличается локализацией липом в надключичных областях.
6. Липоматоз Лери (диффузный симметричный липоматоз Лери) — множественные липомы, расположенные симметрично на верхних конечностях, в области ягодиц, реже на других участках кожного покрова.
7. Липоматоз Мараньона (гипертрофический липоматоз Мараньона) — симметричный распространённый липоматоз с поражением скелетных мышц (внутримышечные липомы). Особенностью внутримышечных липом является их нечёткие границы за счёт инвазивного роста. Такой узел спаян с окружающими тканями. Липомы при данном процессе придают больному «вид Геракла».
8. При липоматозе Фере—Лангмида жировые узлы располагаются на ладонях.
9. Липоматоз Пастера—Валлери—Радо—Бламутье характеризуется локализацией липом в подкожной клетчатке боковых поверхностей грудной клетки.
10. Тазовый липоматоз отличается расположением липом в параректальной и паравезикальной клетчатке. Наиболее часто процесс возникает у лиц негроидной расы различного возраста.
11. Стероидный липоматоз относится к ятрогенным формам липоматоза. Он возникает при длительном применении препаратов глюкокортикоидных гормонов. Липомы образуются прежде всего на лице, передней грудной стенке и над шейными позвонками («буйволиный горб»).
12. При ВИЧ-липодистрофии, помимо различных изменений жировой ткани у больных ВИЧ-инфекцией, может возникать и разрастание жировой ткани в области шеи, молочных желёз и во внутренних органах. Липоматоз при этом связывают с действием специфических антиретровирусных препаратов (ятрогенный липоматоз). На лице и конечностях жировая клетчатка, напротив, может исчезать. У больных выявляются гиперлипидемия и резистентность периферических тканей к инсулину.
Вакатное ожирение
Вака́тное ожирение (от лат. vacuus — пустой) — замещение белой жировой тканью атрофированной паренхимы органа. Наиболее часто вакатное ожирение наблюдается в скелетных мышцах. Орган при этом может увеличиваться (псевдогипертрофия).
Генерализованные резорбтивные ацилглицерозы
К генерализованным резорбтивным ацилглицерозам относятся (1) кахексия (истощение, маразм, общая патологическая атрофия) и (2) синдром гиперпродукции липотропина. Некоторые авторы кахексией называют крайнюю степень истощения (утрату более половины нормальной массы тела), однако в отечественной патологической анатомии эти понятия используются как синонимы.
Кахексия
Кахекси́я (от греч. kakos — плохой, hexis — состояние) развивается (1) при полном длительном голодании или мальабсорбции (алиментарная кахексия, экзогенная кахексия), (2) при нервной анорексии, (3) злокачественных опухолях («раковая кахексия»), (4) при атрофии и некрозе ткани гипоталамуса и гипофиза (гипоталамо-гипофизарная кахексия, или болезнь Симмондса—Шиена), (5) при хронических инфекциях (например, туберкулёзе или длительно протекающих гнойных процессах), (6) при ожоговой болезни («ожоговое истощение»), (7) при аутоиммунных заболеваниях, (8) при общей хронической застойной гиперемии, а также (9) при нарушении метаболизма ряда гормонов (тяжёлая хроническая надпочечниковая недостаточность, гипоинсулинизм, гиперглюкагонемия, гиперсоматостатинемия). «Раневое истощение» является вариантом кахексии при хронической гнойной инфекции («гнойно-резорбтивная лихорадка»). Ведущее значение в развитии эндогенных форм кахексии, кроме гипоталамо-гипофизарной, придаётся ФНО-α (кахектину), основным продуцентом которого являются макрофаги. Гипоталамо-гипофизарная кахексия обусловлена повреждением центра голода, вследствие чего перестаёт вырабатываться основной триггер пищевого поведения нейропептид Y. «Раковая кахексия» отличается более сложным патогенезом. Помимо характерного внешнего вида, для кахексии типичны вторичный липофусциноз (бурая атрофия миокарда и печени), а также концентрация в тканях липохромов (сохранившаяся жировая ткань приобретает вследствие этого буроватую окраску). Жировая клетчатка, особенно подкожная отсутствует. Кожа сухая и дряблая. Внутренние органы уменьшены. На определённой стадии истощения развиваются марантические отёки вследствие снижения онкотического давления в тканях.
Синдром гиперпродукции липотропина
Синдром гиперпродукции липотропина (гипермускулярная липодистрофия, липоатрофический диабет, синдром генерализованной липодистрофии) — заболевание, при котором существенно возрастает синтез и секреция аденогипофизом липотропных гормонов. Синдром встречается главным образом у женщин и проявляется в большинстве случаев в возрасте до 40 лет. Периферическая и висцеральная жировая ткань у больных практически отсутствует, сохраняется только жировая клетчатка на ладонях и подошвах, в ретроорбитальной, эпидуральной или параартикулярных областях, а при приобретённых формах — на лице. Тем не менее, у некоторых больных отмечается избыточное отложение подкожного жира в области лица и шеи. Для синдрома гиперпродукции липотропина характерны выраженная гиперлипидемия и гипергликемия на фоне значительной гиперинсулинемии, однако сахарный диабет при этом редко осложняется кетоацидозом. Гиперлипидемия приводит к развитию тяжёлого стеатоза печени, прогрессирующего в цирроз, а также атеросклероза. Различают наследственный (аутосомно-рецессивный тип наследования) и приобретённый варианты болезни. Ранее предполагалось, что при данном заболевании происходит увеличение продукции аденогипофизом соматотропного гормона, вследствие чего развивается гипертрофия скелетных мышц, внутренних органов и в ряде случаев возобновляется рост в зрелом возрасте, однако это мнение не подтвердилось. Мышечная гипертрофия и висцеромегалия обусловлены выраженной персистирующей эндогенной гиперинсулинемией. Естественная продолжительность жизни больных составляет 35—50 лет. Прогноз во многом зависит от времени манифестации заболевания: наиболее неблагоприятно протекает синдром, развившийся в периоде полового созревания. Основными причинами смерти являются цирроз печени и атеросклеротические поражения сосудов (инфаркт миокарда, ишемический инсульт).
Регионарные резорбтивные ацилглицерозы
Регионарные резорбтивные ацилглицерозы включают две группы патологических процессов: (1) очаговые некрозы жировой ткани (липонекрозы, стеатонекрозы) и (2) местную атрофию белой жировой ткани (регионарные липоатрофии).
Очаговые некрозы жировой ткани
Основными неинфекционными процессами, при которых происходит очаговый некроз жировой ткани, являются следующие формы регионарных ацилглицерозов:
- Болезнь Вебера—Крисчена (спонтанный панникулит)
- Травматическая липогранулёма
- Панкреогенный липонекроз
- Инсулиновый липонекроз.
Кроме того, некроз жировой ткани развивается при холодовом, пост-стероидном (при прекращении приёма препаратов глюкокортикоидных гормонов) и реактивном (вторичное изменение на фоне других заболеваний) панникулитах, а также у новорождённых (некроз подкожной клетчатки новорождённых) и детей раннего возраста (острый мультинодулярный диссеминированный нерецидивирующий липонекроз).
Болезнь Ве́бера—Кри́счена — аутоиммунное поражение жировой ткани. На месте некроза жировой ткани образуется узел, залегающий в гиподерме на разной глубине. Диаметр отдельных узлов от нескольких миллиметров до 10 см и более. Узлы могут располагаться на любом участке кожного покрова, они редко бывают единичными. Излюбленная локализация узлов — кожа конечностей. Иногда узлы сливаются в конгломераты. Крупные узлы содержат полости с желтоватой пенистой массой (жировым детритом) и клинически симулируют абсцесс или флегмону. Заболевание начинается с симптомов общей интоксикации, за исключением варианта Ротманна—Макаи, протекает с рецидивами и ремиссиями. Длительность спонтанного панникулита различна: от нескольких недель до нескольких десятилетий, чаще от недели до года.
Выделяют четыре формы болезни Вебера—Крисчена:
- Острую (редкое фатальное течение болезни с тяжёлым поражением печени)
- Подострую (форма средней тяжести, напоминающая сепсис, с умеренно выраженным поражением печени)
- Хроническую (протекает благоприятно с длительными ремиссиями без поражения печени)
- Форму Ро́тманна—Ма́каи (протекает благоприятно, общая реакция интоксикации отсутствует).
При микроморфологическом исследовании изменённой жировой ткани обнаруживается лимфогистиоцитарный инфильтрат, очаги некроза липоцитов с наличием пенистых макрофагов и образованием характерных жировых кист. В ходе резорбции детрита на его месте разрастается фиброзная ткань, придающая плотность очагам поражения.
Травматическая липогранулёма — очаговый некроз жировой клетчатки в месте её механического повреждения с последующим развитием продуктивного воспаления. Нередко этот процесс неправильно называют «травматической олеогранулёмой». Олеогранулёма (лат. oleum — масло) — очаг продуктивного воспаления в месте введения растительного жира (масляного раствора лекарственного препарата).
Панкреогенный липонекроз (панкреогенная липодистрофия) — некроз жировой ткани, развивающийся под действием ферментов поджелудочной железы (при остром панкреатите, распадающихся злокачественных опухолях этого органа).
Инсулиновый липонекроз (инсулиновая липодистрофия) возникает в местах инъекций инсулина. При этом в подкожной жировой клетчатке образуются плотные уплощённые узлы диаметром от 2 до 20 см с западением в центре и чёткими границами. Кожа над узлом не смещается, т.к. спаяна с ним. Узлы могут появляться в удалённых от мест инъекций участках.
Регионарные липоатрофии
К регионарным липоатрофиям можно отнести упоминавшуюся в разделе «Церебральное ожирение» болезнь Барракера—Симонса (липоатрофия верхней части тела и ожирение нижней). Известны и другие местные липоатрофии: аутосомно-доминантная липоатрофия конечностей, циркулярная и полуциркулярная липоатрофия конечностей, опоясывающая абдоминальная липоатрофия у грудных детей и др.
Холестерозы
Нарушение обмена холестерола и холестеридов проявляется прежде всего накоплением их в тканях (кумулятивные холестерозы), особенно в стенке артерий и в коже, но известно наследственное заболевание с недостаточностью холестерола (резорбтивный холестероз). Холестерол относится к необходимым веществам: из него синтезируются стероидные гормоны, витамин D, желчные кислоты, он входит в состав биомембран всех клеток, поэтому дефицит холестерола приводит к не менее разрушительным последствиям для организма, чем его избыток.
Резорбтивный холестероз
Недостаточность холестерола отмечается у детей с наследственной болезнью Бе́ссена—Ко́рнцвейга, при которой формируется дефект ферментов синтеза холестерола различными клетками организма. Наиболее тяжёлые изменения возникают в головном мозге: его недоразвитие (гипоплазию) связывают именно с дефицитом холестерола и холестеридов. Недоразвитие мозговой ткани сопровождается олигофренией.
Кумулятивные холестерозы
Различают наследственные и приобретённые формы кумулятивных холестерозов. Накопление холестерола и холестеридов в тканях обусловлено прежде всего их поступлением с пищей (экзогенный холестерол), хотя определённую роль играет высокая синтетическая активность клеток (эндогенный холестерол). Холестерол и холестериды откладывают чаще всего в стенках артерий (атеросклероз), дерме (ксантоматоз) и в наружной оболочке глаза (липоидная дуга роговицы). Спектр изменённых тканей при наследственных кумулятивных холестерозах существенно шире: например, при болезни острова Танжер характерно накопление холестерола в нёбных миндалинах («болезнь оранжевых миндалин») и в других лимфоидных органах.
Основной причиной накопления холестерола в тканях является недостаточность механизма его обратного транспорта. Ключевым фактором системы обратного транспорта холестерола (с периферии в печень, откуда его излишки удаляются из организма с желчью) являются липопротеины высокой плотности, точнее входящий в их состав белок апопротеин А. Апопротеин А синтезируется гепатоцитами и покровными энтероцитами, он обладает уникальной способностью поглощать холестерол и его эфиры («магнит для холестерина»). Частицы липопротеинов высокой плотности собирают холестерол не только в интерстициальном секторе, но и внутри клеток. У человека (а также высших обезьян и свиней) существует видовая (характерная для всех представителей вида) недостаточность апопротеина А и, соответственно, липопротеинов высокой плотности. Животные с высоким содержанием этих липопротеинов не страдают холестериновым диатезом, даже при постоянном употреблении богатой холестеролом пищи. Некоторые люди также отличаются довольно высокой концентрацией апопротеина А («синдром долголетия»).
Мезенхимальные углеводные дистрофии
Мезенхимальные углеводные дистрофии включают (1) нарушения обмена гликозаминогликанов, к которым относятся наследственные заболевания мукополисахаридозы, и (2) нарушения обмена гликопротеинов (ослизнение ткани).
Мукополисахаридозы
Мукополисахаридо́зы — тезаурисмозы, сопровождающиеся накоплением в тканях гликозаминогликанов. Термин «мукополисахаридозы» происходит от устаревшего обозначения протеогликанов (нейтральных мукополисахаридов) и входящих в их состав гликозаминогликанов (кислых мукополисахаридов). Для мукополисахаридозов характерен синдром гаргоили́зма (от фр. gargouille — гротескная фигурка, напоминающая человека, которыми украшали крыши, карнизы и другие детали старых домов): массивный череп, короткая шея, западающее широкое переносье, гипертелоризм, толстые губы, высокое нёбо, сросшиеся брови, большой язык.
Наиболее изучена патологическая анатомия первых шести типов мукополисахаридозов: болезни Пфаундлера—Гурлер (I тип), Гюнтера (II тип), Санфилиппо (III тип), Брейлсфорда—Мо́ркио (IV тип), Шейе (V тип) и Марото́—Лями́ (VI тип).
1. Чаще встречается болезнь Пфаундлера—Гурлер [Райнхард фон Пфаундлер и Гертруда Гурлер — немецкие врачи], в основе которой лежит дефицит α-L-идуронидазы. В тканях происходит накопление дерматан- и гепарансульфатов, значительно повышено их содержание в моче. Симптомы появляются к концу первого года жизни. Заболевание быстро прогрессирует, синдром гаргоилизма сочетается с глубокими нарушениями психики, ранним помутнением роговицы, глухотой, гепато- и спленомегалией. Смерть наступает обычно в возрасте 10—12 лет при поражении головного мозга (чаще вследствие гидроцефалии), сердца или от интеркуррентных заболеваний.
При макроморфологическом исследовании обнаруживаются уменьшенные и уплощенные позвонки с шиповидными выростами, межпозвонковые диски грибовидной формы, т.к. они больше поперечника тела позвонка. Трубчатые кости искривлены, эпифизарные линии изогнуты, зазубрены. При микроморфологическом исследовании выявляются нарушения периостального и энхондрального окостенения из-за отложения гликозаминогликанов в хрящевой ткани и в периосте. Хондроциты вздуты, овальной формы, хрящевых колонок не образуют, поэтому рост костей в длину задерживается. Основное вещество и коллагеновые волокна сухожилий, фасций, стенок сосудов, клапанов сердца содержит большое количество гликозаминогликанов, хорошо выявляемых метахроматической окраской. Миокард гипертрофирован, клапанный и париетальный эндокард, сухожильные хорды и стенки венечных артерий утолщены. В митральном и аортальном клапанах отмечаются васкуляризация и кальциноз, отложение гликозаминогликанов в межуточном веществе и наличие клеток накопления Гурлер. Гликозаминогликаны обнаруживаются в гепатоцитах, макрофагах селезёнки и лимфоузлов, клетках аденогипофиза, в роговице, гонадах, мягкой мозговой оболочке, периферических нервах, нейронах головного мозга и вегетативных ганглиев. При электронной микроскопии в клетках выявляются включения, соответствующие повреждённым лизосомам, имеющие вид светлых прозрачных вакуолей, «зебровидных» или аморфных телец.
2. При болезни Гюнтера в тканях накапливаются дерматансульфат и гепарансульфат вследствие недостаточности фермента идуронат-сульфатазы. Тип наследования — рецессивный, сцепленный с X-хромосомой. Течение заболевания длительное по сравнению с мукополисахаридозом I типа. Проявления выражены умеренно, помутнение роговицы в большинстве случаев отсутствует, но характерна прогрессирующая глухота.
3. Болезнь Санфилиппо [Сильвестр Дж. Санфилиппо — американский педиатр]. Различают четыре клинико-лабораторных варианта болезни (вариант A обусловлен дефектом гепарансульфат-сульфатазы, B — α-N-ацетилглюкозаминидазы, C — α-глюкозаминид-N-ацетилтрансферазы, D — ацетилглюкозамин-6-сульфат-сульфатазы). Все варианты болезни фенотипически идентичны. В организме увеличено содержание в основном гепарансульфата, которые накапливаются в нейронах головного мозга, стенках артерий, в печени и селезёнке.
4. Болезнь Брейлсфорда—Мо́ркио [Джеймс Фредерик Брейлсфорд — английский врач, Луис Мóркио (1867—1935) — уругвайский педиатр] проявляется накоплением кератан- и дерматансульфатов. За счёт распада метаэпифизарного хряща больные низкого или карликового роста. Интеллект обычно нормальный.
5. Болезнь Шейе [Гарольд Г. Шейе — американский офтальмолог] характеризуется накоплением в основном дерматансульфата. Заболевание манифестирует лишь в зрелом возрасте. Характерен низкий или карликовый рост. Интеллект нормальный.
6. При болезни Марото́—Лями́ [Пьер Марото́ — французский педиатр и генетик, Мори́с Лями́ (1895—1975) — французский генетик] происходит накопление дерматансульфата. Типичен карликовый рост больных.
К мукополисахаридозам близки муколипидозы и наследственные гликопротеинозы.
Ослизнение ткани
Ослизне́ние тканей — замещение волокнистой и других соединительных тканей слизеподобной массой, образующейся при распаде протеогликанов и высвобождении из них гликозаминогликанов. Ослизнение тканей встречается при (1) кахексии и (2) выраженном гипотиреозе (микседеме). Фиброзная, жировая и хрящевая ткани становятся полупрозрачными, набухшими. При микроморфологическом исследовании их клетки приобретают звёздчатую или причудливо отростчатую форму, коллагеновые волокна (в отличие от мукоидного отёка) также замещаются слизеподобной массой. Процесс может быть обратимым, его прогрессирование приводит к некрозу ткани с образованием кист, заполненных мукоидом.
См. также
Литература
- Давыдовский И. В. Общая патологическая анатомия. 2-е изд.— М., 1969.
- Дифференциальная диагностика кожных болезней: Руководство для врачей / Под ред. Б. А. Беренбейна и А. А. Студницына.— М., 1989.
- Зайчик А. Ш., Чурилов Л. П. Основы патохимии. 2-е изд.— СПб, 2001.
- Калитеевский П. Ф. Макроскопическая дифференциальная диагностика патологических процессов.— М., 1987.
- Микроскопическая техника: Руководство для врачей и лаборантов / Под ред. Д. С. Саркисова и Ю. Л. Перова.— М., 1996.
- Общая патология человека: Руководство для врачей / Под ред. А. И. Струкова, В. В. Серова, Д. С. Саркисова: В 2 т.— Т. 1.— М., 1990.
- Патологическая анатомия болезней плода и ребёнка / Под ред. Т. Е. Ивановской, Б. С. Гусман: В 2 т.— М., 1981.
- Саркисов Д. С. Очерки истории общей патологии.— М., 1988 (1-е изд.), 1993 (2-е изд.).
- Саркисов Д. С., Пальцев М. А., Хитров Н. К. Общая патология человека.— М., 1997.
- Струков А. И., Серов В. В. Патологическая анатомия.— М., 1995.
- Теппермен Дж., Теппермен Х. Физиология обмена веществ и эндокринной системы.— М., 1989. [Цитата: «… жировая ткань — это своеобразный энергетический банк; вклады в него поступают во время еды, а выдачи … могут производиться в любое последующее время».]
- Fletcher C. D. M., Unni K. K., Mertens F. (Eds.): World Health Organization Classification of Tumours. Pathology and Genetics of Tumours of Soft Tissue and Bone.— Lyon: IARC Press, 2002.— С. 23—24. [липоматоз]
%PDF-1.6 % 1 0 obj > endobj 5 0 obj /ModDate (D:20160622160551+03’00’) /Subject >> endobj 2 0 obj > stream application/pdf
Лечение рака эндометрия у больных с ожирением роботом да Винчи
Эпидемия ожирения с каждым годом все больше набирает обороты. Так, по данным ВОЗ за 2015 год более 1,9 миллиарда (39%) людей в возрасте старше 18 лет имели избыточный вес, при этом 600 миллионов, то есть 13% из них страдали от ожирения. За последние 30 лет распространенность ожирения среди взрослого населения планеты увеличилась на 27,5%. Россия занимает 4-ое место по распространенности ожирения как среди женщин (28,5%), так и среди мужчин (15,3%), уступая только США, Китаю и Индии.
Классификация ожирения
Для определения степени избыточности веса определяют индекс массы тела (ИМТ), который рассчитывается по формуле: вес в килограммах / рост в метрах в квадрате.
Классификация ожирения в зависимости от Индекса массы тела
| Типы массы тела | ИМТ (кг/м2) | Риск сопутствующих заболеваний |
| Дефицит массы тела | <18,5 | Низкий (повышен риск других заболеваний) |
| Нормальная масса тела | 18,5-24,9 | Обычный |
| Избыточная масса тела | 25,0-29,9 | Повышенный |
| Ожирение I степени | <30,0-34,9 | Высокий |
| Ожирение II степени | <35,0-39,9 | Очень высокий |
| Ожирение III степени | > 40 | Чрезвычайно высокий |
Как ожирение влияет на репродуктивное здоровье
Доказано, что ожирение значимо увеличивает риск развития заболеваний репродуктивной системы: нарушений менструального цикла, аменореи1, синдрома поликистозных яичников2, дисфункциональных маточных кровотечений, миомы матки, гиперпластических процессов эндометрия3, бесплодия, рака эндометрия и яичников8,9.
Ожирение приводит к нарушению гормонального фона:
- повышает синтез эстрогенов4,
- нарушению транспорта стероидов5,
- изменению активности ферментативных систем стероидного метаболизма,
- повышению синтеза гонадотропинов6,
- нарушению чувствительности тканей к инсулину7.
Длительное воздействие эстрогенов вызывает патологические изменения в эндометрии. По данным многочисленных исследований рак эндометрия у пациенток с ожирением встречается до 9 раз чаще по сравнению с женщинами с нормальной массой тела.
Как проявляется рак эндометрия
У женщин репродуктивного возраста рак эндометрия чаще всего проявляться обильными длительными менструациями и нарушением менструального цикла. Однако в большинстве случаев заболевание возникает у пациенток в постменопаузе, основным симптомом его являются кровянистые выделения из влагалища.
Диагностика заболевания
Для определения рака эндометрия выполняется ультразвуковое исследование органов малого таза, однако более объективный результат у пациентов с ожирением дает МРТ (точность определения глубины инвазии с помощью МРТ составляет 71—97%)10,11. Подтвердить злокачественность новообразования помогает гистологическое исследование тканей эндометрия, полученной при биопсии или раздельном диагностическом выскабливании полости матки и цервикального канала.
Лечение рака эндометрия
Основным методом лечения рака эндометрия является удаление тела и шейки матки, яичников и маточных труб с обеих сторон, такая операция носит название «экстирпация матки с придатками». В ряде случаев требуется лимфаденэктомия — удаление лимфатических узлов тазовой и поясничной области12,13.
Операция может осуществляться открытым, влагалищным, лапароскопическим и высокотехнологичным способом с использованием хирургического робота da Vinci.
Преимущества операции роботом da Vinci
Пациенты с ожирением относятся к группе повышенного анестезиологического и хирургического риска, поэтому они нуждаются в опытных специалистах и наличии высокотехнологичного оборудования — хирургического робота da Vinci. Преимущества такого подхода очевидны:
- эффективная эргономика, трехмерное изображение, простота манипулирования инструментами для хирурга потенциально значительно повышает благоприятный исход операции;
- визуализация в глубоких тазовых областях у пациентов с ожирением в разы выше при использовании робота Да Винчи, чем при открытой или эндоскопической операции;
- применение хирургического робота повышает безопасность выполнения операций высокой сложности у больных с выраженным ожирением;
- травматизация окружающих тканей при использовании робота минимальна, это позволяет значительно уменьшить кровопотери в ходе вмешательства, а также интенсивность боли и осложнений в постоперационный период14-18;
- высокотехнологичная, малоинвазивная операция обеспечивает высокий результат лечения и ускоренный период реабилитации14-18.
Как работает робот da Vinci?
Хирургический робот da Vinci работает под полным управлением опытного хирурга через разрезы в брюшной стенке не более 1 см.
Благодаря специальной видеокамере-эндоскопу специалист получает очень подробное и точное изображение органов, что позволяет ему полностью контролировать операционное поле и управлять работой инструментов внутри брюшной полости с помощью специальных джойстиков. Размер оперирующих инструментов всего 8 мм в диаметре, при этом они имеют семь степеней свободы движения, что обеспечивает им большую артикуляцию, чем у человеческого запястья.
На нашем сайте вы можете найти более подробную информацию о технологии da Vinci, а также о платных и бесплатных способах получения высокотехнологичной помощи.
Вопросы, которые пациентке стоит задать своему лечащему врачу
19- Какие методики лечения подходят в моем случае?
- Какие последствия меня ждут, если я откажусь от хирургического вмешательства?
- В чем разница между открытой, лапароскопической и робот-ассистированной опреацией?
- Расскажите о ваших навыках, опыте работы с da Vinci и о результатах лечения ваших пациентов с использованием технологии da Vinci.
- Как я буду себя чувствовать после операции?
Как и в отношении любой операции, никто не может гарантировать наличие перечисленных преимуществ в каждом случае, поскольку каждый случай уникален. Пациенты должны обсуждать соответствующие хирургические риски со своими врачами.
Подробнее о хирургических рисках
Подробное исследование о лечении пациенток с ожирением робот-ассистированными методом вы можете прочесть в научной статье «Роль робот-ассистированной хирургии в лечении заболеваний внутренних половых органов у пациенток с морбидным ожирением».
В чем разница между гипертрофическим ожирением и гиперклеточным ожирением?
Круглый стол по решениям для борьбы с ожирением, Совет по пищевым продуктам и питанию, Отдел здравоохранения и медицины, Национальные академии наук, инженерии и медицины. Ожирение в раннем детстве: состояние науки и реализация многообещающих решений: итоги семинара. 2016 г. 23 мая. Доступно по адресу http://www.ncbi.nlm.nih.gov/books/NBK368372/.
Шиллер Дж.С., Кларк Т.К., Норрис Т.Ранняя публикация избранных оценок на основе данных национального опроса по вопросам здоровья, проведенного в январе – сентябре 2017 года. Программа раннего выпуска Национального опроса о состоянии здоровья . Март 2018 г. Доступно по адресу https://www.cdc.gov/nchs/data/nhis/earlyrelease/EarlyRelease201803.pdf.
Фреллик М. Уровень ожирения среди взрослого населения США резко вырос за последние 20 лет. Medscape Medical News . 2018 15 марта [Полный текст].
Wijga AH, Scholtens S, Bemelmans WJ, de Jongste JC, Kerkhof M, Schipper M, et al.Сопутствующие заболевания ожирения у школьников: перекрестное исследование в когорте рожденных PIAMA. BMC Public Health . 2010, 9 апреля, 10: 184. [Медлайн]. [Полный текст].
Li C, Ford ES, Zhao G, Croft JB, Balluz LS, Mokdad AH. Распространенность клинически диагностированного апноэ во сне в зависимости от статуса ожирения у мужчин и женщин: Национальное обследование здоровья и питания, 2005–2006 гг. Предыдущая Med . 2010 июль 51 (1): 18-23. [Медлайн].
Катял Н, Боллу ПК.Вентиляция, синдром ожирения-гиповентиляции. 2018, январь [Medline]. [Полный текст].
Jiao L, Berrington de Gonzalez A, Hartge P, Pfeiffer RM, Park Y, Freedman DM, et al. Индекс массы тела, модификаторы эффекта и риск рака поджелудочной железы: объединенное исследование семи проспективных когорт. Контроль причин рака . 2010 21 августа (8): 1305-14. [Медлайн]. [Полный текст].
Kyrgiou M, Kalliala I, Markozannes G, et al. Ожирение и рак на основных анатомических участках: общий обзор литературы. BMJ . 2017 28 февраля. 356: j477. [Медлайн]. [Полный текст].
Малкахи Н. «Веские доказательства»: ожирение связано с 11 видами рака. Medscape Medical News . 2017 28 февраля. [Полный текст].
Ореопулос А., Падвал Р., Макалистер Ф.А., Эзековиц Дж., Шарма А.М., Калантар-Заде К. и др. Связь между ожирением и качеством жизни, связанным со здоровьем, у пациентов с ишемической болезнью сердца. Инт Дж. Обес (Лондон) . 2010 сентябрь 34 (9): 1434-41.[Медлайн].
Galtier-Dereure F, Boegner C, Bringer J. Ожирение и беременность: осложнения и стоимость. Ам Дж. Клин Нутр . 2000 Май. 71 (5 доп.): 1242S-8S. [Медлайн].
Wadden TA, Webb VL, Moran CH, Bailer BA. Модификация образа жизни при ожирении: новые разработки в диете, физической активности и поведенческой терапии. Тираж . 2012 6 марта. 125 (9): 1157-70. [Медлайн]. [Полный текст].
Коули Дж., Мейерхофер К.Затраты на лечение ожирения: метод инструментальных переменных. J Health Econ . 2012 31 января (1): 219-30. [Медлайн].
Финкельштейн EA, DiBonaventura Md, Burgess SM, Hale BC. Издержки ожирения на рабочем месте. Дж. Оккуп Энвирон Мед . 2010 Октябрь 52 (10): 971-6. [Медлайн].
Рынки товаров и услуг для снижения веса. BCC Research. Доступно на http://www.bccresearch.com/report/weight-loss-markets-products-services-fod027c.html. Доступ: 23 апреля 2012 г.
МакКолл Б. Хроническое заболевание, связанное с ожирением: новое название ожирения ?. Medscape Medical News . 2016 29 декабря. [Полный текст].
Галлахер Д., Хеймсфилд С.Б., Хео М., Джебб С.А., Мургатройд П.Р., Сакамото Ю. Диапазоны здорового процентного содержания жира в организме: подход к разработке рекомендаций, основанных на индексе массы тела. Ам Дж. Клин Нутр . 2000 Сентябрь 72 (3): 694-701. [Медлайн].
Палата LC.Сегментный анализ биоэлектрического импеданса: обновленная информация. Curr Opin Clin Nutr Metab Care . 2012 Сентябрь 15 (5): 424-9. [Медлайн].
Shiwaku K, Anuurad E, Enkhmaa B, Kitajima K, Yamane Y. Соответствующий ИМТ для азиатского населения. Ланцет . 2004, 27 марта. 363 (9414): 1077. [Медлайн].
Гранди С. М., Брюер Х. Б. мл., Климан Дж. И., Смит С. К. мл., Ленфант С. Определение метаболического синдрома: Отчет Национального института сердца, легких и крови / Конференция Американской кардиологической ассоциации по научным вопросам, связанным с определением. Тираж . 2004, 27 января. 109 (3): 433-8. [Медлайн]. [Полный текст].
Tan CE, Ma S, Wai D, Chew SK, Tai ES. Можем ли мы применить определение метаболического синдрома Национальной образовательной программы по холестерину для азиатов? Уход за диабетом . 2004 Май. 27 (5): 1182-6. [Медлайн]. [Полный текст].
Тирош А., Шай И., Афек А., Дубнов-Раз Г., Аялон Н., Гордон Б. и др. Траектория ИМТ подростков и риск диабета по сравнению с коронарной болезнью. N Engl J Med . 2011, 7 апреля. 364 (14): 1315-25. [Медлайн].
Montonen J, Boeing H, Schleicher E, Fritsche A, Pischon T. Связь изменений индекса массы тела в раннем взрослом возрасте и в более позднем взрослом возрасте с циркулирующими концентрациями биомаркеров ожирения у мужчин и женщин среднего возраста. Диабетология . 2011 Июль 54 (7): 1676-83. [Медлайн].
Sugerman HJ, Kellum JM, Engle KM, Wolfe L, Starkey JV, Birkenhauer R, et al.Шунтирование желудка для лечения тяжелого ожирения. Ам Дж. Клин Нутр . 1992, 55 февраля (2 доп.): 560S-566S. [Медлайн].
Sugerman HJ. Последствия повышения внутрибрюшного давления при тяжелом ожирении. Surg Clin North Am . 2001 Oct. 81 (5): 1063-75, vi. [Медлайн].
Losina E, Walensky RP, Reichmann WM, Holt HL, Gerlovin H, Solomon DH, et al. Влияние ожирения и остеоартроза коленного сустава на заболеваемость и смертность пожилых американцев. Энн Интерн Мед. . 2011 15 февраля. 154 (4): 217-26. [Медлайн].
Адельман Р.Д., Рестайно И.Г., Алон США, Блоуи Д.Л. Протеинурия и очаговый сегментарный гломерулосклероз у подростков с тяжелым ожирением. J Педиатр . 2001 Апрель 138 (4): 481-5. [Медлайн].
Kasiske BL, Napier J. Гломерулярный склероз у пациентов с массивным ожирением. Ам Дж. Нефрол . 1985. 5 (1): 45-50. [Медлайн].
Дженнетт Дж. К., Чарльз Л., Грабб В.Гломеруломегалия и очаговый сегментарный гломерулосклероз, ассоциированные с ожирением и синдромом апноэ во сне. Am J Kidney Dis . 1987 декабрь 10 (6): 470-2. [Медлайн].
Hairston KG, Bryer-Ash M, Norris JM, Haffner S, Bowden DW, Wagenknecht LE. Продолжительность сна и пятилетнее накопление абдоминального жира в меньшинстве: семейное исследование IRAS. Сон . 2010 марта 33 (3): 289-95. [Медлайн]. [Полный текст].
Spiegel K, Tasali E, Penev P, Van Cauter E.Краткое сообщение: сокращение сна у здоровых молодых мужчин связано со снижением уровня лептина, повышенным уровнем грелина, а также повышенным голодом и аппетитом. Энн Интерн Мед. . 2004, 7 декабря. 141 (11): 846-50. [Медлайн].
Центры по контролю и профилактике заболеваний. Коронавирусная болезнь 2019 (COVID-19): люди любого возраста с сопутствующими заболеваниями. CDC. Доступно по адресу https://www.cdc.gov/coronavirus/2019-ncov/need-extra-precautions/people-with-medical-conditions.html. Обновлено 25 июня 2020 г .; Дата обращения: 27 июня 2020 г.
Всемирная федерация ожирения. COVID-19 и ожирение: Атлас 2021 г. Доступно на https://www.worldobesityday.org/assets/downloads/COVID-19-and-Obesity-The-2021-Atlas.pdf. Март 2021 г .; Дата обращения: 8 марта 2021 г.
Mascarenhas L, Rahim Z. Показатели смертности от Covid-19 в 10 раз выше в странах, где большинство взрослых имеют избыточный вес, говорится в отчете. CNN Health. Доступно по адресу https://www.cnn.com/2021/03/04/health/obesity-covid-death-rate-intl/index.html. 5 марта 2021 г .; Дата обращения: 8 марта 2021 г.
Гарг С., Ким Л., Уитакер М. и др. Частота госпитализаций и характеристики пациентов, госпитализированных с лабораторно подтвержденным коронавирусным заболеванием, 2019 г. — COVID-NET, 14 штатов, 1–30 марта 2020 г. MMWR . 2020 8 апреля [Полный текст].
Зажигалка J, Филлипс М., Хохман С. и др. Ожирение у пациентов моложе 60 лет является фактором риска госпитализации Covid-19. Clin Infect Dis .2020 апр. 9 [Medline]. [Полный текст].
Дэйвпорт Л., Наингголан Л. Связь ожирения с тяжелой формой COVID-19, особенно в возрасте до 60 лет. Medscape Medical News . 2020 16 апреля [Полный текст].
Касс Д.А., Дуггал П., Чинголани О. Ожирение может перенести тяжелую болезнь COVID-19 в более молодой возраст. Ланцет . 2020 4 мая. [Medline]. [Полный текст].
Саламон М. Ожирение может перенести тяжелую форму COVID-19 на более молодые возрастные группы. Medscape Medical News . 2020 11 мая [Полный текст].
Cai Q, Chen F, Wang T, et al. Ожирение и тяжесть COVID-19 в специализированной больнице в Шэньчжэне, Китай. Уход за диабетом . 2020 14 мая. [Medline]. [Полный текст].
Гао Ф, Чжэн К.И., Ван XB и др. Ожирение является фактором риска более серьезного заражения COVID-19. Уход за диабетом . 2020 14 мая. [Medline]. [Полный текст].
Такер МЭ.История диабета и COVID-19 вызывает беспокойство. Medscape Medical News . 2020 26 мая. [Полный текст].
Компанияц Л, Гудман А.Б., Брук Б. и др. Индекс массы тела и риск госпитализации, связанной с COVID-19, поступления в отделение интенсивной терапии, инвазивной механической вентиляции и смерти — США, март – декабрь 2020 г. MMWR Morb Mortal Wkly Rep . 2021 8 марта [Полный текст].
Макнамара Д. Данные CDC усиливают связь между ожирением и тяжелым COVID. Medscape Medical News . 2021 8 марта [Полный текст].
Гао М., Пьернас С., Эстбери Н.М. и др. Связь между индексом массы тела и серьезностью COVID-19 у 6,9 миллионов человек в Англии: проспективное когортное исследование на уровне сообществ. Ланцет, диабет, эндокринол . 2021 28 апреля. [Medline]. [Полный текст].
Мелвилл NA. Тяжесть COVID-19 начинается с нормального диапазона ИМТ, особенно у молодых. Medscape Medical News .2021 5 мая. [Полный текст].
Szatmary P, Arora A, Raraty MGT, Dunne DFJ, Baron RD, Halloran CM. Возникающий фенотип панкреатита, ассоциированного с SARS-CoV2. Гастроэнтерология . 1 июня 2020 г. [Medline]. [Полный текст].
Pass W. Панкреатит, связанный с COVID, может непропорционально сильно поражать молодых мужчин с избыточным весом. Доктор медицины семейной медицины . 2020 24 июня [Полный текст].
McCall B. Ожирение более опасно для мужчин с COVID-19, чем для женщин. Medscape Medical News . 2021 6 мая [Полный текст].
Guerson-Gil A, Palaiodimos L, Assa A, et al. Влияние тяжелого ожирения на исходы госпитализированных пациентов с COVID-19 с учетом пола: большое ретроспективное исследование, проведенное в Бронксе, штат Нью-Йорк. Eur J Clin Microbiol Infect Dis . 2021 6 мая. [Medline]. [Полный текст].
Буско М. Ожирение — наибольший риск развития пневмонии COVID-19, пожилой, мужской пол. Medscape Medical News .2020 5 ноября. [Полный текст].
Аминиан А., Бена Дж., Панталоне К.М., Бургера Б. Ассоциация ожирения с пост-острыми последствиями COVID-19 (PASC). Метаб. Диабета, ожирения . 2021 г., 1 июня. [Medline].
Мелвилл NA. Ожирение увеличивает риск длительного COVID, результаты исследования. Новости здоровья WebMD . 2021 8 июня [Полный текст].
Yao R, Ananth CV, Park BY, Pereira L, Plante LA, для Консорциума перинатальных исследований.Ожирение и риск мертворождения: популяционное когортное исследование [аннотация]. Представлено на: Ежегодном собрании SMFM 2014 г .; 3-8 февраля 2014 г .; Новый Орлеан, Луизиана. Am J Obstet Gynecol . 2014. 210: [Полный текст].
Hackethal V. Риск мертворождения у женщин с ожирением может быть на 25% выше. Medscape Medical News . 27 марта 2014 г. [Полный текст].
Cameron NA, Petito LC, McCabe M, et al. Количественная оценка бремени ожирения по признаку пола, расы / этнической принадлежности на заболеваемость сахарным диабетом в США, 2001–2016 гг .: MESA и NHANES. J Am Heart Assoc . 2021 16 февраля. 10 (4): e018799. [Медлайн]. [Полный текст].
Золер МЛ. Ожирение считается причиной диабета почти в половине случаев в США. Medscape Medical News . 2021 18 февраля [Полный текст].
Абдулла А., Амин Ф.А., Ханум Ф. и др. Оценка риска диабета 2 типа с использованием лет ожирения в современной популяции в рамках исследования Фрамингема. Glob Health Action . 2016. 9: 30421. [Медлайн].
Макколл Б.Новые гены помогают понять, почему многие люди с ожирением не болеют диабетом. Medscape Medical News . 2018 19 марта [Полный текст].
Евангелиста Л.С., Чо В.К., Ким Ю. Ожирение и хроническая болезнь почек: популяционное исследование среди южнокорейцев. PLoS One . 2018. 13 (2): e01
Martinelli CE, Keogh JM, Greenfield JR, Henning E, van der Klaauw AA, Blackwood A, et al. Ожирение из-за дефицита рецептора меланокортина 4 (MC4R) связано с повышенным линейным ростом и конечным ростом, гиперинсулинемией натощак и не полностью подавленной секрецией гормона роста. Дж. Клин Эндокринол Метаб . 2011, январь 96 (1): E181-8. [Медлайн].
Хэмди О. Роль жировой ткани как эндокринной железы. Curr Diab Rep . 2005 Октябрь 5 (5): 317-9. [Медлайн].
Бэйс Х., Блонд Л., Розенсон Р. Адипозопатия: как диета, упражнения и лекарственная терапия для снижения веса улучшают метаболические нарушения у пациентов с избыточным весом ?. Эксперт Rev Cardiovasc Ther . 2006 г., 4 (6): 871-95. [Медлайн].
Ketterer C, Heni M, Thamer C, Herzberg-Schäfer SA, Häring HU, Fritsche A.Острая краткосрочная гиперинсулинемия повышает обонятельный порог у здоровых людей. Инт Дж. Обес (Лондон) . 2011 Август 35 (8): 1135-8. [Медлайн].
Либ В., Салливан Л. М., Харрис Т. Б., Рубенофф Р., Бенджамин Э. Дж., Леви Д. и др. Уровни лептина в плазме и частота сердечной недостаточности, сердечно-сосудистых заболеваний и общая смертность у пожилых людей. Уход за диабетом . 2009 Апрель, 32 (4): 612-6. [Медлайн]. [Полный текст].
Биркетведт Г.С., Флорхольмен Дж., Сундсфьорд Дж., Остеруд Б., Дингес Д., Билкер В. и др.Поведенческие и нейроэндокринные характеристики синдрома ночного переедания. JAMA . 18 августа 1999 г. 282 (7): 657-63. [Медлайн].
Murray PG, Read A, Banerjee I, Whatmore AJ, Pritchard LE, Davies RA, et al. Снижение аппетита и индекса массы тела с задержкой полового созревания у матери и сына: связь с редким новым вариантом последовательности в гене лептина. Eur J Endocrinol . 2011 Апрель 164 (4): 521-7. [Медлайн].
Физическая активность: факты о физической активности.Центры по контролю и профилактике заболеваний. Доступно по адресу http://www.cdc.gov/physicalactivity/data/facts.html. Доступ: 9 января 2013 г.
Марипуу М., Викгрен М., Карлинг П., Адольфссон Р., Норрбак К.Ф. Относительный гипокортизолизм связан с ожирением и метаболическим синдромом при повторяющихся аффективных расстройствах. J Влияет на Disord . 2016, 21 июня. 204: 187-196. [Медлайн].
Tester JM, Phan TT, Tucker JM, et al. Характеристики детей от 2 до 5 лет с тяжелым ожирением. Педиатрия . 2018 27 февраля. [Medline].
Bouchard C, Tremblay A, Després JP, Nadeau A, Lupien PJ, Thériault G и др. Ответ на длительное перекармливание у однояйцевых близнецов. N Engl J Med . 1990, 24 мая. 322 (21): 1477-82. [Медлайн].
Freeman E, Fletcher R, Collins CE, et al. Профилактика и лечение детского ожирения: время обратиться к отцам. Инт Дж. Обес (Лондон) . 2012 января. 36 (1): 12-5. [Медлайн].
Chambers JC, Elliott P, Zabaneh D, Zhang W., Li Y, Froguel P, et al. Общие генетические вариации около MC4R связаны с окружностью талии и инсулинорезистентностью. Нат Генет . 2008 июн. 40 (6): 716-8. [Медлайн].
Frayling TM, Онг К. Собираем лобзик FTO. Биология генома . 2011. 12 (2): 104. [Медлайн]. [Полный текст].
Лоос Р.Дж., Линдгрен К.М., Ли С., Уиллер Э., Чжао Дж. Х., Прокопенко И. и др.Общие варианты, близкие к MC4R, связаны с жировой массой, весом и риском ожирения. Нат Генет . 2008 июн. 40 (6): 768-75. [Медлайн]. [Полный текст].
Scuteri A, Sanna S, Chen WM, Uda M, Albai G, Strait J и др. Полногеномное ассоциативное сканирование показывает, что генетические варианты в гене FTO связаны с признаками, связанными с ожирением. PLoS Genet . 2007 июл.3 (7): e115. [Медлайн]. [Полный текст].
Day FR, Loos RJ. Развитие генетики ожирения в эпоху полногеномных ассоциативных исследований. Дж Нутригенет Нутригеномикс . 2011. 4 (4): 222-38. [Медлайн].
Reinehr T, Kleber M, de Sousa G, et al. Концентрация лептина является прогностическим фактором снижения избыточной массы тела при изменении образа жизни. Int J Pediatr Obes . 13 мая 2009 г .; 1-9: [Medline].
Cummings DE, Schwartz MW. Меланокортины и масса тела: сказка о двух рецепторах. Нат Генет . 2000 Сентябрь 26 (1): 8-9. [Медлайн].
Vaisse C, Clement K, Durand E, Hercberg S, Guy-Grand B, Froguel P.Мутации рецептора меланокортина-4 являются частой и неоднородной причиной патологического ожирения. Дж. Клин Инвест . 2000 июл.106 (2): 253-62. [Медлайн]. [Полный текст].
Wardlaw SL. Клинический обзор 127: Ожирение как нейроэндокринное заболевание: уроки, которые следует извлечь из мутаций рецепторов проопиомеланокортина и меланокортина у мышей и мужчин. Дж. Клин Эндокринол Метаб . 2001 апр. 86 (4): 1442-6. [Медлайн].
Гибсон В.Т., Фаруки И.С., Моро М., ДеПаоли А.М., Лоуренс Э., О’Рахилли С. и др.Врожденный дефицит лептина из-за гомозиготности по мутации Delta133G: отчет о другом случае и оценка ответа на четырехлетнюю терапию лептином. Дж. Клин Эндокринол Метаб . 2004 Октябрь 89 (10): 4821-6. [Медлайн].
Abbasi A, Corpeleijn E, Postmus D, Gansevoort RT, de Jong PE, Gans RO и др. Прокальцитонин в плазме крови связан с ожирением, инсулинорезистентностью и метаболическим синдромом. Дж. Клин Эндокринол Метаб . 2010 сентябрь 95 (9): E26-31.[Медлайн].
Yaemsiri S, Slining MM, Agarwal SK. Воспринимаемый статус веса, диагноз избыточного веса и контроль веса среди взрослых в США: исследование NHANES 2003-2008. Инт Дж. Обес (Лондон) . 2011 Август 35 (8): 1063-70. [Медлайн].
Лайдман Дж. Причины ожирения: 1 из 5 смертей, связанных с избыточным весом. Медицинские новости Medscape. Доступно на http://www.medscape.com/viewarticle/809516. Доступ: 21 августа 2013 г.
Мастерс РК, Райтер ЭН, Пауэрс Д.А., Ян YC, Бургер А.Е., Линк Б.Г.Влияние ожирения на уровни смертности в США: важность возраста и факторов когорты в оценках населения. Am J Public Health . 2013 15 августа
Людвиг Дж., Санбонмацу Л., Геннетиан Л. и др. Окрестности, ожирение и диабет — рандомизированный социальный эксперимент. N Engl J Med . 2011 Октябрь 20, 365 (16): 1509-19. [Медлайн].
Молариус А., Сейделл Дж. К., Санс С., Туомилехто Дж., Кууласмаа К. Варьирующаяся чувствительность уровней действия талии для выявления субъектов с избыточным весом или ожирением в 19 популяциях проекта ВОЗ MONICA. J Clin Epidemiol . 1999 Декабрь 52 (12): 1213-24. [Медлайн].
Молариус А., Сейделл Дж. К., Санс С., Туомилехто Дж., Кууласмаа К. Окружность талии и бедер, а также соотношение талии и бедер в 19 популяциях проекта ВОЗ MONICA. Int J Obes Relat Metab Disord . 1999 23 февраля (2): 116-25. [Медлайн].
Сотрудничество по факторам риска НИЗ. Мировые тенденции индекса массы тела, недостаточного веса, избыточного веса и ожирения с 1975 по 2016 год: объединенный анализ 2416 популяционных исследований из 128.9 миллионов детей, подростков и взрослых. Ланцет . 2017.10.10. [Полный текст].
McCall B. Ожирение у детей растет в 10 раз; С недостаточным весом это «двойное бремя». Medscape Medical News . 11 октября 2017 г. [Полный текст].
Finucane MM, Стивенс GA, Cowan MJ, Danaei G, Lin JK, Paciorek CJ, et al. Национальные, региональные и глобальные тенденции индекса массы тела с 1980 года: систематический анализ обследований состояния здоровья и эпидемиологических исследований с участием 960 стран-лет и 9,1 миллиона участников. Ланцет . 2011 12 февраля. 377 (9765): 557-67. [Медлайн].
Neel СП. «Экономный генотип» 1998г. Нутр Рев . 1999 Май. 57 (5, часть 2): S2-9. [Медлайн].
Меткалф Б.С., Хоскинг Дж., Фремо А.Э., Джеффри А.Н., Фосс Л.Д., Уилкин Т.Дж. ИМТ всегда был прав: более высокие дети действительно толще (значение ИМТ в детстве не зависит от роста) EarlyBird 48. Int J Obes (Lond) . 2011 Апрель, 35 (4): 541-7. [Медлайн].
Н.С., Сучиндран С., Норт К.Э., Попкин Б.М., Гордон-Ларсен П. Ассоциация подросткового ожирения с риском тяжелого ожирения во взрослом возрасте. JAMA . 10 ноября 2010 г. 304 (18): 2042-7. [Медлайн]. [Полный текст].
Эллисон Д. Б., Фонтейн К. Р., Мэнсон Дж. Э., Стивенс Дж., Ван Италли ТБ. Ежегодные смерти от ожирения в США. JAMA . 1999, 27 октября. 282 (16): 1530-8. [Медлайн].
[Рекомендации] Группа экспертов по выявлению, оценке и лечению взрослых с избыточной массой тела.Клинические рекомендации по выявлению, оценке и лечению избыточного веса и ожирения у взрослых: резюме. Группа экспертов по выявлению, оценке и лечению избыточной массы тела у взрослых. Ам Дж. Клин Нутр . 1998 Октябрь 68 (4): 899-917. [Медлайн].
Брей GA. Опасность ожирения для здоровья. Endocrinol Metab Clin North Am . 1996 25 декабря (4): 907-19. [Медлайн].
Flegal KM, Graubard BI, Williamson DF, Gail MH.Чрезмерная смертность, связанная с недостаточной массой тела, избыточной массой тела и ожирением. JAMA . 2005 20 апреля. 293 (15): 1861-7. [Медлайн].
Чжэн В., Маклерран Д.Ф., Роллан Б., Чжан Х, Иноуэ М., Мацуо К. и др. Связь между индексом массы тела и риском смерти у более чем 1 миллиона азиатов. N Engl J Med . 2011 24 февраля. 364 (8): 719-29. [Медлайн].
Беррингтон де Гонсалес А., Хартге П., Серхан Дж. Р., Флинт А. Дж., Ханнан Л., Макиннис Р. Дж. И др.Индекс массы тела и смертность 1,46 миллиона взрослых белых. N Engl J Med . 2010 декабрь 2. 363 (23): 2211-9. [Медлайн]. [Полный текст].
Боггс Д.А., Розенберг Л., Козье Ю.К., Вайз Л.А., Куган П.Ф., Руис-Нарваез Е.А. и др. Общее и абдоминальное ожирение и риск смерти среди чернокожих женщин. N Engl J Med . 2011 сентябрь 8. 365 (10): 901-8. [Медлайн].
Юнг Дж. Х., Ан С. В., Сонг Дж. М. и др. Ожирение как фактор риска увеличения предстательной железы: ретроспективное когортное исследование в Корее. Int Neurourol J . 2016 20 декабря (4): 321-328. [Медлайн].
Stessman J, Jacobs JM, Ein-Mor E, Bursztyn M. Нормальный индекс массы тела, а не ожирение, предсказывает большую смертность среди пожилых людей: Иерусалимское лонгитюдное исследование. Дж. Ам Гериатр Соц . 2009 Декабрь 57 (12): 2232-8. [Медлайн].
Тамакоши А., Яцуя Х, Лин И, Тамакоши К., Кондо Т, Сузуки С. и др. ИМТ и смертность от всех причин среди пожилых людей в Японии: результаты совместного когортного исследования в Японии. Ожирение (Серебряная весна) . 2010 февраля 18 (2): 362-9. [Медлайн].
Wadden TA, Neiberg RH, Wing RR, Clark JM, Delahanty LM, Hill JO и др. Снижение веса за четыре года в исследовании Look AHEAD: факторы, связанные с долгосрочным успехом. Ожирение (Серебряная весна) . 2011 октября 19 (10): 1987-98. [Медлайн]. [Полный текст].
Эльбель Б., Гьямфи Дж., Керш Р. Выбор фаст-фуда для детей и подростков и влияние маркировки калорий: естественный эксперимент. Инт Дж. Обес (Лондон) . 2011 апр. 35 (4): 493-500. [Медлайн].
Waters E, de Silva-Sanigorski A, Hall BJ, et al. Вмешательства по профилактике ожирения у детей. Кокрановская база данных Syst Rev . 7 декабря 2011 г. 12: CD001871. [Медлайн].
Заявление Американской ассоциации клинических эндокринологов по использованию A1C для диагностики диабета. Доступно на http://emedicine.medscape.com/article/117853-workup. Доступ: 6 августа 2012 г.
[Рекомендации] Диагностика и классификация сахарного диабета. Уход за диабетом . 2010, январь 33, Приложение 1: S62-9. [Медлайн]. [Полный текст].
[Рекомендации] Дженсен, М.Д., Райан Д.Х., Аповиан С.М. и др. Руководство AHA / ACC / TOS, 2013 г., по лечению избыточной массы тела и ожирения у взрослых: доклад Целевой группы Американского колледжа кардиологов / Американской кардиологической ассоциации по практическим рекомендациям и Общества ожирения. Дж. Ам Кол Кардиол .2013. [Medline]. [Полный текст].
Наингголан Л. Новые рекомендации по ожирению: авторитетная «дорожная карта» лечения. Medscape Medical News . 12 ноября 2013 г. [Полный текст].
Такер МЭ. Новые рекомендации США по ожирению. Сначала обработайте вес. Медицинские новости Medscape. Доступно на http://www.medscape.com/viewarticle/838285.
Apovian CM, Aronne LJ, Bessesen DH, et al. Фармакологическое лечение ожирения: руководство по клинической практике эндокринного общества. Дж. Клин Эндокринол Метаб . 2015 Февраль 100 (2): 342-62. [Медлайн].
Джолли К., Льюис А., Бич Дж. И др. Сравнение ряда коммерческих программ или программ первичной медико-санитарной помощи по снижению веса с минимальным вмешательством для контроля потери веса при ожирении: рандомизированное контролируемое исследование Lighten Up. BMJ . 2011 г. 3 ноября. 343: d6500. [Медлайн]. [Полный текст].
Брей GA. Лекарства для похудания. Med Clin North Am .2011 сентябрь 95 (5): 989-1008. [Медлайн].
Wing RR, Lang W., Wadden TA, Safford M, Knowler WC, Bertoni AG и др. Преимущества умеренной потери веса в снижении факторов риска сердечно-сосудистых заболеваний у людей с избыточным весом и ожирением с диабетом 2 типа. Уход за диабетом . 2011 июл. 34 (7): 1481-6. [Медлайн]. [Полный текст].
Столли М.Р., Фитцгиббон М.Л., Шиффер Л., Шарп Л.К., Сингх В., Ван Хорн Л. и др. Исследование снижения ожирения черным (ORBIT): результаты за шесть месяцев. Ожирение (Серебряная весна) . 2009 17 января (1): 100-6. [Медлайн].
Ларсен TM, Далсков С.М., ван Баак М., Джебб С.А., Пападаки А., Пфайффер А.Ф. и др. Диеты с высоким или низким содержанием белка и гликемическим индексом для поддержания потери веса. N Engl J Med . 2010 25 ноября. 363 (22): 2102-13. [Медлайн]. [Полный текст].
Блюхер М., Рудич А., Клотинг Н. и др. Две модели динамики адипокина и других биомаркеров при длительном вмешательстве по снижению веса. Уход за диабетом . 2012 Февраль 35 (2): 342-9. [Медлайн]. [Полный текст].
Сумитран П., Прендергаст Л.А., Делбридж Э. и др. Длительное сохранение гормональных адаптаций к похуданию. N Engl J Med . 2011 27 октября. 365 (17): 1597-604. [Медлайн].
Хэмди О., Мотталиб А., Морси А. и др. Долгосрочное влияние интенсивного изменения образа жизни на факторы риска сердечно-сосудистых заболеваний у пациентов с диабетом в реальной клинической практике: 5-летнее продольное исследование. BMJ Open Diabetes Res Care . 2017. 5: e000259. [Полный текст].
Маффейс К. Детское ожирение: генетический интерфейс и среда. Baillieres Best Practices Clin Endocrinol Metab . 1999 г., 13 (1): 31-46. [Медлайн].
Проимос Дж., Сойер С. Ожирение в детском и подростковом возрасте. Врач Aust Fam . 2000 апреля, 29 (4): 321-7. [Медлайн].
Харша Д.В., Брей, Джорджия. Состав тела и детское ожирение. Endocrinol Metab Clin North Am . 1996 25 декабря (4): 871-85. [Медлайн].
Clement K, van den Akker E, Argente J, et al. Эффективность и безопасность сетмеланотида, агониста MC4R, у лиц с тяжелым ожирением из-за дефицита LEPR или POMC: одноранговые, открытые, многоцентровые испытания, фаза 3. Ланцет, диабет, эндокринол . 2020 Декабрь 8 (12): 960-70. [Медлайн].
Imcivree (сетмеланотид) [вставка в упаковку]. Бостон, Массачусетс: Rhythm Pharmaceuticals, Inc.Ноябрь 2020 г. Доступно в [Полный текст].
Пожилые и пожилые люди. В: Потребности человека в энергии: Отчет совместной консультации экспертов ФАО / ВОЗ / УООН. Рим, 17-24 октября 2001 г. Продовольственная и сельскохозяйственная организация ООН. Доступно по адресу http://www.fao.org/docrep/007/y5686e/y5686e09.htm#bm9.
Brooks M. Стандартное значение «1-MET» недействительно при избыточном весе / ожирении. Медицинские новости Medscape. Доступно на http://www.medscape.com/viewarticle/821375.Доступ: 10 марта 2014 г.
Вильмс Б., Эрнст Б., Турнхеер М., Вайссер Б., Шультес Б. Поправочные коэффициенты для расчета метаболических эквивалентов (МЕТ) при избыточном весе у субъектов с крайне ожирением. Инт Дж. Обес (Лондон) . 2014 7 февраля [Medline].
Foster GD, Wyatt HR, Hill JO, Makris AP, Rosenbaum DL, Brill C и др. Вес и метаболические исходы через 2 года на диете с низким содержанием углеводов в сравнении с диетой с низким содержанием жиров: рандомизированное исследование. Энн Интерн Мед. .2010 3 августа. 153 (3): 147-57. [Медлайн]. [Полный текст].
Shai I, Schwarzfuchs D, Henkin Y, Shahar DR, Witkow S, Greenberg I, et al. Похудение с помощью низкоуглеводной, средиземноморской или обезжиренной диеты. N Engl J Med . 17 июля 2008 г., 359 (3): 229-41.
Dansinger ML, Gleason JA, Griffith JL, Selker HP, Schaefer EJ. Сравнение диет Аткинса, Орниша, Weight Watchers и Zone для снижения веса и снижения риска сердечных заболеваний: рандомизированное исследование. JAMA . 2005, 5 января. 293 (1): 43-53. [Медлайн].
Очень низкокалорийные диеты. Наркотик Тер Булл . 2012 май. 50 (5): 54-7. [Медлайн].
Van Nieuwenhove Y, Dambrauskas Z, Campillo-Soto A, van Dielen F, Wiezer R, Janssen I, et al. Предоперационная низкокалорийная диета и исход операции после лапароскопического обходного желудочного анастомоза: рандомизированное многоцентровое исследование. Arch Surg . 2011 ноябрь 146 (11): 1300-5. [Медлайн].
Деннис Э.А., Денго А.Л., Комбер Д.Л., Флэк К.Д., Савла Дж., Дэви К.П. и др.Потребление воды увеличивает потерю веса во время гипокалорийной диеты у людей среднего и пожилого возраста. Ожирение (Серебряная весна) . 2010 февраля 18 (2): 300-7. [Медлайн]. [Полный текст].
Дубнов-Раз Г, Константини Н. В., Ярив Х, Ницца С., Шапира Н. Влияние питья воды на расход энергии в покое у детей с избыточным весом. Инт Дж. Обес (Лондон) . 2011 окт. 35 (10): 1295-300. [Медлайн].
Wiesner S, Haufe S, Engeli S, Mutschler H, Haas U, Luft FC и др.Влияние тренировок с нормобарической гипоксией на физическую подготовку и маркеры метаболического риска при избыточной массе тела у субъектов с ожирением. Ожирение (Серебряная весна) . 2010 января 18 (1): 116-20. [Медлайн].
Jakicic JM, Marcus BH, Lang W, Janney C. Влияние физических упражнений на поддержание потери веса в течение 24 месяцев у женщин с избыточным весом. Arch Intern Med . 28 июля 2008 г. 168 (14): 1550-9; обсуждение 1559-60. [Медлайн]. [Полный текст].
Ballor DL, Poehlman ET.Физические упражнения способствуют сохранению безжировой массы во время похудания, вызванного диетой: метааналитический результат. Int J Obes Relat Metab Disord . 1994 18 января (1): 35-40. [Медлайн].
Villareal DT, Chode S, Parimi N, Sinacore DR, Hilton T, Armamento-Villareal R, et al. Потеря веса, физические упражнения или и то, и другое, а также физическая функция у пожилых людей с ожирением. N Engl J Med . 2011 31 марта. 364 (13): 1218-29. [Медлайн]. [Полный текст].
Goodpaster BH, Delany JP, Otto AD, Kuller L, Vockley J, South-Paul JE, et al.Влияние диет и вмешательств на физическую активность на факторы риска потери веса и кардиометаболизма у взрослых с тяжелым ожирением: рандомизированное исследование. JAMA . 27 октября 2010 г. 304 (16): 1795-802. [Медлайн]. [Полный текст].
Хэнкинсон А.Л., Давиглус М.Л., Бушар С., Карнетон М., Льюис К.Э., Шрайнер П.Дж. и др. Поддержание высокого уровня физической активности более 20 лет и увеличение веса. JAMA . 2010 15 декабря. 304 (23): 2603-10. [Медлайн].
Rejeski WJ, Brubaker PH, Goff DC Jr, Bearon LB, McClelland JW, Perri MG, et al.Внедрение программ снижения веса и физической активности в общину для сохранения подвижности пожилых людей с ожирением и плохим сердечно-сосудистым здоровьем. Arch Intern Med . 2011 г. 23 мая. 171 (10): 880-6. [Медлайн].
Ван Дорстен Б., Линдли Е.М. Когнитивные и поведенческие подходы в лечении ожирения. Med Clin North Am . 2011 Сентябрь 95 (5): 971-88. [Медлайн].
Морган П.Дж., Любанс Д.Р., Каллистер Р., Окели А.Д., Берроуз Т.Л., Флетчер Р. и др.Рандомизированное контролируемое исследование «Здоровые папы, здоровые дети»: эффективность программы здорового образа жизни для отцов с избыточным весом и их детей. Инт Дж. Обес (Лондон) . 2011 марта, 35 (3): 436-47. [Медлайн].
Mozaffarian D, Hao T., Rimm EB, Willett WC, Hu FB. Изменения в диете и образе жизни и длительное увеличение веса у женщин и мужчин. N Engl J Med . 23 июня 2011 г. 364 (25): 2392-404. [Медлайн]. [Полный текст].
Недельчева А.В., Килкус Ю.М., Империал Дж., Шоллер Д.А., Пенев П.Д.Недостаток сна подрывает диетические усилия по снижению ожирения. Энн Интерн Мед. . 2010 Октябрь 5. 153 (7): 435-41. [Медлайн]. [Полный текст].
FDA распространяет предупреждение для потребителей о испорченных таблетках для похудания. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. 8 января 2009 г. Доступно по адресу http://www.fda.gov/newsevents/newsroom/pressannouncements/2008/ucm116998.htm. Доступ: январь 2013 г.
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. BMPEA в диетических добавках.Доступно на http://www.fda.gov/Food/DietarySupplements/QADietarySupplements/ucm443790.htm. Дата обращения: 27 апреля 2015 г.
Буско М. FDA одобрило семаглутид «Gamechanger» для похудания. Medscape Medical News . 2021 г. 4 июня [Полный текст].
Рубино Д., Абрахамссон Н., Дэвис М. и др. Эффект продолжающегося еженедельного подкожного введения семаглутида по сравнению с плацебо на поддержание потери веса у взрослых с избыточной массой тела или ожирением: рандомизированное клиническое испытание ШАГА 4. JAMA . 2021 13 апреля. 325 (14): 1414-25. [Медлайн].
Wilding JPH, Баттерхэм Р.Л., Каланна С. и др. Семаглутид один раз в неделю для взрослых с избыточным весом или ожирением. N Engl J Med . 2021 18 марта. 384 (11): 989. [Медлайн].
Пресс-релиз FDA. FDA одобрило препарат Saxenda для контроля веса. Доступно на http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm427913.htm. Доступ: 23 декабря 2014 г.
Хек А.М., Яновский Ю.А., Калис К.А.Орлистат, новый ингибитор липазы для лечения ожирения. Фармакотерапия . 2000 марта 20 (3): 270-9. [Медлайн].
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. FDA требует отозвать с рынка препарат для похудания Belviq, Belviq XR (лорказерин). FDA. Доступно на https://www.fda.gov/drugs/drug-safety-and-availability/fda-requests-withdrawal-weight-loss-drug-belviq-belviq-xr-lorcaserin-market?utm_campaign=FDA%20requests% 20% 20при отмене% 20% 20% 20 потеря веса% 20% 20% 20% 20% 2C% 20% 20XR% 20% 28Лорказерин% 29 &.13 февраля 2020 г .; Доступ: 14 февраля 2020 г.
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. FDA одобряет Belviq для лечения некоторых взрослых с избыточным весом или ожирением. 27 июня 2012 г. Доступно по адресу http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm309993.htm. Доступ: 12 июля 2012 г.
Списки контролируемых веществ: включение лорказерина в Список IV. Отдел по борьбе с наркотиками. Доступно по адресу http://www.deadiversion.usdoj.gov/fed_regs/rules/2012/fr1219.htm. Доступ: 28 декабря 2012 г.
Smith SR, Weissman NJ, Anderson CM, Sanchez M, Chuang E, Stubbe S, et al. Многоцентровое плацебо-контролируемое исследование лорказерина для контроля веса. N Engl J Med . 15 июля 2010 г. 363 (3): 245-56. [Медлайн]. [Полный текст].
Фидлер М.К., Санчес М., Рэтер Б., Вайсман Н.Дж., Смит С.Р., Шанахан В.Р. и др. Годичное рандомизированное исследование лорказерина для снижения веса у взрослых с ожирением и избыточным весом: исследование BLOSSOM. Дж. Клин Эндокринол Метаб . 2011 Октябрь 96 (10): 3067-77. [Медлайн].
O’Neil PM, Smith SR, Weissman NJ, Fidler MC, Sanchez M, Zhang J, et al. Рандомизированное плацебо-контролируемое клиническое исследование лорказерина для снижения веса при сахарном диабете 2 типа: исследование BLOOM-DM. Ожирение (Серебряная весна) . 2012 июл.20 (7): 1426-36. [Медлайн].
Серретти А., Манделли Л. Антидепрессанты и масса тела: всесторонний обзор и метаанализ. J Clin Psychiatry . 2010 Октябрь 71 (10): 1259-72. [Медлайн].
Goldfield GS, Lorello C, Doucet E. Метилфенидат снижает потребление энергии и потребление жиров с пищей у взрослых: механизм снижения подкрепляющей ценности пищи ?. Ам Дж. Клин Нутр . 2007 августа 86 (2): 308-15. [Медлайн].
Gadde KM, Franciscy DM, Wagner HR 2nd, Krishnan KR. Зонисамид для снижения веса у взрослых с ожирением: рандомизированное контролируемое исследование. JAMA .2003, 9 апреля. 289 (14): 1820-5. [Медлайн].
Lustig RH, Hinds PS, Ringwald-Smith K, Christensen RK, Kaste SC, Schreiber RE, et al. Октреотидная терапия детского гипоталамического ожирения: двойное слепое плацебо-контролируемое исследование. Дж. Клин Эндокринол Метаб . 2003 июнь 88 (6): 2586-92. [Медлайн].
Desilets AR, Dhakal-Karki S, Dunican KC. Роль метформина для контроля веса у пациентов без диабета 2 типа. Энн Фармакотер .2008 июн. 42 (6): 817-26. [Медлайн].
Vilsbøll T, Christensen M, Junker AE, Knop FK, Gluud LL. Влияние агонистов рецепторов глюкагоноподобного пептида-1 на потерю веса: систематический обзор и метаанализ рандомизированных контролируемых исследований. BMJ . 2012, 10 января. 344: d7771. [Медлайн]. [Полный текст].
Gadde KM, Xiong GL. Бупропион для похудания. Эксперт Rev Neurother . 2007 7 января (1): 17-24. [Медлайн].
Черный SC.Антагонисты каннабиноидных рецепторов и ожирение. Curr Opin Investigations Drugs . 2004 г., 5 (4): 389-94. [Медлайн].
Van Gaal LF, Rissanen AM, Scheen AJ, et al. Влияние блокатора рецепторов каннабиноида-1 римонабанта на снижение веса и факторы риска сердечно-сосудистых заболеваний у пациентов с избыточной массой тела: 1-летний опыт исследования RIO-Europe. Ланцет . 16-22 апреля 2005 г. 365 (9468): 1389-97.
Cox SL. Римонабанта гидрохлорид: исследуемый агент для лечения сердечно-сосудистых факторов риска. Наркотики сегодня (Barc) . 2005 г., 41 (8): 499-508. [Медлайн].
Фернандес Дж. Р., Эллисон ДБ. Римонабант Санофи-Синтелабо. Curr Opin Investigations Drugs . 2004 г., 5 (4): 430-5. [Медлайн].
Нагао Т., Мегуро С., Хасэ Т., Оцука К., Комикадо М., Токимицу И. и др. Напиток, богатый катехинами, улучшает контроль за ожирением и уровнем глюкозы в крови у пациентов с диабетом 2 типа. Ожирение (Серебряная весна) . 2009 17 февраля (2): 310-7.[Медлайн].
Дуникан К.С., Адамс Н.М., Десилетс АР. Роль прамлинтида в похудании. Энн Фармакотер . 2010 Март 44 (3): 538-45. [Медлайн].
Баттерхэм Р.Л., Коэн М.А., Эллис С.М., Ле Ру С.В., Уизерс DJ, Фрост Г.С. и др. Подавление приема пищи у пациентов с ожирением пептидом YY3-36. N Engl J Med . 2003 сентябрь 4. 349 (10): 941-8. [Медлайн].
Боггиано М.М., Чандлер П.К., Освальд К.Д., Роджерс Р.Дж., Бланделл Дж.Э., Исии Ю.PYY3-36 в качестве мишени для лечения ожирения. Obes Ред. . 2005 г., 6 (4): 307-22. [Медлайн].
Roth CL, Enriori PJ, Harz K, Woelfle J, Cowley MA, Reinehr T. Пептид YY является регулятором энергетического гомеостаза у детей с ожирением до и после потери веса. Дж. Клин Эндокринол Метаб . 2005 декабрь 90 (12): 6386-91. [Медлайн].
Moon HS, Matarese G, Brennan AM, Chamberland JP, Liu X, Fiorenza CG, et al. Эффективность метрелептина у пациентов с ожирением и диабетом 2 типа: клеточные и молекулярные пути, лежащие в основе толерантности к лептину. Диабет . 2011 июн. 60 (6): 1647-56. [Медлайн]. [Полный текст].
Ravussin E, Smith SR, Mitchell JA, Shringarpure R, Shan K, Maier H, et al. Повышенная потеря веса с помощью прамлинтида / метрелептина: интегрированный нейрогормональный подход к фармакотерапии ожирения. Ожирение (Серебряная весна) . 2009 Сентябрь 17 (9): 1736-43. [Медлайн]. [Полный текст].
Tam CS, Lecoultre V, Ravussin E. Новая стратегия использования лептина для терапии ожирения. Экспертное мнение Biol Ther . 2011 Декабрь 11 (12): 1677-85. [Медлайн].
Sjöström L, Peltonen M, Jacobson P, Sjöström CD, Karason K, Wedel H, et al. Бариатрическая хирургия и отдаленные сердечно-сосудистые события. JAMA . 2012 г. 4 января. 307 (1): 56-65. [Медлайн].
Flum DR, Belle SH, King WC, Wahed AS, Berk P, Chapman W. и др. Периоперационная безопасность в продольной оценке бариатрической хирургии. N Engl J Med . 2009 30 июля.361 (5): 445-54. [Медлайн]. [Полный текст].
Маггард М.А., Шугарман Л.Р., Сатторп М., Маглион М., Сугерман Х.Дж., Ливингстон Э.Х. и др. Метаанализ: хирургическое лечение ожирения. Энн Интерн Мед. . 2005 Apr 5. 142 (7): 547-59. [Медлайн].
Такер МЭ. Новые рекомендации по бариатрической хирургии отражают быстро развивающуюся область. Медицинские новости Medscape. 28 марта 2013 г. Доступно по адресу http://www.medscape.com/viewarticle/781619. Доступ: 3 апреля 2013 г.
Фиоре К.Новые рекомендации по обновлению рукава после операции по снижению веса. MedPage сегодня. Доступно по адресу http://www.medpagetoday.com/Endocrinology/Obesity/38112?utm_content=&utm_medium=email&utm_campaign=DailyHeadlines&utm_source=WC&xid=NL_DHE_2013-03-28&eun=g648601d6d03&use=g648601d1d03d03&eun=g648601d6d03&use=g648601d05d05d05d06d06d06e6 Доступ: 3 апреля 2013 г.
Mechanick JI, Youdim A, Jones DB, Garvey WT, Hurley DL, McMahon MM, et al. Руководство по клинической практике для периоперационной нутритивной, метаболической и нехирургической поддержки пациента, перенесшего бариатрическую операцию — обновление 2013 г.: спонсировано Американской ассоциацией клинических эндокринологов, Обществом ожирения и Американским обществом метаболической и бариатрической хирургии. Endocr Pract . 2013 25 марта. E1-e36. [Медлайн].
Эшли С., Берд Д.Л., Сагден Дж., Ройстон С.М. Вертикально-полосатая гастропластика для лечения патологического ожирения. Br J Surg . 1993 ноябрь 80 (11): 1421-3. [Медлайн].
Фликингер Э.Г., Порис В.Дж., Мелхейм HD, Синар Д.Р., Блоуз Иллинойс, Томас Ф.Т. Обходной желудочный анастомоз по Гринвиллу. Отчет о проделанной работе за 3 года. Энн Сург . 1984 Май. 199 (5): 555-62. [Медлайн]. [Полный текст].
Плекка Остлунд М., Марск Р., Расмуссен Ф., Лагергрен Дж., Нэслунд Э. Заболеваемость и смертность до и после бариатрической операции по поводу патологического ожирения по сравнению с населением в целом. Br J Surg . 2011 июн. 98 (6): 811-6. [Медлайн].
Mingrone G, Panunzi S, De Gaetano A, Guidone C, Iaconelli A, Leccesi L, et al. Бариатрическая хирургия в сравнении с традиционной медикаментозной терапией диабета 2 типа. N Engl J Med . 2012 26 апреля.366 (17): 1577-85. [Медлайн]. [Полный текст].
Søvik TT, Aasheim ET, Taha O, Engström M, Fagerland MW, Björkman S, et al. Потеря веса, факторы риска сердечно-сосудистых заболеваний и качество жизни после желудочного обходного анастомоза и двенадцатиперстной кишки: рандомизированное исследование. Энн Интерн Мед. . 2011 6 сентября. 155 (5): 281-91. [Медлайн].
Hedberg J, Sundbom M. Превосходная потеря веса и более низкий уровень HbA1c через 3 года после переключения на двенадцатиперстную кишку по сравнению с желудочным обходным анастомозом Roux-en-Y — рандомизированное контролируемое исследование. Surg Obes Relat Dis . 2012 май-июнь. 8 (3): 338-43. [Медлайн].
Schiavon CA, Bersch-Ferreira AC, Santucci EV, et al. Эффекты бариатрической хирургии у пациентов с ожирением и гипертонией: рандомизированное исследование GATEWAY (желудочный обходной анастомоз для лечения пациентов с ожирением и устойчивой гипертензией). Тираж . 2017 г. 13 ноября [Medline].
Стайлз С. Бариатрическая хирургия может привести к «ремиссии» АГ при ожирении: GATEWAY. Medscape .2017 г. 13 ноября [Полный текст].
Цигайна В. Желудочная кардиостимуляция как терапия патологического ожирения: предварительные результаты. Obes Surg . 2002 г., 12 апреля, приложение 1: 12С-16С. [Медлайн].
Кляйн С., Фонтана Л., Янг В. Л. и др. Отсутствие влияния липосакции на действие инсулина и факторы риска ишемической болезни сердца. N Engl J Med . 17 июня 2004 г. 350 (25): 2549-57.
Koch TR, Finelli FC. Послеоперационные метаболические и пищевые осложнения бариатрической хирургии. Гастроэнтерол Clin North Am . 2010 марта 39 (1): 109-24. [Медлайн].
Powell-Wiley TM, Poirier P, Burke LE, et al. Ожирение и сердечно-сосудистые заболевания: научное заявление Американской кардиологической ассоциации. Тираж . 2021, 22 апреля. CIR0000000000000973. [Медлайн]. [Полный текст].
Киркнер РМ. В заявлении AHA об ожирении подчеркивается абдоминальный жир, AFib. MDedge . 2021 22 апреля [Полный текст].
Мунирадж Т., Дэй Л.В., Тейген Л.М. и др.Руководство AGA по клинической практике использования внутрижелудочных баллонов при лечении ожирения. Гастроэнтерология . 2021 апр. 160 (5): 1799-1808. [Медлайн]. [Полный текст].
Karon A. AGA Руководство по клинической практике: внутрижелудочные баллоны в лечении ожирения. MDedge . 2021 16 апреля [Полный текст].
Flegal KM, Carroll MD, Kit BK, Ogden CL. Распространенность ожирения и тенденции в распределении индекса массы тела среди взрослого населения США, 1999-2010 гг. JAMA . 2012 г., 1 февраля. 307 (5): 491-7. [Медлайн].
Огден КЛ, Кэрролл Мэриленд, Кит БК, Флегал КМ. Распространенность ожирения и тенденции изменения индекса массы тела среди детей и подростков в США, 1999-2010 гг. JAMA . 2012 г., 1 февраля. 307 (5): 483-90. [Медлайн].
Abbott Laboratories соглашается отозвать свой препарат для лечения ожирения Meridia. FDA, Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. Доступно по адресу http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm228812.htm. Доступ: 8 октября 2010 г.
Андерсон Дж. У., Джавери, Массачусетс. Сокращение приема лекарств при значительной потере веса с поведенческим вмешательством. Curr Clin Pharmacol . 2010 г., 5 (4): 232-8. [Медлайн].
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов. FDA одобрило препарат Qsymia для контроля веса. Доступно на http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm312468.htm. Доступ: 7 августа 2012 г.
Grudell AB, Sweetser S, Camilleri M, Eckert DJ, Vazquez-Roque MI, Carlson PJ, et al.Контролируемое фармакогенетическое исследование сибутрамина по снижению веса и составу тела у взрослых с ожирением или избыточным весом. Гастроэнтерология . 2008 Октябрь 135 (4): 1142-54. [Медлайн]. [Полный текст].
Джеймс В.П., Катерсон ИД, Коутиньо В., Файнер Н., Ван Гаал Л.Ф., Маггиони А.П. и др. Влияние сибутрамина на сердечно-сосудистые исходы у лиц с избыточным весом и ожирением. N Engl J Med . 2010 сентябрь 2. 363 (10): 905-17. [Медлайн].
Лайдман Дж.Пороги ожирения позволяют точно прогнозировать риск для здоровья подростков. Medscape Medical News . 29 января 2014 г. [Полный текст].
Лаурсон К.Р., Велк Г.Дж., Эйзенманн Дж.С. Диагностическая эффективность процентилей ИМТ для выявления подростков с метаболическим синдромом. Педиатрия . 2014 февраль 133 (2): e330-8. [Медлайн].
Makowski CT, Gwinn KM, Hurren KM. Налтрексон / бупропион: исследуемая комбинация для похудания и поддержания веса. Obes Facts . 2011. 4 (6): 489-94. [Медлайн].
Nainggolan L. Желудочный бандаж — это операция первого шага для подростков с болезненным ожирением. Medscape Medical News . 29 мая 2014 г. [Полный текст].
Nainggolan L. FDA одобрило использование бупропиона / налтрексона (Contrave) для лечения ожирения. Медицинские новости Medscape. Доступно по адресу http://www.staging.medscape.com/viewarticle/831513. Доступ: 14 сентября 2014 г.
Schauer PR, Kashyap SR, Wolski K, Brethauer SA, Kirwan JP, Pothier CE, et al.Бариатрическая хирургия в сравнении с интенсивной медикаментозной терапией у пациентов с ожирением и диабетом. N Engl J Med . 2012 26 апреля. 366 (17): 1567-76. [Медлайн]. [Полный текст].
Sjostrom L, Narbro K, Sjostrom CD, et al. Влияние бариатрической хирургии на смертность у шведских пациентов с ожирением. N Engl J Med . 2007 23 августа. 357 (8): 741-52. [Медлайн].
Динамика роста жировой ткани
Abstract
Рост жировой ткани происходит по двум механизмам: гиперплазия (увеличение количества клеток) и гипертрофия (увеличение размера клеток).Генетика и диета влияют на относительный вклад этих двух механизмов в рост жировой ткани при ожирении. В этом исследовании распределение по размеру эпидидимальных жировых клеток от двух линий мышей, устойчивых к ожирению FVB / N и склонных к ожирению C57BL / 6, измеряли через 2, 4 и 12 недель в условиях регулярного кормления и питания с высоким содержанием жира. Общее количество клеток в жировой подушке придатка яичка оценивали по массе жировой подушечки и нормализованному распределению клеток по размеру. Число клеток и средневзвешенный размер клеток увеличиваются в зависимости от массы жировой прослойки.Чтобы точно решить проблему роста жировой ткани, мы разработали математическую модель, описывающую эволюцию распределения размеров жировых клеток как функцию увеличения массы жировой ткани, а не увеличения хронологического времени. Наша модель описывает привлечение новых жировых клеток и их последующее развитие в разных штаммах и с разными режимами питания, с общими механизмами, но с параметрами модели, зависящими от диеты и генетики. По сравнению со штаммом FVB / N, штамм C57BL / 6 имеет большее количество мелких жировых клеток.Гиперплазия усиливается за счет диеты с высоким содержанием жиров в зависимости от штамма, что предполагает синергетическое взаимодействие между генетикой и диетой. Кроме того, кормление с высоким содержанием жиров увеличивает скорость роста жировых клеток, независимо от напряжения, что отражает увеличение количества калорий, требующих хранения. Кроме того, диета с высоким содержанием жиров приводит к резкому увеличению распределения жировых клеток по размерам у обоих штаммов; это означает увеличение колебаний размера жировых клеток за счет липидного обмена.
Авторское резюме
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии.Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. Динамическое изменение жировой ткани in vivo невозможно отслеживать в режиме реального времени из-за текущих технических ограничений. Однако мы можем измерить распределение жировых клеток по размеру у отдельных животных. Наша основная цель — извлечь динамические характеристики ремоделирования тканей из снимков распределения клеток по размерам. Мы разрабатываем математическую модель, которая интерполирует измерения распределения клеток по размеру и предсказывает непрерывное изменение распределения клеток по размеру в зависимости от увеличения массы жировых подушечек.Наша модель роста жировой ткани включает три основных компонента: рекрутирование новых клеток, рост клеток в зависимости от размера и колебания размера клеток. В частности, мы сравнили рост жировой ткани у склонных к ожирению и устойчивых к ожирению мышей при стандартной диете или диете с высоким содержанием жиров, чтобы изучить генетическое и диетическое влияние на рост жировой ткани. Применяя нашу модель к этим различным условиям, мы обнаружили, что увеличение размера жировых клеток зависит от диеты. С другой стороны, увеличение количества жировых клеток, вызванное диетой, зависит от деформации, что предполагает синергию между генетикой и диетой.
Введение
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. На фенотип ожирения жировой ткани влияют диета и генетика, а также их взаимодействие [1] — [4]. Начиная с исследований Джонсона и Хирша [5], существует обширная литература о росте жировой ткани при нормальном и патологическом развитии, характеризующая состояние ткани с точки зрения среднего размера и количества клеток.Гиперпластический рост появляется только на ранних стадиях развития жировой ткани [6], [7]. Гипертрофия возникает до гиперплазии, чтобы удовлетворить потребность в дополнительных накоплениях жира при прогрессировании ожирения [8]. Однако оказалось трудно понять, как диета и генетика конкретно влияют на гиперплазию и / или гипертрофию жировых клеток из-за ограниченных данных о росте жировой ткани.
Помимо этих исследований, недавно стало возможным точно измерить распределение клеток по размерам.Эта подробная информация, по сравнению со средним размером клеток и общим числом клеток, может использоваться для вычисления многих величин, связанных с размером, которые позволяют более точно охарактеризовать процесс роста жировой ткани. Кумулянты распределения клеток по размерам могут использоваться для вычисления других физиологических величин, таких как средневзвешенный размер клеток. Распределение клеток по размерам можно использовать для оценки общего количества клеток в жировой подушке по ее массе. Кроме того, считается, что некоторые специфические метаболические свойства, например.например, инсулинорезистентность [9] и секреция адипокина [10] зависят от точного распределения клеток по размерам, а не от среднего размера клеток. Действительно, в нескольких исследованиях изучается изменение распределения жировых клеток по размерам в различных условиях при развитии куриного эмбриона [11], у худых и страдающих ожирением крыс Zucker [12], [13], крыс линии Wistar с частичной липэктомией [14], биопсии кролика [ 15] и жировой ткани человека [16], [17]. Эти исследования были сосредоточены только на статических различиях между распределениями клеток по размерам в разных условиях.Однако статические распределения размеров клеток в поперечном сечении для ряда снимков развития животных можно использовать для вывода динамики роста жировой ткани, если мы сможем надлежащим образом проанализировать снимки с помощью математического моделирования. Учитывая нынешние технические ограничения, это может быть лучший доступный подход к микроскопическому и продольному пониманию роста жировой ткани in vivo , хотя в недавнем эксперименте были получены микроскопические наблюдения накопления липидов в липидных каплях жировых клеток [18].
Для изучения генетического и диетического влияния на динамический процесс роста жировой ткани мы получили распределения по размерам клеток эпидидимального жира у устойчивых к ожирению линий мышей FVB / N (далее FVB) и склонных к ожирению линий мышей C57BL / 6 (C57) в соответствии с стандартные диеты и диеты с высоким содержанием жиров. Мышь C57 является наиболее охарактеризованной моделью ожирения, вызванного диетой [19], а мышь FVB является предпочтительной моделью для создания трансгенных мышей [20]. Эти два широко используемых инбредных штамма мышей генетически довольно далеки [21], [22] и имеют различные метаболические фенотипы: по сравнению с мышами FVB, мыши C57 имеют низкие уровни циркулирующих триглицеридов [21] и повышенный клиренс триглицеридов [23], [24]; Мыши FVB характеризуются относительно более высокой печеночной инсулинорезистентностью, контррегуляторной реакцией на гипогликемию и сниженной глюкозо-стимулированной секрецией инсулина [25]; Также известно, что мыши FVB являются спонтанно гиперактивными [26] и относительно худыми, поскольку они, по-видимому, менее восприимчивы к диете с высоким содержанием жиров, чем мыши C57 [27].Однако развитие ожирения, вызванного диетой, у этих двух штаммов формально не сравнивалось. В этом исследовании мы разработали математическую модель, прогнозирующую изменение распределения клеток по размерам в зависимости от массы жировой подушечки придатка яичка, для количественного анализа динамических характеристик, зависящих от генетики и / или диеты. Модель роста жировой ткани описывает, сколько новых клеток образуется, как каждая клетка растет в зависимости от ее размера и как липидный обмен приводит к колебаниям размера, которые вызывают расширение распределения клеток по размерам.По мере увеличения массы эпидидимальных жировых подушек распределение клеток по размерам систематически изменяется в зависимости как от генетики, так и от диеты. Сравнивая экспериментальные результаты с теоретической моделью роста, мы обнаружили, что гипертрофия сильно коррелирует с диетой. С другой стороны, гиперплазия зависит от генетики. Изменения при гиперплазии, вызванные диетой, зависят от штамма, что предполагает взаимодействие между диетой и генетикой.
Результаты
Влияние диеты с высоким содержанием жиров на состав тела мышей FVB и C57
В начале эксперимента (в возрасте 5 недель) мыши C57 были значительно легче, чем мыши FVB () из-за разницы в худой масса, хотя общая жировая масса не отличалась ().Когда мышей поддерживали на обычном питании, разница в массе тела исчезла к возрасту 11 недель (6-я неделя эксперимента). В условиях обычной диеты мыши FVB и C57 сохраняли сопоставимую массу жира на протяжении всего эксперимента (). Диета с высоким содержанием жиров вызвала значительное увеличение массы тела и жировой массы у обоих штаммов; однако изменения массы тела и жировой массы были более значительными у мышей C57. У мышей C57 было вдвое больше жира после 12 недель кормления с высоким содержанием жиров ().Общая разница в общей массе жира между мышами FVB и C57 коррелировала с пропорциональными различиями в количестве эпидидимального (внутрибрюшного), пахового (подкожного) и бурого жира (). Потребление калорий и активность были сопоставимы у мышей FVB HF и C57 HF; однако мыши FVB HF имели более высокий уровень покоя и общего потребления кислорода, а также более высокую ректальную температуру, что позволяет предположить, что увеличение расхода энергии, а не снижение потребления калорий, было причиной относительной устойчивости к ожирению, вызванному диетой с высоким содержанием жиров, у мышей FVB.Интересно, что в течение первых 2 недель кормления с высоким содержанием жиров мыши FVB и C57 показали сопоставимое увеличение общей жировой массы (). Мыши C57 HF продолжали быстро увеличивать жировую массу до 10 недели эксперимента, тогда как мыши FVB HF замедляли накопление жира около недели 3. У мышей C57 кормление с высоким содержанием жира вызывало постепенное увеличение как эпидидимальных, так и паховых жировых подушечек; Напротив, у мышей FVB жировая масса придатка яичка увеличилась лишь незначительно после 4 недель кормления с высоким содержанием жира, в то время как паховая жировая подушечка продолжала увеличиваться в размере на протяжении всего эксперимента (рис.S1). Кормление с высоким содержанием жиров вызывало значительное повышение уровня глюкозы и инсулина в крови как у мышей FVB, так и у мышей C57 (). Уровни инсулина и непереносимость глюкозы были выше у мышей C57 HF, чем у мышей FVB HF, что свидетельствует о более тяжелой инсулинорезистентности (). В соответствии с предыдущими сообщениями [23], [24], мыши C57 REG показали пониженные уровни триглицеридов в сыворотке по сравнению с мышами FVB REG без разницы в FFA (). Это происходило не из-за более высокой утилизации жира, так как коэффициент респираторного обмена () и скорость окисления жирных кислот, измеренная in vivo, () и в изолированных скелетных мышцах () были сопоставимы у мышей FVB REG и C57 REG.Более вероятно, что более низкие уровни триглицеридов в сыворотке у мышей C57 REG были вызваны гораздо более эффективным клиренсом циркулирующих триглицеридов, как предполагает тест на клиренс триглицеридов (). Кормление с высоким содержанием жиров снижало уровни циркулирующих триглицеридов как у мышей FVB, так и у мышей C57 и улучшало клиренс триглицеридов у последних штаммов (и). Оба штамма показали сопоставимое снижение коэффициента респираторного обмена, что свидетельствует о сопоставимом увеличении утилизации жирных кислот в условиях диеты с высоким содержанием жиров (). Взятые вместе, эти данные предполагают, что способность эффективно выводить триглицериды из кровотока может способствовать высокой способности накопления жира у мышей C57.
Изменения в составе тела мышей FVB и C57 в условиях стандартного корма (REG) и с высоким содержанием жира (HF).(A) Масса тела. (B) Жировая масса. Состав тела измеряли в указанные моменты времени у 8 мышей в группе с использованием МРТ-анализатора Echo 3-in-1. Высокожировое и контрольное кормление начали в возрасте 5 недель. Ось X показывает недели контролируемого кормления. Показан один из трех независимых экспериментов. Приведенные значения являются средними ± SEM.
Физиологические характеристики мышей FVB и C57.(A) Тест на толерантность к глюкозе был проведен на 10-й неделе контролируемого кормления с пищей и диетой с высоким содержанием жиров. Мышей не кормили в течение ночи и вводили глюкозу (2 мг / г, внутрибрюшинно). Каждая группа была представлена 8 мышами. (B) Окисление олеиновой кислоты в целом организме измеряли у самцов мышей FVB ad C57 в возрасте 10 недель в не голодном состоянии (8 мышей на группу), как описано в Gautam et al. [47]. (C) Окисление пальмитиновой кислоты измеряли в камбаловидной мышце, взятой у 10-недельных мышей FVB и C57, которых содержали на диете (8 мышей на группу).Образцы собирали в 8 часов утра у мышей, которых случайным образом кормили, или мышей, голодавших в течение 18 часов [методы, описанные в Toyoshima et al. [46]]. (D) Клиренс триглицеридов у самцов мышей FVB и C57 после 11 недель контролируемого кормления кормом и диетой с высоким содержанием жиров. Мышей не кормили в течение 4 часов, а затем вводили через желудочный зонд 400 мкл л арахисового масла. Плазму собирали ежечасно в течение 5 часов из хвостовой вены для измерения триглицеридов. Каждая группа была представлена 8 мышами.
Таблица 1
Характеристики мышей FVB и C57 после 12 недель высокожирового и контрольного кормления.
| FVB REG a | FVB HF a | C57 REG a | C57 HF | a 94 996| 912 912 912 912 912 Взаимодействие б | | ||||||||||
| Масса тела (г) | 29,1 ± 0,4 | 35,4 ± 1,4 | 29,1 ± 0,4 | 43,0 ± 1,8 | 0,003 | <0,001 | 0,003 | г)0.47 ± 0,03 | 1,23 ± 0,11 | 0,67 ± 0,08 | 2,00 ± 0,21 | <0,001 | <0,001 | 0,032 | |
| Паховой жир (г) | 0,3412 ± 0,02 | 0,190,3412 ± 0,02 | 0,3412 ± 0,02 | 0,45 ± 0,041,92 ± 0,27 | 0,003 | <0,001 | <0,003 | ||||||||
| Коричневый жир (г) | 0,14 ± 0,01 | 0,27 ± 0,04 | 0,1412 ± 0,01 | 0,01 0,09 0,003<0.001 | 0,003 | ||||||||||
| Потребление пищи (ккал / мышь / день) | 16,6 ± 1,0 | 18,0 ± 1,3 | 13,2 ± 0,8 | 17,7 ± 0,8 | 0,07 | 0,007 92| 3 ns Общее потребление кислорода (мл / г) | 9,5 ± 0,2 | 11,6 ± 0,3 | 9,0 ± 0,2 | 9,4 ± 0,2 | <0,001 | <0,001 | нс | | |
| Потребление кислорода в покое (мл / г) ) | 8.3 ± 0,2 | 9,7 ± 0,3 | 7,3 ± 0,2 | 8,1 ± 0,3 | <0,001 | <0,001 | 0,003 | ||||||||
| Коэффициент респираторного обмена | 0,86 ± 0,01 | 0,75 ± 0,01 0,01 | 0,72 ± 0,00 | нс | <0,001 | 0,006 | |||||||||
| Суммарная активность (торможение луча / мин) | 265 ± 5 | 190 ± 17 | 345 ± 41 | 122011293 нс | <0.001 | нс | |||||||||
| Ходьба (торможение луча / мин) | 105 ± 3 | 53 ± 10 | 144 ± 25 | 38 ± 5 | нс | <0,002 | 91 Ректальная температура (° C) | 36,0 ± 0,2 | 36,6 ± 0,2 | 35,4 ± 0,2 | 35,5 ± 0,2 | <0,001 | 0,057 | нс | |
| Триглицерид сыворотки (мг / дл) | ± 25 | 95 ± 24 | 89 ± 16 | 64 ± 6 | 0.002 | 0,004 | ns | ||||||||
| Свободные жирные кислоты (мМ) | 0,22 ± 0,03 | 0,22 ± 0,01 | 0,26 ± 0,04 | 0,27 ± 0,05 | 1292 ns | ns | ns Глюкоза крови (мг / дл) | 187 ± 16 | 225 ± 11 | 209 ± 11 | 247 ± 15 | нс | 0,009 | нс | |
| Инсулин сыворотки (мг / | 1,1) 912 ± 0,1 | 1,9 ± 0,3 | 2.2 ± 0,8 | 4,9 ± 1,2 | 0,011 | 0,033 | нс | ||||||||
| Клиренс триглицеридов (AUG c ) | 3238 ± 476 | 1651 ± 160 | 48 | 489 | <0,001 | 0,011 | <0,001 | ||||||||
| Толерантность к глюкозе (AUG d ) | 28633 ± 1372 | 39560 ± 3217 | 25947 ± 1212 | 001 | 0,003 |
Гипертрофия и гиперплазия с увеличением жировой массы
Чтобы проверить основной механизм различной скорости накопления жира в эпидидимальном жире мышей FVB и C57, мы измерили распределение массы и размеров клеток в образцах тканей эпидидимальный жир, собранный на 0, 2, 4 и 12 неделях контролируемого кормления (). Поскольку гистологический анализ не позволяет точно определить размер клеток адипоцитов, который будет обсуждаться позже, мы измерили распределение клеток по размеру с помощью счетчика Коултера и рассчитали средневзвешенный размер клеток и общее количество клеток эпидидимальной жировой подушки по этим измерениям, которые аналогичные значения в другом исследовании на мышах [5].Сильная корреляция наблюдалась между массой жировых подушечек и средневзвешенным размером клеток, а также между массой жировых подушечек и общим числом клеток, независимо от напряжения и разницы в диете (). Первая корреляция дала масштабное соотношение между массой жировой подушечки и средневзвешенным по объему размером клеток (). Кроме того, была обнаружена экспоненциальная зависимость между массой жировой подушечки, и общим числом клеток, где начальная масса жировой подушки была получена от контрольных мышей; и начальное количество клеток, и скорость увеличения количества клеток в массе жировой подушечки, были оценены по данным (;).Первоначальное количество клеток у мышей C57 было больше, чем исходное количество клеток у мышей FVB (фиг. S2C и S2D). По мере увеличения массы жировой ткани увеличивается общее количество клеток. Скорость увеличения числа клеток была выше при обычном питании, чем при диете с высоким содержанием жиров, и эта тенденция более очевидна у мышей C57 (;), что указывает на генетическое различие. Соотношение между результатами регулярной диеты и диеты с высоким содержанием жиров составляет 1,42 и 3,22 для мышей FVB и C57 соответственно (). Это может указывать на взаимодействие между генетикой и диетой при увеличении количества клеток.Обратите внимание, что мы также наблюдали аналогичные результаты с массой тела и массой жира, поскольку три величины (масса жировых подушечек придатка яичка, масса жира и масса тела) коррелируют друг с другом. Однако результаты с массой эпидидимальной жировой подушечки были наиболее подходящими: среднеквадратичное отклонение между данными и соответствием составило 9,73, 7,94, 3,56 × 10 5 и 5,54 × 10 6 , соответственно; результат с массой тела составил 11,82, 15,35, 3,65 × 10 5 и 4,82 × 10 6 ; результат с жировой массой 8.58, 8,61, 4,28 × 10 5 и 6,11 × 10 6 .
Нормализованное распределение клеток по размерам в жировой подушке придатка яичка.На графике нанесен один типичный пример контрольных мышей FVB в возрасте пяти недель. Кружки обозначают точки измеренных данных, а линия представляет собой аппроксимирующую кривую, которая представляет собой сумму одной гауссовой и двух экспоненциальных функций.
Таблица 2
Значения параметров модели роста жировой ткани.
Генетические и диетические эффекты на рост жировой ткани
Эти две сильные корреляции между массой жировой подушечки и гипертрофией и между массой жировой подушечки и гиперплазией позволяют предположить, что увеличение жировой ткани можно описать как систематический процесс роста в отношении для увеличения массы жировой прослойки.Мы расположили распределения клеток по размерам, отсортированные по массе эпидидимальных жировых подушечек (). Примечательно, что модель роста жировой ткани в формуле. (1) описывает эволюционирующую картину распределения клеток по размерам в зависимости от увеличения массы жировой прослойки. Модель количественно соответствовала экспериментальным распределениям размеров клеток, несмотря на то, что все распределения представляют собой данные поперечного сечения, полученные от отдельных животных. Различные значения параметров в модели, которые соответствуют каждому индивидуальному распределению размеров клеток для обоих штаммов и обоих режимов питания, дали количественные различия в процессе роста эпидидимальной жировой ткани между штаммами и между диетами ().Во-первых, максимальная зависящая от размера скорость роста и скорость колебаний размера клеток из-за обмена липидов продемонстрировали разницу, вызванную диетой, и меньшую разницу, вызванную деформацией. Зависимый от размера рост и колебания размера, приводящие к гипертрофии, по-видимому, регулируются в основном диетой. В частности, рост, зависящий от размера, сдвинул режим больших клеток в распределении размеров клеток в сторону больших размеров, а флуктуации липидного обмена увеличили разброс распределения вокруг режима больших клеток.Важно отметить, что результаты должны быть тщательно интерпретированы, потому что каждая скорость — это скорость на единицу увеличения массы жировой ткани, а не на единицу увеличения времени. Во-вторых, геометрические параметры (, и), которые определяют форму скорости роста, зависящей от размера, имели практически одинаковые значения независимо от диеты и разницы деформаций, за исключением. Следовательно, нижний критический размер, который дает размер, инициирующий рост клеток в зависимости от размера, и два параметра масштаба могут быть зафиксированы на с l = 37 µ m, η l = 12 µ м и η u = 63 µ м соответственно.С другой стороны, верхний критический размер, ограничивающий зависимый от размера клетки рост больших клеток, зависел от диеты; при кормлении с высоким содержанием жира этот пороговый размер сместился в сторону большего размера (). При диете с высоким содержанием жиров изменения параметров (,, и) увеличивают способность жировых тканей накапливать липиды за счет как гиперплазии, так и гипертрофии. Более низкие уровни триглицеридов в сыворотке при условии диеты с высоким содержанием жиров (и) могут коррелировать с увеличением накопления липидов в увеличенных жировых клетках, потому что не было обнаружено значительных различий в окислении жирных кислот, о чем свидетельствует отсутствие различий в соотношении респираторного обмена ().
Изменения распределения клеток по размерам с увеличением массы эпидидимальной жировой подушечки при кормлении и диетах с высоким содержанием жиров.Начальные распределения размеров клеток пятинедельных контрольных мышей приведены в первой строке каждого столбца пунктирными линиями. Точки показывают экспериментальные результаты. Обратите внимание, что результаты упорядочены по массе эпидидимальной жировой подушечки, а не по хронологическому времени (недели контролируемого кормления), указанному в скобках. Сплошные линии представляют собой нормализованные распределения клеток по размеру, соответствующие данной массе эпидидимальной жировой подушечки, предсказанной моделью роста жировой ткани.
Зависимая от размера скорость роста массы придатка яичка.Здесь для этого графика используются средние значения параметров. Символы показаны для облегчения сравнения строк.
Обсуждение
Наше центральное открытие состоит в том, что гиперплазия и гипертрофия жировых клеток в эпидидимальной жировой подушке является функцией массы жировой подушечки, даже несмотря на то, что у отдельных животных может потребоваться разное время для достижения заданной массы жировой подушечки. Следовательно, рост жировой ткани, представленный как изменения распределения клеток по размерам, можно систематически моделировать как процесс роста по отношению к увеличению массы жировой подушечки; это может отражать корреляцию между массой жировой подушечки и секрецией адипокинов и других сигнальных молекул, контролирующих рост жировой ткани.Соответственно, следует отметить, что нормы в нашей модели — это не обычные нормы увеличения на единицу времени, а нормы увеличения на единицу массы. Таким образом, несколько показателей (, и) в модели имели более высокие значения для животных, соблюдающих диету, чем для животных, соблюдающих диету с высоким содержанием жиров. Однако, если бы эти показатели были преобразованы в обычные нормы увеличения на единицу времени, они имели бы большие значения для диеты с высоким содержанием жиров, потому что требуется меньше времени для увеличения массы жировой подушечки на единицу из более крупных и более многочисленных клеток на диета с высоким содержанием жиров, чем для увеличения такой же величины за счет меньшего и меньшего количества клеток на диете (рис.S1). Было высказано предположение, что когда ожирение прогрессирует, рано возникает гипертрофия жировых клеток, а затем запускается гиперплазия [8]. Наше исследование показало, что рекрутирование новых клеток экспоненциально увеличивается с увеличением массы жировой ткани. Гипертрофия жировых клеток является основным фактором увеличения массы жировой прослойки, тогда как гиперплазия не способствует этому увеличению, потому что она возникает в небольших клетках, в которых хранится гораздо меньший объем жира. Таким образом, наша модель естественным образом воплощает идею о том, что на гиперплазию влияет гипертрофический рост клеток.С другой стороны, сообщалось, что гиперплазия жировых клеток возникает только на ранних стадиях развития; следовательно, не следует ожидать рекрутирования новых клеток на поздних стадиях даже в условиях ожирения [6], [7]. Возможно, возраст животных в нашем исследовании (6 недель) допускает возникновение гиперплазии.
Модель, разработанная здесь, может дать микроскопическое представление о росте жировых клеток, зависящем от размера, который не может быть рассмотрен статическими поперечными исследованиями.Например, мы обнаружили следующие специфические свойства роста клеток, зависящие от размера: нижний критический размер, инициирующий накопление липидов, не зависел от диеты у двух линий мышей, тогда как верхний критический размер ограничивал рост клеток до достижения необходимого уровня. необычайный размер, был увеличен на диете с высоким содержанием жиров. Эта зависимость роста клеток от размера является проверяемой гипотезой. Далее, параметр флуктуации размера клеток различался при обычном питании и рационе с высоким содержанием жиров; оно больше при диете с высоким содержанием жиров, когда оно преобразуется в единицы, соответствующие изменению единицы времени, а не единице изменения массы жировой подушечки.Таким образом, случайный процесс высвобождения и поглощения жира жировыми клетками происходит более активно при диете с высоким содержанием жиров, чем при обычной диете. Может быть интересно посмотреть, можно ли распространить эти результаты на другие штаммы и организмы.
По сравнению с исследованиями, в которых наблюдали единственный пик в распределении размеров клеток жировых клеток [11], [15] — [17], [28], мы наблюдали бимодальные распределения клеток по размерам, как сообщалось другими [12], [13], [29] — [32]. В большинстве исследований [12], [13], [31], [32] по наблюдению за бимодальностью использовалась технология счетчика Коултера, которая имеет несколько преимуществ для оценки всего распределения размеров ячеек [31]: Во-первых, можно доказать, что анализируемые ячейки быть подлинными жировыми клетками на основе морфологии и флотации; во-вторых, объем каждой клетки оценивается независимо от формы и без артефактов смещения центра, что является правилом, а не исключением, с использованием гистологических подходов; наконец, можно подсчитать и измерить достаточное количество частиц, чтобы получить статистически значимые комплексные кривые.Напротив, микроскопические методы гистологии могут не наблюдать мелкие клетки из-за влияния увеличения микроскопа [30], небольшого количества образцов и систематической ошибки выборки. Однако при использовании счетчика Коултера контаминация, не связанная с адипоцитами, может способствовать распределению клеток по размерам, особенно при малых размерах, хотя наш минимальный диаметр клеток, 22 µ м, превышает возможные диапазоны контаминации, от 10 до 20 µ м, упомянутый Mersmann и MacNeil [31]. Чтобы быть уверенным, мы снова проанализировали модифицированные данные, используя только распределения размеров клеток выше 35 µ м в диаметре с моделью, и пришли к тем же выводам (данные не показаны).Надир в распределении размеров клеток () может разделять две популяции клеток. ДеМартини и Франсендез определили маленькие клетки с диаметром менее 35 µ м как «очень маленькие жировые клетки» [29]. Согласно нашей модели, эти клетки имеют незначительный рост в зависимости от размера, поскольку их размер меньше нижнего критического размера, с l = 37 µ m. Следовательно, механизм роста, зависящий от размера, может естественным образом объяснить происхождение бимодальности в распределении жировых клеток по размеру.Клетки с размером только выше могут расти в зависимости от размера, но клетки с размером меньше могут расти случайным образом с флуктуацией размера за счет липидного обмена. Это разделение вызывает накопление клеток ниже размера, что дает более низкий пик в распределении размеров клеток. Эта популяция клеток может служить потенциальным резервуаром для зрелых жировых клеток. Процесс их созревания можно интерпретировать следующим образом: жировые клетки, достигающие критического размера, вследствие случайных колебаний размера могут расти с механизмом роста, зависящим от размера.Как упоминалось выше, изменение размера происходит более активно при диете с высоким содержанием жиров; следовательно, пласт может ускорить процесс созревания в условиях стимуляции.
В модель роста ткани мы включили привлечение новых клеток и рост существующих клеток, но не гибель старых клеток, потому что модель соответствовала данным без апоптоза жировых клеток. Этот результат также согласуется с исследованием, в котором наблюдалось, что жировая ткань придатка яичка мышей C57BL / 6 не демонстрирует значительной гибели клеток до 12 недель при диете с высоким содержанием жиров [33].Однако расширенная диета с высоким содержанием жиров в конечном итоге вызывает апоптоз жировых клеток [33]. Более того, в одном недавнем исследовании сообщается, что жировые клетки человека обновляются в течение десяти лет [7]. Таким образом, наша модель все еще нуждается в улучшении, чтобы ее можно было использовать в различных условиях. Следует учитывать гибель клеток и формально учитывать зависимость параметров модели от диеты. В этом исследовании мы сосредоточились на одном жировом депо, эпидидимальном жире, по нескольким причинам: 1) вес эпидидимальных жировых подушечек может быть определен более точно, чем вес паховых жировых подушечек из-за легкости иссечения; 2) морфология жировых клеток в эпидидимальном жире более однородна, чем в паховом жире, который содержит много клеток, подобных коричневой жировой ткани, особенно у мышей, устойчивых к ожирению, вызванному диетой; и 3) разница между генотипами была более очевидной в росте эпидидимального жира, который достигает плато через 4 недели у мышей FVB, но продолжает расти у мышей C57 на протяжении всего эксперимента, в отличие от пахового жира, который показывает устойчивый рост. у обоих штаммов (рис.S1). Хотя мы не измеряли распределение по размеру клеток других жировых отложений, мы измерили изменение массы паховых и бурых жировых отложений, что показывает аналогичную картину с эпидидимальным жировым депо (рис. S1). Таким образом, представляет интерес применить модель к другим жировым отложениям, которые имеют функциональные различия [34], [35], а также к другим видам, таким как человек, что остается для будущих исследований. Мы ожидаем, что модель может быть применена к таким разнообразным наборам данных, просто настроив параметры модели, потому что модель содержит общие механизмы роста тканей для привлечения новых клеток и их последующего развития.
Наши данные предполагают, что, по крайней мере, три фактора могут объяснить, почему мыши C57 набирают больше жира, чем мыши FVB, при диете с высоким содержанием жиров: во-первых, мыши FVB имеют повышенную скорость метаболизма и повышенную ректальную температуру, скорее всего, из-за повышенного симпатического тонуса. Хотя мы не обнаружили значительных различий в активности между штаммами в нашем исследовании, более подробные измерения поведения показали, что мыши FVB спонтанно гиперактивны по сравнению с мышами C57 [26]. У них также увеличивается частота сердечных сокращений [36], и они реагируют гипергликемией на различные виды лечения [37].Кроме того, они более чувствительны к стрессу, связанному с воздержанием и голоданием [38]. Все эти данные, взятые вместе, предполагают, что активность симпатической нервной системы увеличилась больше у мышей FVB, чем активность у мышей C57. Во-вторых, по сравнению с мышами FVB, мыши C57 очищают циркулирующие триглицериды более эффективно [23], [24], что, по крайней мере, частично можно объяснить более высокой активностью липазы в сыворотке крови [23] и более высокой способностью накапливать триглицериды в печени [23]. ], [24] и жировой ткани (показано в этом исследовании).Хотя молекулярные механизмы клиренса триглицеридов до конца не изучены, жировая ткань явно способствует выведению триглицеридов, поскольку способность выводить циркулирующие триглицериды нарушена у липоатрофных мышей [23]. В частности, сообщалось, что диета с высоким содержанием жиров увеличивает клиренс триглицеридов [39], что может быть связано с индукцией активности липопротеинлипазы в жировой ткани [40]. Наконец, по сравнению с мышами FVB, мыши C57 показали большее количество мелких жировых клеток, особенно при диете с высоким содержанием жиров.Было высказано предположение, что новые жировые клетки могут возникать из клеток-предшественников, которые находятся в депо белого жира взрослых [41], [42], и из других источников, таких как циркулирующие клетки-предшественники костного мозга [43]. Было показано, что рекрутирование обоих типов предшественников стимулируется диетой с высоким содержанием жиров [41], [43]. Возможно, что большее рекрутирование более мелких жировых клеток у мышей C57 может быть вызвано более высоким пулом клеток-предшественников или их более высокой внутренней способностью к дифференцировке адипоцитов.Однако in vitro мезенхимальных стволовых клеток , выделенных из наружного уха мышей FVB и C57, одинаково хорошо дифференцируются в жировые клетки [44]. Попытка сравнить дифференцировку стромальных клеток костного мозга мышей FVB и C57 в жировые клетки не увенчалась успехом из-за очень низкого выхода и плохой пролиферативной способности клеток, выделенных от мышей C57 [45]; однако костный мозг, по-видимому, не является основным источником новых жировых клеток, по крайней мере, у мышей [43]. Наша модель предполагает, что разница между генотипами в рекрутировании малых жировых клеток может быть автономной по отношению к жировым подушечкам, но молекулярный механизм, лежащий в основе этого различия, неясен.Жировая подушечка представляет собой сложный орган, содержащий множество различных типов клеток, включая зрелые жировые, предадипозные и сосудистые клетки, нервы, макрофаги и фибробласты. Количество предшественников адипоцитов и их пролиферация в ответ на внешние сигналы варьируются в зависимости от жировых отложений [35]. Необходимы дальнейшие исследования, чтобы определить, как специфичное для генотипа взаимодействие между разными типами клеток и секретируемыми факторами может влиять на скорость рекрутирования адипоцитов в определенные жировые депо.
Таким образом, мы получили математическую модель, описывающую рост жировой ткани с увеличением количества и размера клеток в зависимости от массы жировой подушечки придатка яичка.Основываясь на этой динамической модели, мы исследовали влияние генетики и диеты на рост жировой ткани. Сравнивая распределение размеров клеток двух штаммов и двух диет, мы пришли к выводу, что изменение размера клеток зависит от диеты, а изменение количества клеток зависит от генетики и диеты, а также от их взаимодействия.
Материалы и методы
Животные
Все процедуры были одобрены Комитетом по уходу и использованию животных Национального института диабета, болезней органов пищеварения и почек.Самцы мышей FVB и C57 были получены из лаборатории Джексона (Бар-Харбор, штат Мэн). Мышей выращивали по четыре в клетке с 12-часовым циклом свет / темнота (свет 06: 00–18: 00). В возрасте 5 недель мышей каждой линии разделили на 2 группы. Половине мышей давали обычную пищу NIH-07 (далее REG; Zeigler Brothers, Inc., Gardners, PA), содержащую 4,08 ккал / г (11% калорий из жира, 62% из углеводов и 26% из белка). Другая половина получала диету с высоким содержанием жиров, F3282 (далее HF; Bio-Serv, Frenchtown, NJ), содержащую 5.45 ккал / г (59% жиров, 26% углеводов и 15% белков). Вода и диета были предоставлены ad libitum . Было проведено пять независимых экспериментов, в каждой из которых использовалось 4 группы мышей: FVB REG, FVB HF, C57 REG и C57 HF. В трех экспериментах мышей содержали на контролируемой диете в течение 12 недель и использовали для анализа состава тела, физиологической характеристики и распределения клеток по размеру. Две дополнительные группы мышей были умерщвлены после 2 недель и 4 недель контрольного кормления с высоким содержанием жира только для определения распределения клеток по размеру.
Эксперименты in vivo
Состав тела, потребление пищи, скорость метаболизма, толерантность к глюкозе, клиренс триглицеридов и окисление жирных кислот в изолированной камбаловидной мышце измеряли, как описано ранее [46]. Окисление жирных кислот всего тела измеряли, как описано в Gautam et al. [47]. Кровь для биохимических анализов брали из хвостовой вены без голодания. Уровни глюкозы измеряли с помощью Glucometer Elite (Bayer, Elkhart, IN). Сывороточный инсулин анализировали с помощью радиоиммуноанализа (Linco Research, St.Чарльз, Миссури). Триглицериды сыворотки, холестерин (Thermo DMA, Луисвилл, Колорадо) и свободная жирная кислота (FFA) (Roche Applied Science, Индианаполис, Индиана) измеряли в соответствии с процедурами производителя.
Измерение размера клеток эпидидимального жира
Распределение размеров клеток эпидидимального жира измеряли через 2, 4 и 12 недель высокожирового и контрольного кормления с использованием Beckman Coulter Multisizer III, как описано ранее [9]. Вкратце, 20–30 мг жировой ткани были взяты из средней части путем рассечения и последующего удаления образца для фиксации из центра разрезанного эпидидимального жира.Образцы тканей немедленно фиксировали в четырехокиси осмия [48], инкубировали на водяной бане при 37 ° C в течение 48 ч, а затем определяли размер жировых клеток на приборе Beckman Coulter Multisizer III с апертурой 400 µ м. Диапазон размеров ячеек, которые можно эффективно измерить с помощью этой апертуры, составляет 20–240 µ м. Прибор был настроен на подсчет 6000 частиц, и суспензия фиксированных клеток была разбавлена так, чтобы совпадающий счет был <10%. После сбора размеров импульсов данные были выражены в виде диаметров частиц и отображены в виде гистограмм отсчетов по диаметру с использованием линейных интервалов и линейной шкалы для оси x ().Распределение клеток по размерам было измерено в четырех образцах из каждой группы, за исключением мышей C57 после 4-недельного воздействия диеты с высоким содержанием жиров, у которых было только три доступных образца. Образец был взят из каждой жировой подушечки и обработан отдельно. Затем каждый образец был посчитан как минимум дважды. Затем кривые для двух образцов усредняются, но только после проверки воспроизводимости между двумя образцами.
Модель роста жировой ткани
Распределение размеров клеток включает всю информацию, относящуюся к размерам клеток в ткани, и его изменения дают статистическое представление о детальном процессе роста каждой клетки.Чтобы изучить рост жировой ткани с точки зрения лежащих в основе микроскопических процессов, мы рассматриваем математическую модель, количественно определяющую процессы, которые изменяют распределение клеток по размерам. Модель может предсказать, сколько новых клеток образуется и как клетки разного размера растут по мере увеличения массы жировой ткани. Плотность количества клеток определенного размера (диаметра) при данной массе жировой подушечки — это конкретное количество, которое необходимо учитывать. Мы рассматриваем, как это распределение клеток по размерам изменяется с постепенным изменением массы жировой ткани.Эволюция распределения клеток по размерам с массой жировой ткани может быть смоделирована уравнением в частных производных:
(1)
Это уравнение включает три основных компонента процесса роста жировой ткани. Во-первых, мы предполагаем, что рекрутирование новых клеток происходит только при минимальном наблюдаемом размере клетки, который математически выражается как дельта-функция. Скорость набора по отношению к массе жировой подушечки определяется экспоненциальной функцией:
(2)
, где — исходное общее количество клеток при данной начальной массе жировой подушечки, а — скорость увеличения количества клеток на единицу изменения. в массе жировой подушечки.Изменение общего числа клеток — это скорость пополнения новых клеток, если гибель клеток незначительна; мы не обнаружили необходимости включать апоптоз в клетки любого размера, чтобы соответствовать этим экспериментальным данным. Следовательно, этот коэффициент пополнения может быть непосредственно получен из экспериментального результата, используя соотношение между общим числом клеток и массой жировой подушечки путем дифференцирования функции по отношению к. Во-вторых, рост клеток зависит от размера клетки. После созревания жировых клеток до определенного размера они могут накапливать жир, вызывая гипертрофию.В дополнение к этой ограничивающей скорости роста маленьких жировых клеток, может также существовать верхний предел роста, потому что большие жировые клетки не могут расти бесконечно в результате этого процесса роста, хотя они могут достигать больших размеров из-за колебаний размера, вызванных обменом липидов. Повышение и снижение скорости роста клеток в зависимости от размера клетки можно описать с помощью общей функциональной формы, умножающей две сигмоидальные функции,
(3)
, где представляет максимальную скорость роста; и — соответственно нижний и верхний критические размеры, которые дают половину максимальной скорости роста; и укажите их масштаб ().Наконец, последний член в формуле. (1) представляет колебания размера клеток с постоянной скоростью, которые отражают обмен липидов, случайным образом происходящий в жировых клетках. Этот липидный оборот является единственным механизмом роста для крупных клеток, размер которых превышает верхний критический размер. Как правило, рост клеток, зависящий от размера, приводит к увеличению их распределения по размерам, в то время как колебания размеров распространяют распределение по размерам.
Метод оптимизации
Чтобы оптимизировать параметры модели, чтобы они могли точно описывать эволюцию распределения размеров ячеек в эксперименте, мы использовали минимизацию «функции стоимости», которая количественно определяет отклонение между моделью и результатами эксперимента.Чтобы определить функцию стоимости, нормализованное распределение размеров клеток при заданной массе жира сравнивалось с данными моделирования с набором параметров:
(6)
, где — общее количество интервалов размера клеток, а — общее количество заданных масса жира. Масштаб функции стоимости был рассчитан на основе внутренней флуктуации экспериментальных данных, которую можно определить как квадрат отклонения между измеренным распределением ячеек по размерам и функцией плавной аппроксимации:
(7)
Эта внутренняя флуктуация численно составляет около 10 процентов квадрата отклонения между экспериментальными и модельными данными.В качестве функции аппроксимации мы использовали сумму двух экспонент и гауссову форму,
(8)
, форму, которая использовалась для аппроксимации распределений размеров жировых клеток [9]. Подбор этих параметров был выполнен с использованием процедуры подбора нелинейной кривой в MATLAB R2007a (Натик, Массачусетс, США). Для процесса оптимизации мы специально использовали метод Монте-Карло с параллельным темперированием, чтобы найти глобальный минимум функции стоимости [50]. Мы использовали 10 равномерно распределенных значений (от 0,1 до 1) для параметра темперирования и запускали десять цепочек параллельно с вероятностью обновления.На каждых 20 шагах случайным образом выбиралась пара смежных имитаций по десяти параметрам отпуска, и их состояния параметров менялись с вероятностью. После достижения равновесия было использовано 20 000 итераций с фиксированным параметром отпуска для оценки оптимальных значений параметров и их стандартных ошибок.
Мы также использовали этот метод для оценки исходного общего количества клеток, и скорости его увеличения, исходя из соотношения между жировой массой и общим количеством клеток (). При минимизации функции аппроксимации и экспериментальных данных мы использовали ограничение, согласно которому начальное количество клеток одинаково независимо от условий диеты, т.е.е., обычные и жирные диеты. Средняя жировая масса четырех контрольных мышей перед регулярной диетой и диетой с высоким содержанием жиров была использована в качестве начальной жировой массы, которая составляет 0,34 г и 0,29 г для мышей FVB и C57, соответственно. Мы оценили погрешности и, распространив 10-процентные статистические флуктуации, наблюдаемые в экспериментальных данных.
Численное решение дифференциального уравнения с частными производными
Мы решили следующую дискретную версию нашей модели, заданную как непрерывное дифференциальное уравнение с частными производными в формуле.(1):
(9)
с интервалом масс δm = 0,1 мг и интервалом размеров δs = 0,73 µ м.
Динамика роста жировой ткани
Abstract
Рост жировой ткани происходит по двум механизмам: гиперплазия (увеличение количества клеток) и гипертрофия (увеличение размера клеток). Генетика и диета влияют на относительный вклад этих двух механизмов в рост жировой ткани при ожирении. В этом исследовании распределение по размеру эпидидимальных жировых клеток от двух линий мышей, устойчивых к ожирению FVB / N и склонных к ожирению C57BL / 6, измеряли через 2, 4 и 12 недель в условиях регулярного кормления и питания с высоким содержанием жира.Общее количество клеток в жировой подушке придатка яичка оценивали по массе жировой подушечки и нормализованному распределению клеток по размеру. Число клеток и средневзвешенный размер клеток увеличиваются в зависимости от массы жировой прослойки. Чтобы точно решить проблему роста жировой ткани, мы разработали математическую модель, описывающую эволюцию распределения размеров жировых клеток как функцию увеличения массы жировой ткани, а не увеличения хронологического времени. Наша модель описывает привлечение новых жировых клеток и их последующее развитие в разных штаммах и с разными режимами питания, с общими механизмами, но с параметрами модели, зависящими от диеты и генетики.По сравнению со штаммом FVB / N, штамм C57BL / 6 имеет большее количество мелких жировых клеток. Гиперплазия усиливается за счет диеты с высоким содержанием жиров в зависимости от штамма, что предполагает синергетическое взаимодействие между генетикой и диетой. Кроме того, кормление с высоким содержанием жиров увеличивает скорость роста жировых клеток, независимо от напряжения, что отражает увеличение количества калорий, требующих хранения. Кроме того, диета с высоким содержанием жиров приводит к резкому увеличению распределения жировых клеток по размерам у обоих штаммов; это означает увеличение колебаний размера жировых клеток за счет липидного обмена.
Авторское резюме
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. Динамическое изменение жировой ткани in vivo невозможно отслеживать в режиме реального времени из-за текущих технических ограничений. Однако мы можем измерить распределение жировых клеток по размеру у отдельных животных. Наша основная цель — извлечь динамические характеристики ремоделирования тканей из снимков распределения клеток по размерам.Мы разрабатываем математическую модель, которая интерполирует измерения распределения клеток по размеру и предсказывает непрерывное изменение распределения клеток по размеру в зависимости от увеличения массы жировых подушечек. Наша модель роста жировой ткани включает три основных компонента: рекрутирование новых клеток, рост клеток в зависимости от размера и колебания размера клеток. В частности, мы сравнили рост жировой ткани у склонных к ожирению и устойчивых к ожирению мышей при стандартной диете или диете с высоким содержанием жиров, чтобы изучить генетическое и диетическое влияние на рост жировой ткани.Применяя нашу модель к этим различным условиям, мы обнаружили, что увеличение размера жировых клеток зависит от диеты. С другой стороны, увеличение количества жировых клеток, вызванное диетой, зависит от деформации, что предполагает синергию между генетикой и диетой.
Введение
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. На фенотип ожирения жировой ткани влияют диета и генетика, а также их взаимодействие [1] — [4].Начиная с исследований Джонсона и Хирша [5], существует обширная литература о росте жировой ткани при нормальном и патологическом развитии, характеризующая состояние ткани с точки зрения среднего размера и количества клеток. Гиперпластический рост появляется только на ранних стадиях развития жировой ткани [6], [7]. Гипертрофия возникает до гиперплазии, чтобы удовлетворить потребность в дополнительных накоплениях жира при прогрессировании ожирения [8]. Однако оказалось трудно понять, как диета и генетика конкретно влияют на гиперплазию и / или гипертрофию жировых клеток из-за ограниченных данных о росте жировой ткани.
Помимо этих исследований, недавно стало возможным точно измерить распределение клеток по размерам. Эта подробная информация, по сравнению со средним размером клеток и общим числом клеток, может использоваться для вычисления многих величин, связанных с размером, которые позволяют более точно охарактеризовать процесс роста жировой ткани. Кумулянты распределения клеток по размерам могут использоваться для вычисления других физиологических величин, таких как средневзвешенный размер клеток. Распределение клеток по размерам можно использовать для оценки общего количества клеток в жировой подушке по ее массе.Более того, считается, что некоторые специфические метаболические свойства, например, резистентность к инсулину [9] и секреция адипокина [10], зависят от точного распределения клеток по размерам, а не от среднего размера клеток. Действительно, в нескольких исследованиях изучается изменение распределения жировых клеток по размерам в различных условиях при развитии куриного эмбриона [11], у худых и страдающих ожирением крыс Zucker [12], [13], крыс линии Wistar с частичной липэктомией [14], биопсии кролика [ 15] и жировой ткани человека [16], [17]. Эти исследования были сосредоточены только на статических различиях между распределениями клеток по размерам в разных условиях.Однако статические распределения размеров клеток в поперечном сечении для ряда снимков развития животных можно использовать для вывода динамики роста жировой ткани, если мы сможем надлежащим образом проанализировать снимки с помощью математического моделирования. Учитывая нынешние технические ограничения, это может быть лучший доступный подход к микроскопическому и продольному пониманию роста жировой ткани in vivo , хотя в недавнем эксперименте были получены микроскопические наблюдения накопления липидов в липидных каплях жировых клеток [18].
Для изучения генетического и диетического влияния на динамический процесс роста жировой ткани мы получили распределения по размерам клеток эпидидимального жира у устойчивых к ожирению линий мышей FVB / N (далее FVB) и склонных к ожирению линий мышей C57BL / 6 (C57) в соответствии с стандартные диеты и диеты с высоким содержанием жиров. Мышь C57 является наиболее охарактеризованной моделью ожирения, вызванного диетой [19], а мышь FVB является предпочтительной моделью для создания трансгенных мышей [20]. Эти два широко используемых инбредных штамма мышей генетически довольно далеки [21], [22] и имеют различные метаболические фенотипы: по сравнению с мышами FVB, мыши C57 имеют низкие уровни циркулирующих триглицеридов [21] и повышенный клиренс триглицеридов [23], [24]; Мыши FVB характеризуются относительно более высокой печеночной инсулинорезистентностью, контррегуляторной реакцией на гипогликемию и сниженной глюкозо-стимулированной секрецией инсулина [25]; Также известно, что мыши FVB являются спонтанно гиперактивными [26] и относительно худыми, поскольку они, по-видимому, менее восприимчивы к диете с высоким содержанием жиров, чем мыши C57 [27].Однако развитие ожирения, вызванного диетой, у этих двух штаммов формально не сравнивалось. В этом исследовании мы разработали математическую модель, прогнозирующую изменение распределения клеток по размерам в зависимости от массы жировой подушечки придатка яичка, для количественного анализа динамических характеристик, зависящих от генетики и / или диеты. Модель роста жировой ткани описывает, сколько новых клеток образуется, как каждая клетка растет в зависимости от ее размера и как липидный обмен приводит к колебаниям размера, которые вызывают расширение распределения клеток по размерам.По мере увеличения массы эпидидимальных жировых подушек распределение клеток по размерам систематически изменяется в зависимости как от генетики, так и от диеты. Сравнивая экспериментальные результаты с теоретической моделью роста, мы обнаружили, что гипертрофия сильно коррелирует с диетой. С другой стороны, гиперплазия зависит от генетики. Изменения при гиперплазии, вызванные диетой, зависят от штамма, что предполагает взаимодействие между диетой и генетикой.
Результаты
Влияние диеты с высоким содержанием жиров на состав тела мышей FVB и C57
В начале эксперимента (в возрасте 5 недель) мыши C57 были значительно легче, чем мыши FVB () из-за разницы в худой масса, хотя общая жировая масса не отличалась ().Когда мышей поддерживали на обычном питании, разница в массе тела исчезла к возрасту 11 недель (6-я неделя эксперимента). В условиях обычной диеты мыши FVB и C57 сохраняли сопоставимую массу жира на протяжении всего эксперимента (). Диета с высоким содержанием жиров вызвала значительное увеличение массы тела и жировой массы у обоих штаммов; однако изменения массы тела и жировой массы были более значительными у мышей C57. У мышей C57 было вдвое больше жира после 12 недель кормления с высоким содержанием жиров ().Общая разница в общей массе жира между мышами FVB и C57 коррелировала с пропорциональными различиями в количестве эпидидимального (внутрибрюшного), пахового (подкожного) и бурого жира (). Потребление калорий и активность были сопоставимы у мышей FVB HF и C57 HF; однако мыши FVB HF имели более высокий уровень покоя и общего потребления кислорода, а также более высокую ректальную температуру, что позволяет предположить, что увеличение расхода энергии, а не снижение потребления калорий, было причиной относительной устойчивости к ожирению, вызванному диетой с высоким содержанием жиров, у мышей FVB.Интересно, что в течение первых 2 недель кормления с высоким содержанием жиров мыши FVB и C57 показали сопоставимое увеличение общей жировой массы (). Мыши C57 HF продолжали быстро увеличивать жировую массу до 10 недели эксперимента, тогда как мыши FVB HF замедляли накопление жира около недели 3. У мышей C57 кормление с высоким содержанием жира вызывало постепенное увеличение как эпидидимальных, так и паховых жировых подушечек; Напротив, у мышей FVB жировая масса придатка яичка увеличилась лишь незначительно после 4 недель кормления с высоким содержанием жира, в то время как паховая жировая подушечка продолжала увеличиваться в размере на протяжении всего эксперимента (рис.S1). Кормление с высоким содержанием жиров вызывало значительное повышение уровня глюкозы и инсулина в крови как у мышей FVB, так и у мышей C57 (). Уровни инсулина и непереносимость глюкозы были выше у мышей C57 HF, чем у мышей FVB HF, что свидетельствует о более тяжелой инсулинорезистентности (). В соответствии с предыдущими сообщениями [23], [24], мыши C57 REG показали пониженные уровни триглицеридов в сыворотке по сравнению с мышами FVB REG без разницы в FFA (). Это происходило не из-за более высокой утилизации жира, так как коэффициент респираторного обмена () и скорость окисления жирных кислот, измеренная in vivo, () и в изолированных скелетных мышцах () были сопоставимы у мышей FVB REG и C57 REG.Более вероятно, что более низкие уровни триглицеридов в сыворотке у мышей C57 REG были вызваны гораздо более эффективным клиренсом циркулирующих триглицеридов, как предполагает тест на клиренс триглицеридов (). Кормление с высоким содержанием жиров снижало уровни циркулирующих триглицеридов как у мышей FVB, так и у мышей C57 и улучшало клиренс триглицеридов у последних штаммов (и). Оба штамма показали сопоставимое снижение коэффициента респираторного обмена, что свидетельствует о сопоставимом увеличении утилизации жирных кислот в условиях диеты с высоким содержанием жиров (). Взятые вместе, эти данные предполагают, что способность эффективно выводить триглицериды из кровотока может способствовать высокой способности накопления жира у мышей C57.
Изменения в составе тела мышей FVB и C57 в условиях стандартного корма (REG) и с высоким содержанием жира (HF).(A) Масса тела. (B) Жировая масса. Состав тела измеряли в указанные моменты времени у 8 мышей в группе с использованием МРТ-анализатора Echo 3-in-1. Высокожировое и контрольное кормление начали в возрасте 5 недель. Ось X показывает недели контролируемого кормления. Показан один из трех независимых экспериментов. Приведенные значения являются средними ± SEM.
Физиологические характеристики мышей FVB и C57.(A) Тест на толерантность к глюкозе был проведен на 10-й неделе контролируемого кормления с пищей и диетой с высоким содержанием жиров. Мышей не кормили в течение ночи и вводили глюкозу (2 мг / г, внутрибрюшинно). Каждая группа была представлена 8 мышами. (B) Окисление олеиновой кислоты в целом организме измеряли у самцов мышей FVB ad C57 в возрасте 10 недель в не голодном состоянии (8 мышей на группу), как описано в Gautam et al. [47]. (C) Окисление пальмитиновой кислоты измеряли в камбаловидной мышце, взятой у 10-недельных мышей FVB и C57, которых содержали на диете (8 мышей на группу).Образцы собирали в 8 часов утра у мышей, которых случайным образом кормили, или мышей, голодавших в течение 18 часов [методы, описанные в Toyoshima et al. [46]]. (D) Клиренс триглицеридов у самцов мышей FVB и C57 после 11 недель контролируемого кормления кормом и диетой с высоким содержанием жиров. Мышей не кормили в течение 4 часов, а затем вводили через желудочный зонд 400 мкл л арахисового масла. Плазму собирали ежечасно в течение 5 часов из хвостовой вены для измерения триглицеридов. Каждая группа была представлена 8 мышами.
Таблица 1
Характеристики мышей FVB и C57 после 12 недель высокожирового и контрольного кормления.
| FVB REG a | FVB HF a | C57 REG a | C57 HF | a 94 996| 912 912 912 912 912 Взаимодействие б | | ||||||||||
| Масса тела (г) | 29,1 ± 0,4 | 35,4 ± 1,4 | 29,1 ± 0,4 | 43,0 ± 1,8 | 0,003 | <0,001 | 0,003 | г)0.47 ± 0,03 | 1,23 ± 0,11 | 0,67 ± 0,08 | 2,00 ± 0,21 | <0,001 | <0,001 | 0,032 | |
| Паховой жир (г) | 0,3412 ± 0,02 | 0,190,3412 ± 0,02 | 0,3412 ± 0,02 | 0,45 ± 0,041,92 ± 0,27 | 0,003 | <0,001 | <0,003 | ||||||||
| Коричневый жир (г) | 0,14 ± 0,01 | 0,27 ± 0,04 | 0,1412 ± 0,01 | 0,01 0,09 0,003<0.001 | 0,003 | ||||||||||
| Потребление пищи (ккал / мышь / день) | 16,6 ± 1,0 | 18,0 ± 1,3 | 13,2 ± 0,8 | 17,7 ± 0,8 | 0,07 | 0,007 92| 3 ns Общее потребление кислорода (мл / г) | 9,5 ± 0,2 | 11,6 ± 0,3 | 9,0 ± 0,2 | 9,4 ± 0,2 | <0,001 | <0,001 | нс | | |
| Потребление кислорода в покое (мл / г) ) | 8.3 ± 0,2 | 9,7 ± 0,3 | 7,3 ± 0,2 | 8,1 ± 0,3 | <0,001 | <0,001 | 0,003 | ||||||||
| Коэффициент респираторного обмена | 0,86 ± 0,01 | 0,75 ± 0,01 0,01 | 0,72 ± 0,00 | нс | <0,001 | 0,006 | |||||||||
| Суммарная активность (торможение луча / мин) | 265 ± 5 | 190 ± 17 | 345 ± 41 | 122011293 нс | <0.001 | нс | |||||||||
| Ходьба (торможение луча / мин) | 105 ± 3 | 53 ± 10 | 144 ± 25 | 38 ± 5 | нс | <0,002 | 91 Ректальная температура (° C) | 36,0 ± 0,2 | 36,6 ± 0,2 | 35,4 ± 0,2 | 35,5 ± 0,2 | <0,001 | 0,057 | нс | |
| Триглицерид сыворотки (мг / дл) | ± 25 | 95 ± 24 | 89 ± 16 | 64 ± 6 | 0.002 | 0,004 | ns | ||||||||
| Свободные жирные кислоты (мМ) | 0,22 ± 0,03 | 0,22 ± 0,01 | 0,26 ± 0,04 | 0,27 ± 0,05 | 1292 ns | ns | ns Глюкоза крови (мг / дл) | 187 ± 16 | 225 ± 11 | 209 ± 11 | 247 ± 15 | нс | 0,009 | нс | |
| Инсулин сыворотки (мг / | 1,1) 912 ± 0,1 | 1,9 ± 0,3 | 2.2 ± 0,8 | 4,9 ± 1,2 | 0,011 | 0,033 | нс | ||||||||
| Клиренс триглицеридов (AUG c ) | 3238 ± 476 | 1651 ± 160 | 48 | 489 | <0,001 | 0,011 | <0,001 | ||||||||
| Толерантность к глюкозе (AUG d ) | 28633 ± 1372 | 39560 ± 3217 | 25947 ± 1212 | 001 | 0,003 |
Гипертрофия и гиперплазия с увеличением жировой массы
Чтобы проверить основной механизм различной скорости накопления жира в эпидидимальном жире мышей FVB и C57, мы измерили распределение массы и размеров клеток в образцах тканей эпидидимальный жир, собранный на 0, 2, 4 и 12 неделях контролируемого кормления (). Поскольку гистологический анализ не позволяет точно определить размер клеток адипоцитов, который будет обсуждаться позже, мы измерили распределение клеток по размеру с помощью счетчика Коултера и рассчитали средневзвешенный размер клеток и общее количество клеток эпидидимальной жировой подушки по этим измерениям, которые аналогичные значения в другом исследовании на мышах [5].Сильная корреляция наблюдалась между массой жировых подушечек и средневзвешенным размером клеток, а также между массой жировых подушечек и общим числом клеток, независимо от напряжения и разницы в диете (). Первая корреляция дала масштабное соотношение между массой жировой подушечки и средневзвешенным по объему размером клеток (). Кроме того, была обнаружена экспоненциальная зависимость между массой жировой подушечки, и общим числом клеток, где начальная масса жировой подушки была получена от контрольных мышей; и начальное количество клеток, и скорость увеличения количества клеток в массе жировой подушечки, были оценены по данным (;).Первоначальное количество клеток у мышей C57 было больше, чем исходное количество клеток у мышей FVB (фиг. S2C и S2D). По мере увеличения массы жировой ткани увеличивается общее количество клеток. Скорость увеличения числа клеток была выше при обычном питании, чем при диете с высоким содержанием жиров, и эта тенденция более очевидна у мышей C57 (;), что указывает на генетическое различие. Соотношение между результатами регулярной диеты и диеты с высоким содержанием жиров составляет 1,42 и 3,22 для мышей FVB и C57 соответственно (). Это может указывать на взаимодействие между генетикой и диетой при увеличении количества клеток.Обратите внимание, что мы также наблюдали аналогичные результаты с массой тела и массой жира, поскольку три величины (масса жировых подушечек придатка яичка, масса жира и масса тела) коррелируют друг с другом. Однако результаты с массой эпидидимальной жировой подушечки были наиболее подходящими: среднеквадратичное отклонение между данными и соответствием составило 9,73, 7,94, 3,56 × 10 5 и 5,54 × 10 6 , соответственно; результат с массой тела составил 11,82, 15,35, 3,65 × 10 5 и 4,82 × 10 6 ; результат с жировой массой 8.58, 8,61, 4,28 × 10 5 и 6,11 × 10 6 .
Нормализованное распределение клеток по размерам в жировой подушке придатка яичка.На графике нанесен один типичный пример контрольных мышей FVB в возрасте пяти недель. Кружки обозначают точки измеренных данных, а линия представляет собой аппроксимирующую кривую, которая представляет собой сумму одной гауссовой и двух экспоненциальных функций.
Таблица 2
Значения параметров модели роста жировой ткани.
Генетические и диетические эффекты на рост жировой ткани
Эти две сильные корреляции между массой жировой подушечки и гипертрофией и между массой жировой подушечки и гиперплазией позволяют предположить, что увеличение жировой ткани можно описать как систематический процесс роста в отношении для увеличения массы жировой прослойки.Мы расположили распределения клеток по размерам, отсортированные по массе эпидидимальных жировых подушечек (). Примечательно, что модель роста жировой ткани в формуле. (1) описывает эволюционирующую картину распределения клеток по размерам в зависимости от увеличения массы жировой прослойки. Модель количественно соответствовала экспериментальным распределениям размеров клеток, несмотря на то, что все распределения представляют собой данные поперечного сечения, полученные от отдельных животных. Различные значения параметров в модели, которые соответствуют каждому индивидуальному распределению размеров клеток для обоих штаммов и обоих режимов питания, дали количественные различия в процессе роста эпидидимальной жировой ткани между штаммами и между диетами ().Во-первых, максимальная зависящая от размера скорость роста и скорость колебаний размера клеток из-за обмена липидов продемонстрировали разницу, вызванную диетой, и меньшую разницу, вызванную деформацией. Зависимый от размера рост и колебания размера, приводящие к гипертрофии, по-видимому, регулируются в основном диетой. В частности, рост, зависящий от размера, сдвинул режим больших клеток в распределении размеров клеток в сторону больших размеров, а флуктуации липидного обмена увеличили разброс распределения вокруг режима больших клеток.Важно отметить, что результаты должны быть тщательно интерпретированы, потому что каждая скорость — это скорость на единицу увеличения массы жировой ткани, а не на единицу увеличения времени. Во-вторых, геометрические параметры (, и), которые определяют форму скорости роста, зависящей от размера, имели практически одинаковые значения независимо от диеты и разницы деформаций, за исключением. Следовательно, нижний критический размер, который дает размер, инициирующий рост клеток в зависимости от размера, и два параметра масштаба могут быть зафиксированы на с l = 37 µ m, η l = 12 µ м и η u = 63 µ м соответственно.С другой стороны, верхний критический размер, ограничивающий зависимый от размера клетки рост больших клеток, зависел от диеты; при кормлении с высоким содержанием жира этот пороговый размер сместился в сторону большего размера (). При диете с высоким содержанием жиров изменения параметров (,, и) увеличивают способность жировых тканей накапливать липиды за счет как гиперплазии, так и гипертрофии. Более низкие уровни триглицеридов в сыворотке при условии диеты с высоким содержанием жиров (и) могут коррелировать с увеличением накопления липидов в увеличенных жировых клетках, потому что не было обнаружено значительных различий в окислении жирных кислот, о чем свидетельствует отсутствие различий в соотношении респираторного обмена ().
Изменения распределения клеток по размерам с увеличением массы эпидидимальной жировой подушечки при кормлении и диетах с высоким содержанием жиров.Начальные распределения размеров клеток пятинедельных контрольных мышей приведены в первой строке каждого столбца пунктирными линиями. Точки показывают экспериментальные результаты. Обратите внимание, что результаты упорядочены по массе эпидидимальной жировой подушечки, а не по хронологическому времени (недели контролируемого кормления), указанному в скобках. Сплошные линии представляют собой нормализованные распределения клеток по размеру, соответствующие данной массе эпидидимальной жировой подушечки, предсказанной моделью роста жировой ткани.
Зависимая от размера скорость роста массы придатка яичка.Здесь для этого графика используются средние значения параметров. Символы показаны для облегчения сравнения строк.
Обсуждение
Наше центральное открытие состоит в том, что гиперплазия и гипертрофия жировых клеток в эпидидимальной жировой подушке является функцией массы жировой подушечки, даже несмотря на то, что у отдельных животных может потребоваться разное время для достижения заданной массы жировой подушечки. Следовательно, рост жировой ткани, представленный как изменения распределения клеток по размерам, можно систематически моделировать как процесс роста по отношению к увеличению массы жировой подушечки; это может отражать корреляцию между массой жировой подушечки и секрецией адипокинов и других сигнальных молекул, контролирующих рост жировой ткани.Соответственно, следует отметить, что нормы в нашей модели — это не обычные нормы увеличения на единицу времени, а нормы увеличения на единицу массы. Таким образом, несколько показателей (, и) в модели имели более высокие значения для животных, соблюдающих диету, чем для животных, соблюдающих диету с высоким содержанием жиров. Однако, если бы эти показатели были преобразованы в обычные нормы увеличения на единицу времени, они имели бы большие значения для диеты с высоким содержанием жиров, потому что требуется меньше времени для увеличения массы жировой подушечки на единицу из более крупных и более многочисленных клеток на диета с высоким содержанием жиров, чем для увеличения такой же величины за счет меньшего и меньшего количества клеток на диете (рис.S1). Было высказано предположение, что когда ожирение прогрессирует, рано возникает гипертрофия жировых клеток, а затем запускается гиперплазия [8]. Наше исследование показало, что рекрутирование новых клеток экспоненциально увеличивается с увеличением массы жировой ткани. Гипертрофия жировых клеток является основным фактором увеличения массы жировой прослойки, тогда как гиперплазия не способствует этому увеличению, потому что она возникает в небольших клетках, в которых хранится гораздо меньший объем жира. Таким образом, наша модель естественным образом воплощает идею о том, что на гиперплазию влияет гипертрофический рост клеток.С другой стороны, сообщалось, что гиперплазия жировых клеток возникает только на ранних стадиях развития; следовательно, не следует ожидать рекрутирования новых клеток на поздних стадиях даже в условиях ожирения [6], [7]. Возможно, возраст животных в нашем исследовании (6 недель) допускает возникновение гиперплазии.
Модель, разработанная здесь, может дать микроскопическое представление о росте жировых клеток, зависящем от размера, который не может быть рассмотрен статическими поперечными исследованиями.Например, мы обнаружили следующие специфические свойства роста клеток, зависящие от размера: нижний критический размер, инициирующий накопление липидов, не зависел от диеты у двух линий мышей, тогда как верхний критический размер ограничивал рост клеток до достижения необходимого уровня. необычайный размер, был увеличен на диете с высоким содержанием жиров. Эта зависимость роста клеток от размера является проверяемой гипотезой. Далее, параметр флуктуации размера клеток различался при обычном питании и рационе с высоким содержанием жиров; оно больше при диете с высоким содержанием жиров, когда оно преобразуется в единицы, соответствующие изменению единицы времени, а не единице изменения массы жировой подушечки.Таким образом, случайный процесс высвобождения и поглощения жира жировыми клетками происходит более активно при диете с высоким содержанием жиров, чем при обычной диете. Может быть интересно посмотреть, можно ли распространить эти результаты на другие штаммы и организмы.
По сравнению с исследованиями, в которых наблюдали единственный пик в распределении размеров клеток жировых клеток [11], [15] — [17], [28], мы наблюдали бимодальные распределения клеток по размерам, как сообщалось другими [12], [13], [29] — [32]. В большинстве исследований [12], [13], [31], [32] по наблюдению за бимодальностью использовалась технология счетчика Коултера, которая имеет несколько преимуществ для оценки всего распределения размеров ячеек [31]: Во-первых, можно доказать, что анализируемые ячейки быть подлинными жировыми клетками на основе морфологии и флотации; во-вторых, объем каждой клетки оценивается независимо от формы и без артефактов смещения центра, что является правилом, а не исключением, с использованием гистологических подходов; наконец, можно подсчитать и измерить достаточное количество частиц, чтобы получить статистически значимые комплексные кривые.Напротив, микроскопические методы гистологии могут не наблюдать мелкие клетки из-за влияния увеличения микроскопа [30], небольшого количества образцов и систематической ошибки выборки. Однако при использовании счетчика Коултера контаминация, не связанная с адипоцитами, может способствовать распределению клеток по размерам, особенно при малых размерах, хотя наш минимальный диаметр клеток, 22 µ м, превышает возможные диапазоны контаминации, от 10 до 20 µ м, упомянутый Mersmann и MacNeil [31]. Чтобы быть уверенным, мы снова проанализировали модифицированные данные, используя только распределения размеров клеток выше 35 µ м в диаметре с моделью, и пришли к тем же выводам (данные не показаны).Надир в распределении размеров клеток () может разделять две популяции клеток. ДеМартини и Франсендез определили маленькие клетки с диаметром менее 35 µ м как «очень маленькие жировые клетки» [29]. Согласно нашей модели, эти клетки имеют незначительный рост в зависимости от размера, поскольку их размер меньше нижнего критического размера, с l = 37 µ m. Следовательно, механизм роста, зависящий от размера, может естественным образом объяснить происхождение бимодальности в распределении жировых клеток по размеру.Клетки с размером только выше могут расти в зависимости от размера, но клетки с размером меньше могут расти случайным образом с флуктуацией размера за счет липидного обмена. Это разделение вызывает накопление клеток ниже размера, что дает более низкий пик в распределении размеров клеток. Эта популяция клеток может служить потенциальным резервуаром для зрелых жировых клеток. Процесс их созревания можно интерпретировать следующим образом: жировые клетки, достигающие критического размера, вследствие случайных колебаний размера могут расти с механизмом роста, зависящим от размера.Как упоминалось выше, изменение размера происходит более активно при диете с высоким содержанием жиров; следовательно, пласт может ускорить процесс созревания в условиях стимуляции.
В модель роста ткани мы включили привлечение новых клеток и рост существующих клеток, но не гибель старых клеток, потому что модель соответствовала данным без апоптоза жировых клеток. Этот результат также согласуется с исследованием, в котором наблюдалось, что жировая ткань придатка яичка мышей C57BL / 6 не демонстрирует значительной гибели клеток до 12 недель при диете с высоким содержанием жиров [33].Однако расширенная диета с высоким содержанием жиров в конечном итоге вызывает апоптоз жировых клеток [33]. Более того, в одном недавнем исследовании сообщается, что жировые клетки человека обновляются в течение десяти лет [7]. Таким образом, наша модель все еще нуждается в улучшении, чтобы ее можно было использовать в различных условиях. Следует учитывать гибель клеток и формально учитывать зависимость параметров модели от диеты. В этом исследовании мы сосредоточились на одном жировом депо, эпидидимальном жире, по нескольким причинам: 1) вес эпидидимальных жировых подушечек может быть определен более точно, чем вес паховых жировых подушечек из-за легкости иссечения; 2) морфология жировых клеток в эпидидимальном жире более однородна, чем в паховом жире, который содержит много клеток, подобных коричневой жировой ткани, особенно у мышей, устойчивых к ожирению, вызванному диетой; и 3) разница между генотипами была более очевидной в росте эпидидимального жира, который достигает плато через 4 недели у мышей FVB, но продолжает расти у мышей C57 на протяжении всего эксперимента, в отличие от пахового жира, который показывает устойчивый рост. у обоих штаммов (рис.S1). Хотя мы не измеряли распределение по размеру клеток других жировых отложений, мы измерили изменение массы паховых и бурых жировых отложений, что показывает аналогичную картину с эпидидимальным жировым депо (рис. S1). Таким образом, представляет интерес применить модель к другим жировым отложениям, которые имеют функциональные различия [34], [35], а также к другим видам, таким как человек, что остается для будущих исследований. Мы ожидаем, что модель может быть применена к таким разнообразным наборам данных, просто настроив параметры модели, потому что модель содержит общие механизмы роста тканей для привлечения новых клеток и их последующего развития.
Наши данные предполагают, что, по крайней мере, три фактора могут объяснить, почему мыши C57 набирают больше жира, чем мыши FVB, при диете с высоким содержанием жиров: во-первых, мыши FVB имеют повышенную скорость метаболизма и повышенную ректальную температуру, скорее всего, из-за повышенного симпатического тонуса. Хотя мы не обнаружили значительных различий в активности между штаммами в нашем исследовании, более подробные измерения поведения показали, что мыши FVB спонтанно гиперактивны по сравнению с мышами C57 [26]. У них также увеличивается частота сердечных сокращений [36], и они реагируют гипергликемией на различные виды лечения [37].Кроме того, они более чувствительны к стрессу, связанному с воздержанием и голоданием [38]. Все эти данные, взятые вместе, предполагают, что активность симпатической нервной системы увеличилась больше у мышей FVB, чем активность у мышей C57. Во-вторых, по сравнению с мышами FVB, мыши C57 очищают циркулирующие триглицериды более эффективно [23], [24], что, по крайней мере, частично можно объяснить более высокой активностью липазы в сыворотке крови [23] и более высокой способностью накапливать триглицериды в печени [23]. ], [24] и жировой ткани (показано в этом исследовании).Хотя молекулярные механизмы клиренса триглицеридов до конца не изучены, жировая ткань явно способствует выведению триглицеридов, поскольку способность выводить циркулирующие триглицериды нарушена у липоатрофных мышей [23]. В частности, сообщалось, что диета с высоким содержанием жиров увеличивает клиренс триглицеридов [39], что может быть связано с индукцией активности липопротеинлипазы в жировой ткани [40]. Наконец, по сравнению с мышами FVB, мыши C57 показали большее количество мелких жировых клеток, особенно при диете с высоким содержанием жиров.Было высказано предположение, что новые жировые клетки могут возникать из клеток-предшественников, которые находятся в депо белого жира взрослых [41], [42], и из других источников, таких как циркулирующие клетки-предшественники костного мозга [43]. Было показано, что рекрутирование обоих типов предшественников стимулируется диетой с высоким содержанием жиров [41], [43]. Возможно, что большее рекрутирование более мелких жировых клеток у мышей C57 может быть вызвано более высоким пулом клеток-предшественников или их более высокой внутренней способностью к дифференцировке адипоцитов.Однако in vitro мезенхимальных стволовых клеток , выделенных из наружного уха мышей FVB и C57, одинаково хорошо дифференцируются в жировые клетки [44]. Попытка сравнить дифференцировку стромальных клеток костного мозга мышей FVB и C57 в жировые клетки не увенчалась успехом из-за очень низкого выхода и плохой пролиферативной способности клеток, выделенных от мышей C57 [45]; однако костный мозг, по-видимому, не является основным источником новых жировых клеток, по крайней мере, у мышей [43]. Наша модель предполагает, что разница между генотипами в рекрутировании малых жировых клеток может быть автономной по отношению к жировым подушечкам, но молекулярный механизм, лежащий в основе этого различия, неясен.Жировая подушечка представляет собой сложный орган, содержащий множество различных типов клеток, включая зрелые жировые, предадипозные и сосудистые клетки, нервы, макрофаги и фибробласты. Количество предшественников адипоцитов и их пролиферация в ответ на внешние сигналы варьируются в зависимости от жировых отложений [35]. Необходимы дальнейшие исследования, чтобы определить, как специфичное для генотипа взаимодействие между разными типами клеток и секретируемыми факторами может влиять на скорость рекрутирования адипоцитов в определенные жировые депо.
Таким образом, мы получили математическую модель, описывающую рост жировой ткани с увеличением количества и размера клеток в зависимости от массы жировой подушечки придатка яичка.Основываясь на этой динамической модели, мы исследовали влияние генетики и диеты на рост жировой ткани. Сравнивая распределение размеров клеток двух штаммов и двух диет, мы пришли к выводу, что изменение размера клеток зависит от диеты, а изменение количества клеток зависит от генетики и диеты, а также от их взаимодействия.
Материалы и методы
Животные
Все процедуры были одобрены Комитетом по уходу и использованию животных Национального института диабета, болезней органов пищеварения и почек.Самцы мышей FVB и C57 были получены из лаборатории Джексона (Бар-Харбор, штат Мэн). Мышей выращивали по четыре в клетке с 12-часовым циклом свет / темнота (свет 06: 00–18: 00). В возрасте 5 недель мышей каждой линии разделили на 2 группы. Половине мышей давали обычную пищу NIH-07 (далее REG; Zeigler Brothers, Inc., Gardners, PA), содержащую 4,08 ккал / г (11% калорий из жира, 62% из углеводов и 26% из белка). Другая половина получала диету с высоким содержанием жиров, F3282 (далее HF; Bio-Serv, Frenchtown, NJ), содержащую 5.45 ккал / г (59% жиров, 26% углеводов и 15% белков). Вода и диета были предоставлены ad libitum . Было проведено пять независимых экспериментов, в каждой из которых использовалось 4 группы мышей: FVB REG, FVB HF, C57 REG и C57 HF. В трех экспериментах мышей содержали на контролируемой диете в течение 12 недель и использовали для анализа состава тела, физиологической характеристики и распределения клеток по размеру. Две дополнительные группы мышей были умерщвлены после 2 недель и 4 недель контрольного кормления с высоким содержанием жира только для определения распределения клеток по размеру.
Эксперименты in vivo
Состав тела, потребление пищи, скорость метаболизма, толерантность к глюкозе, клиренс триглицеридов и окисление жирных кислот в изолированной камбаловидной мышце измеряли, как описано ранее [46]. Окисление жирных кислот всего тела измеряли, как описано в Gautam et al. [47]. Кровь для биохимических анализов брали из хвостовой вены без голодания. Уровни глюкозы измеряли с помощью Glucometer Elite (Bayer, Elkhart, IN). Сывороточный инсулин анализировали с помощью радиоиммуноанализа (Linco Research, St.Чарльз, Миссури). Триглицериды сыворотки, холестерин (Thermo DMA, Луисвилл, Колорадо) и свободная жирная кислота (FFA) (Roche Applied Science, Индианаполис, Индиана) измеряли в соответствии с процедурами производителя.
Измерение размера клеток эпидидимального жира
Распределение размеров клеток эпидидимального жира измеряли через 2, 4 и 12 недель высокожирового и контрольного кормления с использованием Beckman Coulter Multisizer III, как описано ранее [9]. Вкратце, 20–30 мг жировой ткани были взяты из средней части путем рассечения и последующего удаления образца для фиксации из центра разрезанного эпидидимального жира.Образцы тканей немедленно фиксировали в четырехокиси осмия [48], инкубировали на водяной бане при 37 ° C в течение 48 ч, а затем определяли размер жировых клеток на приборе Beckman Coulter Multisizer III с апертурой 400 µ м. Диапазон размеров ячеек, которые можно эффективно измерить с помощью этой апертуры, составляет 20–240 µ м. Прибор был настроен на подсчет 6000 частиц, и суспензия фиксированных клеток была разбавлена так, чтобы совпадающий счет был <10%. После сбора размеров импульсов данные были выражены в виде диаметров частиц и отображены в виде гистограмм отсчетов по диаметру с использованием линейных интервалов и линейной шкалы для оси x ().Распределение клеток по размерам было измерено в четырех образцах из каждой группы, за исключением мышей C57 после 4-недельного воздействия диеты с высоким содержанием жиров, у которых было только три доступных образца. Образец был взят из каждой жировой подушечки и обработан отдельно. Затем каждый образец был посчитан как минимум дважды. Затем кривые для двух образцов усредняются, но только после проверки воспроизводимости между двумя образцами.
Модель роста жировой ткани
Распределение размеров клеток включает всю информацию, относящуюся к размерам клеток в ткани, и его изменения дают статистическое представление о детальном процессе роста каждой клетки.Чтобы изучить рост жировой ткани с точки зрения лежащих в основе микроскопических процессов, мы рассматриваем математическую модель, количественно определяющую процессы, которые изменяют распределение клеток по размерам. Модель может предсказать, сколько новых клеток образуется и как клетки разного размера растут по мере увеличения массы жировой ткани. Плотность количества клеток определенного размера (диаметра) при данной массе жировой подушечки — это конкретное количество, которое необходимо учитывать. Мы рассматриваем, как это распределение клеток по размерам изменяется с постепенным изменением массы жировой ткани.Эволюция распределения клеток по размерам с массой жировой ткани может быть смоделирована уравнением в частных производных:
(1)
Это уравнение включает три основных компонента процесса роста жировой ткани. Во-первых, мы предполагаем, что рекрутирование новых клеток происходит только при минимальном наблюдаемом размере клетки, который математически выражается как дельта-функция. Скорость набора по отношению к массе жировой подушечки определяется экспоненциальной функцией:
(2)
, где — исходное общее количество клеток при данной начальной массе жировой подушечки, а — скорость увеличения количества клеток на единицу изменения. в массе жировой подушечки.Изменение общего числа клеток — это скорость пополнения новых клеток, если гибель клеток незначительна; мы не обнаружили необходимости включать апоптоз в клетки любого размера, чтобы соответствовать этим экспериментальным данным. Следовательно, этот коэффициент пополнения может быть непосредственно получен из экспериментального результата, используя соотношение между общим числом клеток и массой жировой подушечки путем дифференцирования функции по отношению к. Во-вторых, рост клеток зависит от размера клетки. После созревания жировых клеток до определенного размера они могут накапливать жир, вызывая гипертрофию.В дополнение к этой ограничивающей скорости роста маленьких жировых клеток, может также существовать верхний предел роста, потому что большие жировые клетки не могут расти бесконечно в результате этого процесса роста, хотя они могут достигать больших размеров из-за колебаний размера, вызванных обменом липидов. Повышение и снижение скорости роста клеток в зависимости от размера клетки можно описать с помощью общей функциональной формы, умножающей две сигмоидальные функции,
(3)
, где представляет максимальную скорость роста; и — соответственно нижний и верхний критические размеры, которые дают половину максимальной скорости роста; и укажите их масштаб ().Наконец, последний член в формуле. (1) представляет колебания размера клеток с постоянной скоростью, которые отражают обмен липидов, случайным образом происходящий в жировых клетках. Этот липидный оборот является единственным механизмом роста для крупных клеток, размер которых превышает верхний критический размер. Как правило, рост клеток, зависящий от размера, приводит к увеличению их распределения по размерам, в то время как колебания размеров распространяют распределение по размерам.
Метод оптимизации
Чтобы оптимизировать параметры модели, чтобы они могли точно описывать эволюцию распределения размеров ячеек в эксперименте, мы использовали минимизацию «функции стоимости», которая количественно определяет отклонение между моделью и результатами эксперимента.Чтобы определить функцию стоимости, нормализованное распределение размеров клеток при заданной массе жира сравнивалось с данными моделирования с набором параметров:
(6)
, где — общее количество интервалов размера клеток, а — общее количество заданных масса жира. Масштаб функции стоимости был рассчитан на основе внутренней флуктуации экспериментальных данных, которую можно определить как квадрат отклонения между измеренным распределением ячеек по размерам и функцией плавной аппроксимации:
(7)
Эта внутренняя флуктуация численно составляет около 10 процентов квадрата отклонения между экспериментальными и модельными данными.В качестве функции аппроксимации мы использовали сумму двух экспонент и гауссову форму,
(8)
, форму, которая использовалась для аппроксимации распределений размеров жировых клеток [9]. Подбор этих параметров был выполнен с использованием процедуры подбора нелинейной кривой в MATLAB R2007a (Натик, Массачусетс, США). Для процесса оптимизации мы специально использовали метод Монте-Карло с параллельным темперированием, чтобы найти глобальный минимум функции стоимости [50]. Мы использовали 10 равномерно распределенных значений (от 0,1 до 1) для параметра темперирования и запускали десять цепочек параллельно с вероятностью обновления.На каждых 20 шагах случайным образом выбиралась пара смежных имитаций по десяти параметрам отпуска, и их состояния параметров менялись с вероятностью. После достижения равновесия было использовано 20 000 итераций с фиксированным параметром отпуска для оценки оптимальных значений параметров и их стандартных ошибок.
Мы также использовали этот метод для оценки исходного общего количества клеток, и скорости его увеличения, исходя из соотношения между жировой массой и общим количеством клеток (). При минимизации функции аппроксимации и экспериментальных данных мы использовали ограничение, согласно которому начальное количество клеток одинаково независимо от условий диеты, т.е.е., обычные и жирные диеты. Средняя жировая масса четырех контрольных мышей перед регулярной диетой и диетой с высоким содержанием жиров была использована в качестве начальной жировой массы, которая составляет 0,34 г и 0,29 г для мышей FVB и C57, соответственно. Мы оценили погрешности и, распространив 10-процентные статистические флуктуации, наблюдаемые в экспериментальных данных.
Численное решение дифференциального уравнения с частными производными
Мы решили следующую дискретную версию нашей модели, заданную как непрерывное дифференциальное уравнение с частными производными в формуле.(1):
(9)
с интервалом масс δm = 0,1 мг и интервалом размеров δs = 0,73 µ м.
Динамика роста жировой ткани
Abstract
Рост жировой ткани происходит по двум механизмам: гиперплазия (увеличение количества клеток) и гипертрофия (увеличение размера клеток). Генетика и диета влияют на относительный вклад этих двух механизмов в рост жировой ткани при ожирении. В этом исследовании распределение по размеру эпидидимальных жировых клеток от двух линий мышей, устойчивых к ожирению FVB / N и склонных к ожирению C57BL / 6, измеряли через 2, 4 и 12 недель в условиях регулярного кормления и питания с высоким содержанием жира.Общее количество клеток в жировой подушке придатка яичка оценивали по массе жировой подушечки и нормализованному распределению клеток по размеру. Число клеток и средневзвешенный размер клеток увеличиваются в зависимости от массы жировой прослойки. Чтобы точно решить проблему роста жировой ткани, мы разработали математическую модель, описывающую эволюцию распределения размеров жировых клеток как функцию увеличения массы жировой ткани, а не увеличения хронологического времени. Наша модель описывает привлечение новых жировых клеток и их последующее развитие в разных штаммах и с разными режимами питания, с общими механизмами, но с параметрами модели, зависящими от диеты и генетики.По сравнению со штаммом FVB / N, штамм C57BL / 6 имеет большее количество мелких жировых клеток. Гиперплазия усиливается за счет диеты с высоким содержанием жиров в зависимости от штамма, что предполагает синергетическое взаимодействие между генетикой и диетой. Кроме того, кормление с высоким содержанием жиров увеличивает скорость роста жировых клеток, независимо от напряжения, что отражает увеличение количества калорий, требующих хранения. Кроме того, диета с высоким содержанием жиров приводит к резкому увеличению распределения жировых клеток по размерам у обоих штаммов; это означает увеличение колебаний размера жировых клеток за счет липидного обмена.
Авторское резюме
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. Динамическое изменение жировой ткани in vivo невозможно отслеживать в режиме реального времени из-за текущих технических ограничений. Однако мы можем измерить распределение жировых клеток по размеру у отдельных животных. Наша основная цель — извлечь динамические характеристики ремоделирования тканей из снимков распределения клеток по размерам.Мы разрабатываем математическую модель, которая интерполирует измерения распределения клеток по размеру и предсказывает непрерывное изменение распределения клеток по размеру в зависимости от увеличения массы жировых подушечек. Наша модель роста жировой ткани включает три основных компонента: рекрутирование новых клеток, рост клеток в зависимости от размера и колебания размера клеток. В частности, мы сравнили рост жировой ткани у склонных к ожирению и устойчивых к ожирению мышей при стандартной диете или диете с высоким содержанием жиров, чтобы изучить генетическое и диетическое влияние на рост жировой ткани.Применяя нашу модель к этим различным условиям, мы обнаружили, что увеличение размера жировых клеток зависит от диеты. С другой стороны, увеличение количества жировых клеток, вызванное диетой, зависит от деформации, что предполагает синергию между генетикой и диетой.
Введение
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. На фенотип ожирения жировой ткани влияют диета и генетика, а также их взаимодействие [1] — [4].Начиная с исследований Джонсона и Хирша [5], существует обширная литература о росте жировой ткани при нормальном и патологическом развитии, характеризующая состояние ткани с точки зрения среднего размера и количества клеток. Гиперпластический рост появляется только на ранних стадиях развития жировой ткани [6], [7]. Гипертрофия возникает до гиперплазии, чтобы удовлетворить потребность в дополнительных накоплениях жира при прогрессировании ожирения [8]. Однако оказалось трудно понять, как диета и генетика конкретно влияют на гиперплазию и / или гипертрофию жировых клеток из-за ограниченных данных о росте жировой ткани.
Помимо этих исследований, недавно стало возможным точно измерить распределение клеток по размерам. Эта подробная информация, по сравнению со средним размером клеток и общим числом клеток, может использоваться для вычисления многих величин, связанных с размером, которые позволяют более точно охарактеризовать процесс роста жировой ткани. Кумулянты распределения клеток по размерам могут использоваться для вычисления других физиологических величин, таких как средневзвешенный размер клеток. Распределение клеток по размерам можно использовать для оценки общего количества клеток в жировой подушке по ее массе.Более того, считается, что некоторые специфические метаболические свойства, например, резистентность к инсулину [9] и секреция адипокина [10], зависят от точного распределения клеток по размерам, а не от среднего размера клеток. Действительно, в нескольких исследованиях изучается изменение распределения жировых клеток по размерам в различных условиях при развитии куриного эмбриона [11], у худых и страдающих ожирением крыс Zucker [12], [13], крыс линии Wistar с частичной липэктомией [14], биопсии кролика [ 15] и жировой ткани человека [16], [17]. Эти исследования были сосредоточены только на статических различиях между распределениями клеток по размерам в разных условиях.Однако статические распределения размеров клеток в поперечном сечении для ряда снимков развития животных можно использовать для вывода динамики роста жировой ткани, если мы сможем надлежащим образом проанализировать снимки с помощью математического моделирования. Учитывая нынешние технические ограничения, это может быть лучший доступный подход к микроскопическому и продольному пониманию роста жировой ткани in vivo , хотя в недавнем эксперименте были получены микроскопические наблюдения накопления липидов в липидных каплях жировых клеток [18].
Для изучения генетического и диетического влияния на динамический процесс роста жировой ткани мы получили распределения по размерам клеток эпидидимального жира у устойчивых к ожирению линий мышей FVB / N (далее FVB) и склонных к ожирению линий мышей C57BL / 6 (C57) в соответствии с стандартные диеты и диеты с высоким содержанием жиров. Мышь C57 является наиболее охарактеризованной моделью ожирения, вызванного диетой [19], а мышь FVB является предпочтительной моделью для создания трансгенных мышей [20]. Эти два широко используемых инбредных штамма мышей генетически довольно далеки [21], [22] и имеют различные метаболические фенотипы: по сравнению с мышами FVB, мыши C57 имеют низкие уровни циркулирующих триглицеридов [21] и повышенный клиренс триглицеридов [23], [24]; Мыши FVB характеризуются относительно более высокой печеночной инсулинорезистентностью, контррегуляторной реакцией на гипогликемию и сниженной глюкозо-стимулированной секрецией инсулина [25]; Также известно, что мыши FVB являются спонтанно гиперактивными [26] и относительно худыми, поскольку они, по-видимому, менее восприимчивы к диете с высоким содержанием жиров, чем мыши C57 [27].Однако развитие ожирения, вызванного диетой, у этих двух штаммов формально не сравнивалось. В этом исследовании мы разработали математическую модель, прогнозирующую изменение распределения клеток по размерам в зависимости от массы жировой подушечки придатка яичка, для количественного анализа динамических характеристик, зависящих от генетики и / или диеты. Модель роста жировой ткани описывает, сколько новых клеток образуется, как каждая клетка растет в зависимости от ее размера и как липидный обмен приводит к колебаниям размера, которые вызывают расширение распределения клеток по размерам.По мере увеличения массы эпидидимальных жировых подушек распределение клеток по размерам систематически изменяется в зависимости как от генетики, так и от диеты. Сравнивая экспериментальные результаты с теоретической моделью роста, мы обнаружили, что гипертрофия сильно коррелирует с диетой. С другой стороны, гиперплазия зависит от генетики. Изменения при гиперплазии, вызванные диетой, зависят от штамма, что предполагает взаимодействие между диетой и генетикой.
Результаты
Влияние диеты с высоким содержанием жиров на состав тела мышей FVB и C57
В начале эксперимента (в возрасте 5 недель) мыши C57 были значительно легче, чем мыши FVB () из-за разницы в худой масса, хотя общая жировая масса не отличалась ().Когда мышей поддерживали на обычном питании, разница в массе тела исчезла к возрасту 11 недель (6-я неделя эксперимента). В условиях обычной диеты мыши FVB и C57 сохраняли сопоставимую массу жира на протяжении всего эксперимента (). Диета с высоким содержанием жиров вызвала значительное увеличение массы тела и жировой массы у обоих штаммов; однако изменения массы тела и жировой массы были более значительными у мышей C57. У мышей C57 было вдвое больше жира после 12 недель кормления с высоким содержанием жиров ().Общая разница в общей массе жира между мышами FVB и C57 коррелировала с пропорциональными различиями в количестве эпидидимального (внутрибрюшного), пахового (подкожного) и бурого жира (). Потребление калорий и активность были сопоставимы у мышей FVB HF и C57 HF; однако мыши FVB HF имели более высокий уровень покоя и общего потребления кислорода, а также более высокую ректальную температуру, что позволяет предположить, что увеличение расхода энергии, а не снижение потребления калорий, было причиной относительной устойчивости к ожирению, вызванному диетой с высоким содержанием жиров, у мышей FVB.Интересно, что в течение первых 2 недель кормления с высоким содержанием жиров мыши FVB и C57 показали сопоставимое увеличение общей жировой массы (). Мыши C57 HF продолжали быстро увеличивать жировую массу до 10 недели эксперимента, тогда как мыши FVB HF замедляли накопление жира около недели 3. У мышей C57 кормление с высоким содержанием жира вызывало постепенное увеличение как эпидидимальных, так и паховых жировых подушечек; Напротив, у мышей FVB жировая масса придатка яичка увеличилась лишь незначительно после 4 недель кормления с высоким содержанием жира, в то время как паховая жировая подушечка продолжала увеличиваться в размере на протяжении всего эксперимента (рис.S1). Кормление с высоким содержанием жиров вызывало значительное повышение уровня глюкозы и инсулина в крови как у мышей FVB, так и у мышей C57 (). Уровни инсулина и непереносимость глюкозы были выше у мышей C57 HF, чем у мышей FVB HF, что свидетельствует о более тяжелой инсулинорезистентности (). В соответствии с предыдущими сообщениями [23], [24], мыши C57 REG показали пониженные уровни триглицеридов в сыворотке по сравнению с мышами FVB REG без разницы в FFA (). Это происходило не из-за более высокой утилизации жира, так как коэффициент респираторного обмена () и скорость окисления жирных кислот, измеренная in vivo, () и в изолированных скелетных мышцах () были сопоставимы у мышей FVB REG и C57 REG.Более вероятно, что более низкие уровни триглицеридов в сыворотке у мышей C57 REG были вызваны гораздо более эффективным клиренсом циркулирующих триглицеридов, как предполагает тест на клиренс триглицеридов (). Кормление с высоким содержанием жиров снижало уровни циркулирующих триглицеридов как у мышей FVB, так и у мышей C57 и улучшало клиренс триглицеридов у последних штаммов (и). Оба штамма показали сопоставимое снижение коэффициента респираторного обмена, что свидетельствует о сопоставимом увеличении утилизации жирных кислот в условиях диеты с высоким содержанием жиров (). Взятые вместе, эти данные предполагают, что способность эффективно выводить триглицериды из кровотока может способствовать высокой способности накопления жира у мышей C57.
Изменения в составе тела мышей FVB и C57 в условиях стандартного корма (REG) и с высоким содержанием жира (HF).(A) Масса тела. (B) Жировая масса. Состав тела измеряли в указанные моменты времени у 8 мышей в группе с использованием МРТ-анализатора Echo 3-in-1. Высокожировое и контрольное кормление начали в возрасте 5 недель. Ось X показывает недели контролируемого кормления. Показан один из трех независимых экспериментов. Приведенные значения являются средними ± SEM.
Физиологические характеристики мышей FVB и C57.(A) Тест на толерантность к глюкозе был проведен на 10-й неделе контролируемого кормления с пищей и диетой с высоким содержанием жиров. Мышей не кормили в течение ночи и вводили глюкозу (2 мг / г, внутрибрюшинно). Каждая группа была представлена 8 мышами. (B) Окисление олеиновой кислоты в целом организме измеряли у самцов мышей FVB ad C57 в возрасте 10 недель в не голодном состоянии (8 мышей на группу), как описано в Gautam et al. [47]. (C) Окисление пальмитиновой кислоты измеряли в камбаловидной мышце, взятой у 10-недельных мышей FVB и C57, которых содержали на диете (8 мышей на группу).Образцы собирали в 8 часов утра у мышей, которых случайным образом кормили, или мышей, голодавших в течение 18 часов [методы, описанные в Toyoshima et al. [46]]. (D) Клиренс триглицеридов у самцов мышей FVB и C57 после 11 недель контролируемого кормления кормом и диетой с высоким содержанием жиров. Мышей не кормили в течение 4 часов, а затем вводили через желудочный зонд 400 мкл л арахисового масла. Плазму собирали ежечасно в течение 5 часов из хвостовой вены для измерения триглицеридов. Каждая группа была представлена 8 мышами.
Таблица 1
Характеристики мышей FVB и C57 после 12 недель высокожирового и контрольного кормления.
| FVB REG a | FVB HF a | C57 REG a | C57 HF | a 94 996| 912 912 912 912 912 Взаимодействие б | | ||||||||||
| Масса тела (г) | 29,1 ± 0,4 | 35,4 ± 1,4 | 29,1 ± 0,4 | 43,0 ± 1,8 | 0,003 | <0,001 | 0,003 | г)0.47 ± 0,03 | 1,23 ± 0,11 | 0,67 ± 0,08 | 2,00 ± 0,21 | <0,001 | <0,001 | 0,032 | |
| Паховой жир (г) | 0,3412 ± 0,02 | 0,190,3412 ± 0,02 | 0,3412 ± 0,02 | 0,45 ± 0,041,92 ± 0,27 | 0,003 | <0,001 | <0,003 | ||||||||
| Коричневый жир (г) | 0,14 ± 0,01 | 0,27 ± 0,04 | 0,1412 ± 0,01 | 0,01 0,09 0,003<0.001 | 0,003 | ||||||||||
| Потребление пищи (ккал / мышь / день) | 16,6 ± 1,0 | 18,0 ± 1,3 | 13,2 ± 0,8 | 17,7 ± 0,8 | 0,07 | 0,007 92| 3 ns Общее потребление кислорода (мл / г) | 9,5 ± 0,2 | 11,6 ± 0,3 | 9,0 ± 0,2 | 9,4 ± 0,2 | <0,001 | <0,001 | нс | | |
| Потребление кислорода в покое (мл / г) ) | 8.3 ± 0,2 | 9,7 ± 0,3 | 7,3 ± 0,2 | 8,1 ± 0,3 | <0,001 | <0,001 | 0,003 | ||||||||
| Коэффициент респираторного обмена | 0,86 ± 0,01 | 0,75 ± 0,01 0,01 | 0,72 ± 0,00 | нс | <0,001 | 0,006 | |||||||||
| Суммарная активность (торможение луча / мин) | 265 ± 5 | 190 ± 17 | 345 ± 41 | 122011293 нс | <0.001 | нс | |||||||||
| Ходьба (торможение луча / мин) | 105 ± 3 | 53 ± 10 | 144 ± 25 | 38 ± 5 | нс | <0,002 | 91 Ректальная температура (° C) | 36,0 ± 0,2 | 36,6 ± 0,2 | 35,4 ± 0,2 | 35,5 ± 0,2 | <0,001 | 0,057 | нс | |
| Триглицерид сыворотки (мг / дл) | ± 25 | 95 ± 24 | 89 ± 16 | 64 ± 6 | 0.002 | 0,004 | ns | ||||||||
| Свободные жирные кислоты (мМ) | 0,22 ± 0,03 | 0,22 ± 0,01 | 0,26 ± 0,04 | 0,27 ± 0,05 | 1292 ns | ns | ns Глюкоза крови (мг / дл) | 187 ± 16 | 225 ± 11 | 209 ± 11 | 247 ± 15 | нс | 0,009 | нс | |
| Инсулин сыворотки (мг / | 1,1) 912 ± 0,1 | 1,9 ± 0,3 | 2.2 ± 0,8 | 4,9 ± 1,2 | 0,011 | 0,033 | нс | ||||||||
| Клиренс триглицеридов (AUG c ) | 3238 ± 476 | 1651 ± 160 | 48 | 489 | <0,001 | 0,011 | <0,001 | ||||||||
| Толерантность к глюкозе (AUG d ) | 28633 ± 1372 | 39560 ± 3217 | 25947 ± 1212 | 001 | 0,003 |
Гипертрофия и гиперплазия с увеличением жировой массы
Чтобы проверить основной механизм различной скорости накопления жира в эпидидимальном жире мышей FVB и C57, мы измерили распределение массы и размеров клеток в образцах тканей эпидидимальный жир, собранный на 0, 2, 4 и 12 неделях контролируемого кормления (). Поскольку гистологический анализ не позволяет точно определить размер клеток адипоцитов, который будет обсуждаться позже, мы измерили распределение клеток по размеру с помощью счетчика Коултера и рассчитали средневзвешенный размер клеток и общее количество клеток эпидидимальной жировой подушки по этим измерениям, которые аналогичные значения в другом исследовании на мышах [5].Сильная корреляция наблюдалась между массой жировых подушечек и средневзвешенным размером клеток, а также между массой жировых подушечек и общим числом клеток, независимо от напряжения и разницы в диете (). Первая корреляция дала масштабное соотношение между массой жировой подушечки и средневзвешенным по объему размером клеток (). Кроме того, была обнаружена экспоненциальная зависимость между массой жировой подушечки, и общим числом клеток, где начальная масса жировой подушки была получена от контрольных мышей; и начальное количество клеток, и скорость увеличения количества клеток в массе жировой подушечки, были оценены по данным (;).Первоначальное количество клеток у мышей C57 было больше, чем исходное количество клеток у мышей FVB (фиг. S2C и S2D). По мере увеличения массы жировой ткани увеличивается общее количество клеток. Скорость увеличения числа клеток была выше при обычном питании, чем при диете с высоким содержанием жиров, и эта тенденция более очевидна у мышей C57 (;), что указывает на генетическое различие. Соотношение между результатами регулярной диеты и диеты с высоким содержанием жиров составляет 1,42 и 3,22 для мышей FVB и C57 соответственно (). Это может указывать на взаимодействие между генетикой и диетой при увеличении количества клеток.Обратите внимание, что мы также наблюдали аналогичные результаты с массой тела и массой жира, поскольку три величины (масса жировых подушечек придатка яичка, масса жира и масса тела) коррелируют друг с другом. Однако результаты с массой эпидидимальной жировой подушечки были наиболее подходящими: среднеквадратичное отклонение между данными и соответствием составило 9,73, 7,94, 3,56 × 10 5 и 5,54 × 10 6 , соответственно; результат с массой тела составил 11,82, 15,35, 3,65 × 10 5 и 4,82 × 10 6 ; результат с жировой массой 8.58, 8,61, 4,28 × 10 5 и 6,11 × 10 6 .
Нормализованное распределение клеток по размерам в жировой подушке придатка яичка.На графике нанесен один типичный пример контрольных мышей FVB в возрасте пяти недель. Кружки обозначают точки измеренных данных, а линия представляет собой аппроксимирующую кривую, которая представляет собой сумму одной гауссовой и двух экспоненциальных функций.
Таблица 2
Значения параметров модели роста жировой ткани.
Генетические и диетические эффекты на рост жировой ткани
Эти две сильные корреляции между массой жировой подушечки и гипертрофией и между массой жировой подушечки и гиперплазией позволяют предположить, что увеличение жировой ткани можно описать как систематический процесс роста в отношении для увеличения массы жировой прослойки.Мы расположили распределения клеток по размерам, отсортированные по массе эпидидимальных жировых подушечек (). Примечательно, что модель роста жировой ткани в формуле. (1) описывает эволюционирующую картину распределения клеток по размерам в зависимости от увеличения массы жировой прослойки. Модель количественно соответствовала экспериментальным распределениям размеров клеток, несмотря на то, что все распределения представляют собой данные поперечного сечения, полученные от отдельных животных. Различные значения параметров в модели, которые соответствуют каждому индивидуальному распределению размеров клеток для обоих штаммов и обоих режимов питания, дали количественные различия в процессе роста эпидидимальной жировой ткани между штаммами и между диетами ().Во-первых, максимальная зависящая от размера скорость роста и скорость колебаний размера клеток из-за обмена липидов продемонстрировали разницу, вызванную диетой, и меньшую разницу, вызванную деформацией. Зависимый от размера рост и колебания размера, приводящие к гипертрофии, по-видимому, регулируются в основном диетой. В частности, рост, зависящий от размера, сдвинул режим больших клеток в распределении размеров клеток в сторону больших размеров, а флуктуации липидного обмена увеличили разброс распределения вокруг режима больших клеток.Важно отметить, что результаты должны быть тщательно интерпретированы, потому что каждая скорость — это скорость на единицу увеличения массы жировой ткани, а не на единицу увеличения времени. Во-вторых, геометрические параметры (, и), которые определяют форму скорости роста, зависящей от размера, имели практически одинаковые значения независимо от диеты и разницы деформаций, за исключением. Следовательно, нижний критический размер, который дает размер, инициирующий рост клеток в зависимости от размера, и два параметра масштаба могут быть зафиксированы на с l = 37 µ m, η l = 12 µ м и η u = 63 µ м соответственно.С другой стороны, верхний критический размер, ограничивающий зависимый от размера клетки рост больших клеток, зависел от диеты; при кормлении с высоким содержанием жира этот пороговый размер сместился в сторону большего размера (). При диете с высоким содержанием жиров изменения параметров (,, и) увеличивают способность жировых тканей накапливать липиды за счет как гиперплазии, так и гипертрофии. Более низкие уровни триглицеридов в сыворотке при условии диеты с высоким содержанием жиров (и) могут коррелировать с увеличением накопления липидов в увеличенных жировых клетках, потому что не было обнаружено значительных различий в окислении жирных кислот, о чем свидетельствует отсутствие различий в соотношении респираторного обмена ().
Изменения распределения клеток по размерам с увеличением массы эпидидимальной жировой подушечки при кормлении и диетах с высоким содержанием жиров.Начальные распределения размеров клеток пятинедельных контрольных мышей приведены в первой строке каждого столбца пунктирными линиями. Точки показывают экспериментальные результаты. Обратите внимание, что результаты упорядочены по массе эпидидимальной жировой подушечки, а не по хронологическому времени (недели контролируемого кормления), указанному в скобках. Сплошные линии представляют собой нормализованные распределения клеток по размеру, соответствующие данной массе эпидидимальной жировой подушечки, предсказанной моделью роста жировой ткани.
Зависимая от размера скорость роста массы придатка яичка.Здесь для этого графика используются средние значения параметров. Символы показаны для облегчения сравнения строк.
Обсуждение
Наше центральное открытие состоит в том, что гиперплазия и гипертрофия жировых клеток в эпидидимальной жировой подушке является функцией массы жировой подушечки, даже несмотря на то, что у отдельных животных может потребоваться разное время для достижения заданной массы жировой подушечки. Следовательно, рост жировой ткани, представленный как изменения распределения клеток по размерам, можно систематически моделировать как процесс роста по отношению к увеличению массы жировой подушечки; это может отражать корреляцию между массой жировой подушечки и секрецией адипокинов и других сигнальных молекул, контролирующих рост жировой ткани.Соответственно, следует отметить, что нормы в нашей модели — это не обычные нормы увеличения на единицу времени, а нормы увеличения на единицу массы. Таким образом, несколько показателей (, и) в модели имели более высокие значения для животных, соблюдающих диету, чем для животных, соблюдающих диету с высоким содержанием жиров. Однако, если бы эти показатели были преобразованы в обычные нормы увеличения на единицу времени, они имели бы большие значения для диеты с высоким содержанием жиров, потому что требуется меньше времени для увеличения массы жировой подушечки на единицу из более крупных и более многочисленных клеток на диета с высоким содержанием жиров, чем для увеличения такой же величины за счет меньшего и меньшего количества клеток на диете (рис.S1). Было высказано предположение, что когда ожирение прогрессирует, рано возникает гипертрофия жировых клеток, а затем запускается гиперплазия [8]. Наше исследование показало, что рекрутирование новых клеток экспоненциально увеличивается с увеличением массы жировой ткани. Гипертрофия жировых клеток является основным фактором увеличения массы жировой прослойки, тогда как гиперплазия не способствует этому увеличению, потому что она возникает в небольших клетках, в которых хранится гораздо меньший объем жира. Таким образом, наша модель естественным образом воплощает идею о том, что на гиперплазию влияет гипертрофический рост клеток.С другой стороны, сообщалось, что гиперплазия жировых клеток возникает только на ранних стадиях развития; следовательно, не следует ожидать рекрутирования новых клеток на поздних стадиях даже в условиях ожирения [6], [7]. Возможно, возраст животных в нашем исследовании (6 недель) допускает возникновение гиперплазии.
Модель, разработанная здесь, может дать микроскопическое представление о росте жировых клеток, зависящем от размера, который не может быть рассмотрен статическими поперечными исследованиями.Например, мы обнаружили следующие специфические свойства роста клеток, зависящие от размера: нижний критический размер, инициирующий накопление липидов, не зависел от диеты у двух линий мышей, тогда как верхний критический размер ограничивал рост клеток до достижения необходимого уровня. необычайный размер, был увеличен на диете с высоким содержанием жиров. Эта зависимость роста клеток от размера является проверяемой гипотезой. Далее, параметр флуктуации размера клеток различался при обычном питании и рационе с высоким содержанием жиров; оно больше при диете с высоким содержанием жиров, когда оно преобразуется в единицы, соответствующие изменению единицы времени, а не единице изменения массы жировой подушечки.Таким образом, случайный процесс высвобождения и поглощения жира жировыми клетками происходит более активно при диете с высоким содержанием жиров, чем при обычной диете. Может быть интересно посмотреть, можно ли распространить эти результаты на другие штаммы и организмы.
По сравнению с исследованиями, в которых наблюдали единственный пик в распределении размеров клеток жировых клеток [11], [15] — [17], [28], мы наблюдали бимодальные распределения клеток по размерам, как сообщалось другими [12], [13], [29] — [32]. В большинстве исследований [12], [13], [31], [32] по наблюдению за бимодальностью использовалась технология счетчика Коултера, которая имеет несколько преимуществ для оценки всего распределения размеров ячеек [31]: Во-первых, можно доказать, что анализируемые ячейки быть подлинными жировыми клетками на основе морфологии и флотации; во-вторых, объем каждой клетки оценивается независимо от формы и без артефактов смещения центра, что является правилом, а не исключением, с использованием гистологических подходов; наконец, можно подсчитать и измерить достаточное количество частиц, чтобы получить статистически значимые комплексные кривые.Напротив, микроскопические методы гистологии могут не наблюдать мелкие клетки из-за влияния увеличения микроскопа [30], небольшого количества образцов и систематической ошибки выборки. Однако при использовании счетчика Коултера контаминация, не связанная с адипоцитами, может способствовать распределению клеток по размерам, особенно при малых размерах, хотя наш минимальный диаметр клеток, 22 µ м, превышает возможные диапазоны контаминации, от 10 до 20 µ м, упомянутый Mersmann и MacNeil [31]. Чтобы быть уверенным, мы снова проанализировали модифицированные данные, используя только распределения размеров клеток выше 35 µ м в диаметре с моделью, и пришли к тем же выводам (данные не показаны).Надир в распределении размеров клеток () может разделять две популяции клеток. ДеМартини и Франсендез определили маленькие клетки с диаметром менее 35 µ м как «очень маленькие жировые клетки» [29]. Согласно нашей модели, эти клетки имеют незначительный рост в зависимости от размера, поскольку их размер меньше нижнего критического размера, с l = 37 µ m. Следовательно, механизм роста, зависящий от размера, может естественным образом объяснить происхождение бимодальности в распределении жировых клеток по размеру.Клетки с размером только выше могут расти в зависимости от размера, но клетки с размером меньше могут расти случайным образом с флуктуацией размера за счет липидного обмена. Это разделение вызывает накопление клеток ниже размера, что дает более низкий пик в распределении размеров клеток. Эта популяция клеток может служить потенциальным резервуаром для зрелых жировых клеток. Процесс их созревания можно интерпретировать следующим образом: жировые клетки, достигающие критического размера, вследствие случайных колебаний размера могут расти с механизмом роста, зависящим от размера.Как упоминалось выше, изменение размера происходит более активно при диете с высоким содержанием жиров; следовательно, пласт может ускорить процесс созревания в условиях стимуляции.
В модель роста ткани мы включили привлечение новых клеток и рост существующих клеток, но не гибель старых клеток, потому что модель соответствовала данным без апоптоза жировых клеток. Этот результат также согласуется с исследованием, в котором наблюдалось, что жировая ткань придатка яичка мышей C57BL / 6 не демонстрирует значительной гибели клеток до 12 недель при диете с высоким содержанием жиров [33].Однако расширенная диета с высоким содержанием жиров в конечном итоге вызывает апоптоз жировых клеток [33]. Более того, в одном недавнем исследовании сообщается, что жировые клетки человека обновляются в течение десяти лет [7]. Таким образом, наша модель все еще нуждается в улучшении, чтобы ее можно было использовать в различных условиях. Следует учитывать гибель клеток и формально учитывать зависимость параметров модели от диеты. В этом исследовании мы сосредоточились на одном жировом депо, эпидидимальном жире, по нескольким причинам: 1) вес эпидидимальных жировых подушечек может быть определен более точно, чем вес паховых жировых подушечек из-за легкости иссечения; 2) морфология жировых клеток в эпидидимальном жире более однородна, чем в паховом жире, который содержит много клеток, подобных коричневой жировой ткани, особенно у мышей, устойчивых к ожирению, вызванному диетой; и 3) разница между генотипами была более очевидной в росте эпидидимального жира, который достигает плато через 4 недели у мышей FVB, но продолжает расти у мышей C57 на протяжении всего эксперимента, в отличие от пахового жира, который показывает устойчивый рост. у обоих штаммов (рис.S1). Хотя мы не измеряли распределение по размеру клеток других жировых отложений, мы измерили изменение массы паховых и бурых жировых отложений, что показывает аналогичную картину с эпидидимальным жировым депо (рис. S1). Таким образом, представляет интерес применить модель к другим жировым отложениям, которые имеют функциональные различия [34], [35], а также к другим видам, таким как человек, что остается для будущих исследований. Мы ожидаем, что модель может быть применена к таким разнообразным наборам данных, просто настроив параметры модели, потому что модель содержит общие механизмы роста тканей для привлечения новых клеток и их последующего развития.
Наши данные предполагают, что, по крайней мере, три фактора могут объяснить, почему мыши C57 набирают больше жира, чем мыши FVB, при диете с высоким содержанием жиров: во-первых, мыши FVB имеют повышенную скорость метаболизма и повышенную ректальную температуру, скорее всего, из-за повышенного симпатического тонуса. Хотя мы не обнаружили значительных различий в активности между штаммами в нашем исследовании, более подробные измерения поведения показали, что мыши FVB спонтанно гиперактивны по сравнению с мышами C57 [26]. У них также увеличивается частота сердечных сокращений [36], и они реагируют гипергликемией на различные виды лечения [37].Кроме того, они более чувствительны к стрессу, связанному с воздержанием и голоданием [38]. Все эти данные, взятые вместе, предполагают, что активность симпатической нервной системы увеличилась больше у мышей FVB, чем активность у мышей C57. Во-вторых, по сравнению с мышами FVB, мыши C57 очищают циркулирующие триглицериды более эффективно [23], [24], что, по крайней мере, частично можно объяснить более высокой активностью липазы в сыворотке крови [23] и более высокой способностью накапливать триглицериды в печени [23]. ], [24] и жировой ткани (показано в этом исследовании).Хотя молекулярные механизмы клиренса триглицеридов до конца не изучены, жировая ткань явно способствует выведению триглицеридов, поскольку способность выводить циркулирующие триглицериды нарушена у липоатрофных мышей [23]. В частности, сообщалось, что диета с высоким содержанием жиров увеличивает клиренс триглицеридов [39], что может быть связано с индукцией активности липопротеинлипазы в жировой ткани [40]. Наконец, по сравнению с мышами FVB, мыши C57 показали большее количество мелких жировых клеток, особенно при диете с высоким содержанием жиров.Было высказано предположение, что новые жировые клетки могут возникать из клеток-предшественников, которые находятся в депо белого жира взрослых [41], [42], и из других источников, таких как циркулирующие клетки-предшественники костного мозга [43]. Было показано, что рекрутирование обоих типов предшественников стимулируется диетой с высоким содержанием жиров [41], [43]. Возможно, что большее рекрутирование более мелких жировых клеток у мышей C57 может быть вызвано более высоким пулом клеток-предшественников или их более высокой внутренней способностью к дифференцировке адипоцитов.Однако in vitro мезенхимальных стволовых клеток , выделенных из наружного уха мышей FVB и C57, одинаково хорошо дифференцируются в жировые клетки [44]. Попытка сравнить дифференцировку стромальных клеток костного мозга мышей FVB и C57 в жировые клетки не увенчалась успехом из-за очень низкого выхода и плохой пролиферативной способности клеток, выделенных от мышей C57 [45]; однако костный мозг, по-видимому, не является основным источником новых жировых клеток, по крайней мере, у мышей [43]. Наша модель предполагает, что разница между генотипами в рекрутировании малых жировых клеток может быть автономной по отношению к жировым подушечкам, но молекулярный механизм, лежащий в основе этого различия, неясен.Жировая подушечка представляет собой сложный орган, содержащий множество различных типов клеток, включая зрелые жировые, предадипозные и сосудистые клетки, нервы, макрофаги и фибробласты. Количество предшественников адипоцитов и их пролиферация в ответ на внешние сигналы варьируются в зависимости от жировых отложений [35]. Необходимы дальнейшие исследования, чтобы определить, как специфичное для генотипа взаимодействие между разными типами клеток и секретируемыми факторами может влиять на скорость рекрутирования адипоцитов в определенные жировые депо.
Таким образом, мы получили математическую модель, описывающую рост жировой ткани с увеличением количества и размера клеток в зависимости от массы жировой подушечки придатка яичка.Основываясь на этой динамической модели, мы исследовали влияние генетики и диеты на рост жировой ткани. Сравнивая распределение размеров клеток двух штаммов и двух диет, мы пришли к выводу, что изменение размера клеток зависит от диеты, а изменение количества клеток зависит от генетики и диеты, а также от их взаимодействия.
Материалы и методы
Животные
Все процедуры были одобрены Комитетом по уходу и использованию животных Национального института диабета, болезней органов пищеварения и почек.Самцы мышей FVB и C57 были получены из лаборатории Джексона (Бар-Харбор, штат Мэн). Мышей выращивали по четыре в клетке с 12-часовым циклом свет / темнота (свет 06: 00–18: 00). В возрасте 5 недель мышей каждой линии разделили на 2 группы. Половине мышей давали обычную пищу NIH-07 (далее REG; Zeigler Brothers, Inc., Gardners, PA), содержащую 4,08 ккал / г (11% калорий из жира, 62% из углеводов и 26% из белка). Другая половина получала диету с высоким содержанием жиров, F3282 (далее HF; Bio-Serv, Frenchtown, NJ), содержащую 5.45 ккал / г (59% жиров, 26% углеводов и 15% белков). Вода и диета были предоставлены ad libitum . Было проведено пять независимых экспериментов, в каждой из которых использовалось 4 группы мышей: FVB REG, FVB HF, C57 REG и C57 HF. В трех экспериментах мышей содержали на контролируемой диете в течение 12 недель и использовали для анализа состава тела, физиологической характеристики и распределения клеток по размеру. Две дополнительные группы мышей были умерщвлены после 2 недель и 4 недель контрольного кормления с высоким содержанием жира только для определения распределения клеток по размеру.
Эксперименты in vivo
Состав тела, потребление пищи, скорость метаболизма, толерантность к глюкозе, клиренс триглицеридов и окисление жирных кислот в изолированной камбаловидной мышце измеряли, как описано ранее [46]. Окисление жирных кислот всего тела измеряли, как описано в Gautam et al. [47]. Кровь для биохимических анализов брали из хвостовой вены без голодания. Уровни глюкозы измеряли с помощью Glucometer Elite (Bayer, Elkhart, IN). Сывороточный инсулин анализировали с помощью радиоиммуноанализа (Linco Research, St.Чарльз, Миссури). Триглицериды сыворотки, холестерин (Thermo DMA, Луисвилл, Колорадо) и свободная жирная кислота (FFA) (Roche Applied Science, Индианаполис, Индиана) измеряли в соответствии с процедурами производителя.
Измерение размера клеток эпидидимального жира
Распределение размеров клеток эпидидимального жира измеряли через 2, 4 и 12 недель высокожирового и контрольного кормления с использованием Beckman Coulter Multisizer III, как описано ранее [9]. Вкратце, 20–30 мг жировой ткани были взяты из средней части путем рассечения и последующего удаления образца для фиксации из центра разрезанного эпидидимального жира.Образцы тканей немедленно фиксировали в четырехокиси осмия [48], инкубировали на водяной бане при 37 ° C в течение 48 ч, а затем определяли размер жировых клеток на приборе Beckman Coulter Multisizer III с апертурой 400 µ м. Диапазон размеров ячеек, которые можно эффективно измерить с помощью этой апертуры, составляет 20–240 µ м. Прибор был настроен на подсчет 6000 частиц, и суспензия фиксированных клеток была разбавлена так, чтобы совпадающий счет был <10%. После сбора размеров импульсов данные были выражены в виде диаметров частиц и отображены в виде гистограмм отсчетов по диаметру с использованием линейных интервалов и линейной шкалы для оси x ().Распределение клеток по размерам было измерено в четырех образцах из каждой группы, за исключением мышей C57 после 4-недельного воздействия диеты с высоким содержанием жиров, у которых было только три доступных образца. Образец был взят из каждой жировой подушечки и обработан отдельно. Затем каждый образец был посчитан как минимум дважды. Затем кривые для двух образцов усредняются, но только после проверки воспроизводимости между двумя образцами.
Модель роста жировой ткани
Распределение размеров клеток включает всю информацию, относящуюся к размерам клеток в ткани, и его изменения дают статистическое представление о детальном процессе роста каждой клетки.Чтобы изучить рост жировой ткани с точки зрения лежащих в основе микроскопических процессов, мы рассматриваем математическую модель, количественно определяющую процессы, которые изменяют распределение клеток по размерам. Модель может предсказать, сколько новых клеток образуется и как клетки разного размера растут по мере увеличения массы жировой ткани. Плотность количества клеток определенного размера (диаметра) при данной массе жировой подушечки — это конкретное количество, которое необходимо учитывать. Мы рассматриваем, как это распределение клеток по размерам изменяется с постепенным изменением массы жировой ткани.Эволюция распределения клеток по размерам с массой жировой ткани может быть смоделирована уравнением в частных производных:
(1)
Это уравнение включает три основных компонента процесса роста жировой ткани. Во-первых, мы предполагаем, что рекрутирование новых клеток происходит только при минимальном наблюдаемом размере клетки, который математически выражается как дельта-функция. Скорость набора по отношению к массе жировой подушечки определяется экспоненциальной функцией:
(2)
, где — исходное общее количество клеток при данной начальной массе жировой подушечки, а — скорость увеличения количества клеток на единицу изменения. в массе жировой подушечки.Изменение общего числа клеток — это скорость пополнения новых клеток, если гибель клеток незначительна; мы не обнаружили необходимости включать апоптоз в клетки любого размера, чтобы соответствовать этим экспериментальным данным. Следовательно, этот коэффициент пополнения может быть непосредственно получен из экспериментального результата, используя соотношение между общим числом клеток и массой жировой подушечки путем дифференцирования функции по отношению к. Во-вторых, рост клеток зависит от размера клетки. После созревания жировых клеток до определенного размера они могут накапливать жир, вызывая гипертрофию.В дополнение к этой ограничивающей скорости роста маленьких жировых клеток, может также существовать верхний предел роста, потому что большие жировые клетки не могут расти бесконечно в результате этого процесса роста, хотя они могут достигать больших размеров из-за колебаний размера, вызванных обменом липидов. Повышение и снижение скорости роста клеток в зависимости от размера клетки можно описать с помощью общей функциональной формы, умножающей две сигмоидальные функции,
(3)
, где представляет максимальную скорость роста; и — соответственно нижний и верхний критические размеры, которые дают половину максимальной скорости роста; и укажите их масштаб ().Наконец, последний член в формуле. (1) представляет колебания размера клеток с постоянной скоростью, которые отражают обмен липидов, случайным образом происходящий в жировых клетках. Этот липидный оборот является единственным механизмом роста для крупных клеток, размер которых превышает верхний критический размер. Как правило, рост клеток, зависящий от размера, приводит к увеличению их распределения по размерам, в то время как колебания размеров распространяют распределение по размерам.
Метод оптимизации
Чтобы оптимизировать параметры модели, чтобы они могли точно описывать эволюцию распределения размеров ячеек в эксперименте, мы использовали минимизацию «функции стоимости», которая количественно определяет отклонение между моделью и результатами эксперимента.Чтобы определить функцию стоимости, нормализованное распределение размеров клеток при заданной массе жира сравнивалось с данными моделирования с набором параметров:
(6)
, где — общее количество интервалов размера клеток, а — общее количество заданных масса жира. Масштаб функции стоимости был рассчитан на основе внутренней флуктуации экспериментальных данных, которую можно определить как квадрат отклонения между измеренным распределением ячеек по размерам и функцией плавной аппроксимации:
(7)
Эта внутренняя флуктуация численно составляет около 10 процентов квадрата отклонения между экспериментальными и модельными данными.В качестве функции аппроксимации мы использовали сумму двух экспонент и гауссову форму,
(8)
, форму, которая использовалась для аппроксимации распределений размеров жировых клеток [9]. Подбор этих параметров был выполнен с использованием процедуры подбора нелинейной кривой в MATLAB R2007a (Натик, Массачусетс, США). Для процесса оптимизации мы специально использовали метод Монте-Карло с параллельным темперированием, чтобы найти глобальный минимум функции стоимости [50]. Мы использовали 10 равномерно распределенных значений (от 0,1 до 1) для параметра темперирования и запускали десять цепочек параллельно с вероятностью обновления.На каждых 20 шагах случайным образом выбиралась пара смежных имитаций по десяти параметрам отпуска, и их состояния параметров менялись с вероятностью. После достижения равновесия было использовано 20 000 итераций с фиксированным параметром отпуска для оценки оптимальных значений параметров и их стандартных ошибок.
Мы также использовали этот метод для оценки исходного общего количества клеток, и скорости его увеличения, исходя из соотношения между жировой массой и общим количеством клеток (). При минимизации функции аппроксимации и экспериментальных данных мы использовали ограничение, согласно которому начальное количество клеток одинаково независимо от условий диеты, т.е.е., обычные и жирные диеты. Средняя жировая масса четырех контрольных мышей перед регулярной диетой и диетой с высоким содержанием жиров была использована в качестве начальной жировой массы, которая составляет 0,34 г и 0,29 г для мышей FVB и C57, соответственно. Мы оценили погрешности и, распространив 10-процентные статистические флуктуации, наблюдаемые в экспериментальных данных.
Численное решение дифференциального уравнения с частными производными
Мы решили следующую дискретную версию нашей модели, заданную как непрерывное дифференциальное уравнение с частными производными в формуле.(1):
(9)
с интервалом масс δm = 0,1 мг и интервалом размеров δs = 0,73 µ м.
Динамика роста жировой ткани
Abstract
Рост жировой ткани происходит по двум механизмам: гиперплазия (увеличение количества клеток) и гипертрофия (увеличение размера клеток). Генетика и диета влияют на относительный вклад этих двух механизмов в рост жировой ткани при ожирении. В этом исследовании распределение по размеру эпидидимальных жировых клеток от двух линий мышей, устойчивых к ожирению FVB / N и склонных к ожирению C57BL / 6, измеряли через 2, 4 и 12 недель в условиях регулярного кормления и питания с высоким содержанием жира.Общее количество клеток в жировой подушке придатка яичка оценивали по массе жировой подушечки и нормализованному распределению клеток по размеру. Число клеток и средневзвешенный размер клеток увеличиваются в зависимости от массы жировой прослойки. Чтобы точно решить проблему роста жировой ткани, мы разработали математическую модель, описывающую эволюцию распределения размеров жировых клеток как функцию увеличения массы жировой ткани, а не увеличения хронологического времени. Наша модель описывает привлечение новых жировых клеток и их последующее развитие в разных штаммах и с разными режимами питания, с общими механизмами, но с параметрами модели, зависящими от диеты и генетики.По сравнению со штаммом FVB / N, штамм C57BL / 6 имеет большее количество мелких жировых клеток. Гиперплазия усиливается за счет диеты с высоким содержанием жиров в зависимости от штамма, что предполагает синергетическое взаимодействие между генетикой и диетой. Кроме того, кормление с высоким содержанием жиров увеличивает скорость роста жировых клеток, независимо от напряжения, что отражает увеличение количества калорий, требующих хранения. Кроме того, диета с высоким содержанием жиров приводит к резкому увеличению распределения жировых клеток по размерам у обоих штаммов; это означает увеличение колебаний размера жировых клеток за счет липидного обмена.
Авторское резюме
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. Динамическое изменение жировой ткани in vivo невозможно отслеживать в режиме реального времени из-за текущих технических ограничений. Однако мы можем измерить распределение жировых клеток по размеру у отдельных животных. Наша основная цель — извлечь динамические характеристики ремоделирования тканей из снимков распределения клеток по размерам.Мы разрабатываем математическую модель, которая интерполирует измерения распределения клеток по размеру и предсказывает непрерывное изменение распределения клеток по размеру в зависимости от увеличения массы жировых подушечек. Наша модель роста жировой ткани включает три основных компонента: рекрутирование новых клеток, рост клеток в зависимости от размера и колебания размера клеток. В частности, мы сравнили рост жировой ткани у склонных к ожирению и устойчивых к ожирению мышей при стандартной диете или диете с высоким содержанием жиров, чтобы изучить генетическое и диетическое влияние на рост жировой ткани.Применяя нашу модель к этим различным условиям, мы обнаружили, что увеличение размера жировых клеток зависит от диеты. С другой стороны, увеличение количества жировых клеток, вызванное диетой, зависит от деформации, что предполагает синергию между генетикой и диетой.
Введение
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. На фенотип ожирения жировой ткани влияют диета и генетика, а также их взаимодействие [1] — [4].Начиная с исследований Джонсона и Хирша [5], существует обширная литература о росте жировой ткани при нормальном и патологическом развитии, характеризующая состояние ткани с точки зрения среднего размера и количества клеток. Гиперпластический рост появляется только на ранних стадиях развития жировой ткани [6], [7]. Гипертрофия возникает до гиперплазии, чтобы удовлетворить потребность в дополнительных накоплениях жира при прогрессировании ожирения [8]. Однако оказалось трудно понять, как диета и генетика конкретно влияют на гиперплазию и / или гипертрофию жировых клеток из-за ограниченных данных о росте жировой ткани.
Помимо этих исследований, недавно стало возможным точно измерить распределение клеток по размерам. Эта подробная информация, по сравнению со средним размером клеток и общим числом клеток, может использоваться для вычисления многих величин, связанных с размером, которые позволяют более точно охарактеризовать процесс роста жировой ткани. Кумулянты распределения клеток по размерам могут использоваться для вычисления других физиологических величин, таких как средневзвешенный размер клеток. Распределение клеток по размерам можно использовать для оценки общего количества клеток в жировой подушке по ее массе.Более того, считается, что некоторые специфические метаболические свойства, например, резистентность к инсулину [9] и секреция адипокина [10], зависят от точного распределения клеток по размерам, а не от среднего размера клеток. Действительно, в нескольких исследованиях изучается изменение распределения жировых клеток по размерам в различных условиях при развитии куриного эмбриона [11], у худых и страдающих ожирением крыс Zucker [12], [13], крыс линии Wistar с частичной липэктомией [14], биопсии кролика [ 15] и жировой ткани человека [16], [17]. Эти исследования были сосредоточены только на статических различиях между распределениями клеток по размерам в разных условиях.Однако статические распределения размеров клеток в поперечном сечении для ряда снимков развития животных можно использовать для вывода динамики роста жировой ткани, если мы сможем надлежащим образом проанализировать снимки с помощью математического моделирования. Учитывая нынешние технические ограничения, это может быть лучший доступный подход к микроскопическому и продольному пониманию роста жировой ткани in vivo , хотя в недавнем эксперименте были получены микроскопические наблюдения накопления липидов в липидных каплях жировых клеток [18].
Для изучения генетического и диетического влияния на динамический процесс роста жировой ткани мы получили распределения по размерам клеток эпидидимального жира у устойчивых к ожирению линий мышей FVB / N (далее FVB) и склонных к ожирению линий мышей C57BL / 6 (C57) в соответствии с стандартные диеты и диеты с высоким содержанием жиров. Мышь C57 является наиболее охарактеризованной моделью ожирения, вызванного диетой [19], а мышь FVB является предпочтительной моделью для создания трансгенных мышей [20]. Эти два широко используемых инбредных штамма мышей генетически довольно далеки [21], [22] и имеют различные метаболические фенотипы: по сравнению с мышами FVB, мыши C57 имеют низкие уровни циркулирующих триглицеридов [21] и повышенный клиренс триглицеридов [23], [24]; Мыши FVB характеризуются относительно более высокой печеночной инсулинорезистентностью, контррегуляторной реакцией на гипогликемию и сниженной глюкозо-стимулированной секрецией инсулина [25]; Также известно, что мыши FVB являются спонтанно гиперактивными [26] и относительно худыми, поскольку они, по-видимому, менее восприимчивы к диете с высоким содержанием жиров, чем мыши C57 [27].Однако развитие ожирения, вызванного диетой, у этих двух штаммов формально не сравнивалось. В этом исследовании мы разработали математическую модель, прогнозирующую изменение распределения клеток по размерам в зависимости от массы жировой подушечки придатка яичка, для количественного анализа динамических характеристик, зависящих от генетики и / или диеты. Модель роста жировой ткани описывает, сколько новых клеток образуется, как каждая клетка растет в зависимости от ее размера и как липидный обмен приводит к колебаниям размера, которые вызывают расширение распределения клеток по размерам.По мере увеличения массы эпидидимальных жировых подушек распределение клеток по размерам систематически изменяется в зависимости как от генетики, так и от диеты. Сравнивая экспериментальные результаты с теоретической моделью роста, мы обнаружили, что гипертрофия сильно коррелирует с диетой. С другой стороны, гиперплазия зависит от генетики. Изменения при гиперплазии, вызванные диетой, зависят от штамма, что предполагает взаимодействие между диетой и генетикой.
Результаты
Влияние диеты с высоким содержанием жиров на состав тела мышей FVB и C57
В начале эксперимента (в возрасте 5 недель) мыши C57 были значительно легче, чем мыши FVB () из-за разницы в худой масса, хотя общая жировая масса не отличалась ().Когда мышей поддерживали на обычном питании, разница в массе тела исчезла к возрасту 11 недель (6-я неделя эксперимента). В условиях обычной диеты мыши FVB и C57 сохраняли сопоставимую массу жира на протяжении всего эксперимента (). Диета с высоким содержанием жиров вызвала значительное увеличение массы тела и жировой массы у обоих штаммов; однако изменения массы тела и жировой массы были более значительными у мышей C57. У мышей C57 было вдвое больше жира после 12 недель кормления с высоким содержанием жиров ().Общая разница в общей массе жира между мышами FVB и C57 коррелировала с пропорциональными различиями в количестве эпидидимального (внутрибрюшного), пахового (подкожного) и бурого жира (). Потребление калорий и активность были сопоставимы у мышей FVB HF и C57 HF; однако мыши FVB HF имели более высокий уровень покоя и общего потребления кислорода, а также более высокую ректальную температуру, что позволяет предположить, что увеличение расхода энергии, а не снижение потребления калорий, было причиной относительной устойчивости к ожирению, вызванному диетой с высоким содержанием жиров, у мышей FVB.Интересно, что в течение первых 2 недель кормления с высоким содержанием жиров мыши FVB и C57 показали сопоставимое увеличение общей жировой массы (). Мыши C57 HF продолжали быстро увеличивать жировую массу до 10 недели эксперимента, тогда как мыши FVB HF замедляли накопление жира около недели 3. У мышей C57 кормление с высоким содержанием жира вызывало постепенное увеличение как эпидидимальных, так и паховых жировых подушечек; Напротив, у мышей FVB жировая масса придатка яичка увеличилась лишь незначительно после 4 недель кормления с высоким содержанием жира, в то время как паховая жировая подушечка продолжала увеличиваться в размере на протяжении всего эксперимента (рис.S1). Кормление с высоким содержанием жиров вызывало значительное повышение уровня глюкозы и инсулина в крови как у мышей FVB, так и у мышей C57 (). Уровни инсулина и непереносимость глюкозы были выше у мышей C57 HF, чем у мышей FVB HF, что свидетельствует о более тяжелой инсулинорезистентности (). В соответствии с предыдущими сообщениями [23], [24], мыши C57 REG показали пониженные уровни триглицеридов в сыворотке по сравнению с мышами FVB REG без разницы в FFA (). Это происходило не из-за более высокой утилизации жира, так как коэффициент респираторного обмена () и скорость окисления жирных кислот, измеренная in vivo, () и в изолированных скелетных мышцах () были сопоставимы у мышей FVB REG и C57 REG.Более вероятно, что более низкие уровни триглицеридов в сыворотке у мышей C57 REG были вызваны гораздо более эффективным клиренсом циркулирующих триглицеридов, как предполагает тест на клиренс триглицеридов (). Кормление с высоким содержанием жиров снижало уровни циркулирующих триглицеридов как у мышей FVB, так и у мышей C57 и улучшало клиренс триглицеридов у последних штаммов (и). Оба штамма показали сопоставимое снижение коэффициента респираторного обмена, что свидетельствует о сопоставимом увеличении утилизации жирных кислот в условиях диеты с высоким содержанием жиров (). Взятые вместе, эти данные предполагают, что способность эффективно выводить триглицериды из кровотока может способствовать высокой способности накопления жира у мышей C57.
Изменения в составе тела мышей FVB и C57 в условиях стандартного корма (REG) и с высоким содержанием жира (HF).(A) Масса тела. (B) Жировая масса. Состав тела измеряли в указанные моменты времени у 8 мышей в группе с использованием МРТ-анализатора Echo 3-in-1. Высокожировое и контрольное кормление начали в возрасте 5 недель. Ось X показывает недели контролируемого кормления. Показан один из трех независимых экспериментов. Приведенные значения являются средними ± SEM.
Физиологические характеристики мышей FVB и C57.(A) Тест на толерантность к глюкозе был проведен на 10-й неделе контролируемого кормления с пищей и диетой с высоким содержанием жиров. Мышей не кормили в течение ночи и вводили глюкозу (2 мг / г, внутрибрюшинно). Каждая группа была представлена 8 мышами. (B) Окисление олеиновой кислоты в целом организме измеряли у самцов мышей FVB ad C57 в возрасте 10 недель в не голодном состоянии (8 мышей на группу), как описано в Gautam et al. [47]. (C) Окисление пальмитиновой кислоты измеряли в камбаловидной мышце, взятой у 10-недельных мышей FVB и C57, которых содержали на диете (8 мышей на группу).Образцы собирали в 8 часов утра у мышей, которых случайным образом кормили, или мышей, голодавших в течение 18 часов [методы, описанные в Toyoshima et al. [46]]. (D) Клиренс триглицеридов у самцов мышей FVB и C57 после 11 недель контролируемого кормления кормом и диетой с высоким содержанием жиров. Мышей не кормили в течение 4 часов, а затем вводили через желудочный зонд 400 мкл л арахисового масла. Плазму собирали ежечасно в течение 5 часов из хвостовой вены для измерения триглицеридов. Каждая группа была представлена 8 мышами.
Таблица 1
Характеристики мышей FVB и C57 после 12 недель высокожирового и контрольного кормления.
| FVB REG a | FVB HF a | C57 REG a | C57 HF | a 94 996| 912 912 912 912 912 Взаимодействие б | | ||||||||||
| Масса тела (г) | 29,1 ± 0,4 | 35,4 ± 1,4 | 29,1 ± 0,4 | 43,0 ± 1,8 | 0,003 | <0,001 | 0,003 | г)0.47 ± 0,03 | 1,23 ± 0,11 | 0,67 ± 0,08 | 2,00 ± 0,21 | <0,001 | <0,001 | 0,032 | |
| Паховой жир (г) | 0,3412 ± 0,02 | 0,190,3412 ± 0,02 | 0,3412 ± 0,02 | 0,45 ± 0,041,92 ± 0,27 | 0,003 | <0,001 | <0,003 | ||||||||
| Коричневый жир (г) | 0,14 ± 0,01 | 0,27 ± 0,04 | 0,1412 ± 0,01 | 0,01 0,09 0,003<0.001 | 0,003 | ||||||||||
| Потребление пищи (ккал / мышь / день) | 16,6 ± 1,0 | 18,0 ± 1,3 | 13,2 ± 0,8 | 17,7 ± 0,8 | 0,07 | 0,007 92| 3 ns Общее потребление кислорода (мл / г) | 9,5 ± 0,2 | 11,6 ± 0,3 | 9,0 ± 0,2 | 9,4 ± 0,2 | <0,001 | <0,001 | нс | | |
| Потребление кислорода в покое (мл / г) ) | 8.3 ± 0,2 | 9,7 ± 0,3 | 7,3 ± 0,2 | 8,1 ± 0,3 | <0,001 | <0,001 | 0,003 | ||||||||
| Коэффициент респираторного обмена | 0,86 ± 0,01 | 0,75 ± 0,01 0,01 | 0,72 ± 0,00 | нс | <0,001 | 0,006 | |||||||||
| Суммарная активность (торможение луча / мин) | 265 ± 5 | 190 ± 17 | 345 ± 41 | 122011293 нс | <0.001 | нс | |||||||||
| Ходьба (торможение луча / мин) | 105 ± 3 | 53 ± 10 | 144 ± 25 | 38 ± 5 | нс | <0,002 | 91 Ректальная температура (° C) | 36,0 ± 0,2 | 36,6 ± 0,2 | 35,4 ± 0,2 | 35,5 ± 0,2 | <0,001 | 0,057 | нс | |
| Триглицерид сыворотки (мг / дл) | ± 25 | 95 ± 24 | 89 ± 16 | 64 ± 6 | 0.002 | 0,004 | ns | ||||||||
| Свободные жирные кислоты (мМ) | 0,22 ± 0,03 | 0,22 ± 0,01 | 0,26 ± 0,04 | 0,27 ± 0,05 | 1292 ns | ns | ns Глюкоза крови (мг / дл) | 187 ± 16 | 225 ± 11 | 209 ± 11 | 247 ± 15 | нс | 0,009 | нс | |
| Инсулин сыворотки (мг / | 1,1) 912 ± 0,1 | 1,9 ± 0,3 | 2.2 ± 0,8 | 4,9 ± 1,2 | 0,011 | 0,033 | нс | ||||||||
| Клиренс триглицеридов (AUG c ) | 3238 ± 476 | 1651 ± 160 | 48 | 489 | <0,001 | 0,011 | <0,001 | ||||||||
| Толерантность к глюкозе (AUG d ) | 28633 ± 1372 | 39560 ± 3217 | 25947 ± 1212 | 001 | 0,003 |
Гипертрофия и гиперплазия с увеличением жировой массы
Чтобы проверить основной механизм различной скорости накопления жира в эпидидимальном жире мышей FVB и C57, мы измерили распределение массы и размеров клеток в образцах тканей эпидидимальный жир, собранный на 0, 2, 4 и 12 неделях контролируемого кормления (). Поскольку гистологический анализ не позволяет точно определить размер клеток адипоцитов, который будет обсуждаться позже, мы измерили распределение клеток по размеру с помощью счетчика Коултера и рассчитали средневзвешенный размер клеток и общее количество клеток эпидидимальной жировой подушки по этим измерениям, которые аналогичные значения в другом исследовании на мышах [5].Сильная корреляция наблюдалась между массой жировых подушечек и средневзвешенным размером клеток, а также между массой жировых подушечек и общим числом клеток, независимо от напряжения и разницы в диете (). Первая корреляция дала масштабное соотношение между массой жировой подушечки и средневзвешенным по объему размером клеток (). Кроме того, была обнаружена экспоненциальная зависимость между массой жировой подушечки, и общим числом клеток, где начальная масса жировой подушки была получена от контрольных мышей; и начальное количество клеток, и скорость увеличения количества клеток в массе жировой подушечки, были оценены по данным (;).Первоначальное количество клеток у мышей C57 было больше, чем исходное количество клеток у мышей FVB (фиг. S2C и S2D). По мере увеличения массы жировой ткани увеличивается общее количество клеток. Скорость увеличения числа клеток была выше при обычном питании, чем при диете с высоким содержанием жиров, и эта тенденция более очевидна у мышей C57 (;), что указывает на генетическое различие. Соотношение между результатами регулярной диеты и диеты с высоким содержанием жиров составляет 1,42 и 3,22 для мышей FVB и C57 соответственно (). Это может указывать на взаимодействие между генетикой и диетой при увеличении количества клеток.Обратите внимание, что мы также наблюдали аналогичные результаты с массой тела и массой жира, поскольку три величины (масса жировых подушечек придатка яичка, масса жира и масса тела) коррелируют друг с другом. Однако результаты с массой эпидидимальной жировой подушечки были наиболее подходящими: среднеквадратичное отклонение между данными и соответствием составило 9,73, 7,94, 3,56 × 10 5 и 5,54 × 10 6 , соответственно; результат с массой тела составил 11,82, 15,35, 3,65 × 10 5 и 4,82 × 10 6 ; результат с жировой массой 8.58, 8,61, 4,28 × 10 5 и 6,11 × 10 6 .
Нормализованное распределение клеток по размерам в жировой подушке придатка яичка.На графике нанесен один типичный пример контрольных мышей FVB в возрасте пяти недель. Кружки обозначают точки измеренных данных, а линия представляет собой аппроксимирующую кривую, которая представляет собой сумму одной гауссовой и двух экспоненциальных функций.
Таблица 2
Значения параметров модели роста жировой ткани.
Генетические и диетические эффекты на рост жировой ткани
Эти две сильные корреляции между массой жировой подушечки и гипертрофией и между массой жировой подушечки и гиперплазией позволяют предположить, что увеличение жировой ткани можно описать как систематический процесс роста в отношении для увеличения массы жировой прослойки.Мы расположили распределения клеток по размерам, отсортированные по массе эпидидимальных жировых подушечек (). Примечательно, что модель роста жировой ткани в формуле. (1) описывает эволюционирующую картину распределения клеток по размерам в зависимости от увеличения массы жировой прослойки. Модель количественно соответствовала экспериментальным распределениям размеров клеток, несмотря на то, что все распределения представляют собой данные поперечного сечения, полученные от отдельных животных. Различные значения параметров в модели, которые соответствуют каждому индивидуальному распределению размеров клеток для обоих штаммов и обоих режимов питания, дали количественные различия в процессе роста эпидидимальной жировой ткани между штаммами и между диетами ().Во-первых, максимальная зависящая от размера скорость роста и скорость колебаний размера клеток из-за обмена липидов продемонстрировали разницу, вызванную диетой, и меньшую разницу, вызванную деформацией. Зависимый от размера рост и колебания размера, приводящие к гипертрофии, по-видимому, регулируются в основном диетой. В частности, рост, зависящий от размера, сдвинул режим больших клеток в распределении размеров клеток в сторону больших размеров, а флуктуации липидного обмена увеличили разброс распределения вокруг режима больших клеток.Важно отметить, что результаты должны быть тщательно интерпретированы, потому что каждая скорость — это скорость на единицу увеличения массы жировой ткани, а не на единицу увеличения времени. Во-вторых, геометрические параметры (, и), которые определяют форму скорости роста, зависящей от размера, имели практически одинаковые значения независимо от диеты и разницы деформаций, за исключением. Следовательно, нижний критический размер, который дает размер, инициирующий рост клеток в зависимости от размера, и два параметра масштаба могут быть зафиксированы на с l = 37 µ m, η l = 12 µ м и η u = 63 µ м соответственно.С другой стороны, верхний критический размер, ограничивающий зависимый от размера клетки рост больших клеток, зависел от диеты; при кормлении с высоким содержанием жира этот пороговый размер сместился в сторону большего размера (). При диете с высоким содержанием жиров изменения параметров (,, и) увеличивают способность жировых тканей накапливать липиды за счет как гиперплазии, так и гипертрофии. Более низкие уровни триглицеридов в сыворотке при условии диеты с высоким содержанием жиров (и) могут коррелировать с увеличением накопления липидов в увеличенных жировых клетках, потому что не было обнаружено значительных различий в окислении жирных кислот, о чем свидетельствует отсутствие различий в соотношении респираторного обмена ().
Изменения распределения клеток по размерам с увеличением массы эпидидимальной жировой подушечки при кормлении и диетах с высоким содержанием жиров.Начальные распределения размеров клеток пятинедельных контрольных мышей приведены в первой строке каждого столбца пунктирными линиями. Точки показывают экспериментальные результаты. Обратите внимание, что результаты упорядочены по массе эпидидимальной жировой подушечки, а не по хронологическому времени (недели контролируемого кормления), указанному в скобках. Сплошные линии представляют собой нормализованные распределения клеток по размеру, соответствующие данной массе эпидидимальной жировой подушечки, предсказанной моделью роста жировой ткани.
Зависимая от размера скорость роста массы придатка яичка.Здесь для этого графика используются средние значения параметров. Символы показаны для облегчения сравнения строк.
Обсуждение
Наше центральное открытие состоит в том, что гиперплазия и гипертрофия жировых клеток в эпидидимальной жировой подушке является функцией массы жировой подушечки, даже несмотря на то, что у отдельных животных может потребоваться разное время для достижения заданной массы жировой подушечки. Следовательно, рост жировой ткани, представленный как изменения распределения клеток по размерам, можно систематически моделировать как процесс роста по отношению к увеличению массы жировой подушечки; это может отражать корреляцию между массой жировой подушечки и секрецией адипокинов и других сигнальных молекул, контролирующих рост жировой ткани.Соответственно, следует отметить, что нормы в нашей модели — это не обычные нормы увеличения на единицу времени, а нормы увеличения на единицу массы. Таким образом, несколько показателей (, и) в модели имели более высокие значения для животных, соблюдающих диету, чем для животных, соблюдающих диету с высоким содержанием жиров. Однако, если бы эти показатели были преобразованы в обычные нормы увеличения на единицу времени, они имели бы большие значения для диеты с высоким содержанием жиров, потому что требуется меньше времени для увеличения массы жировой подушечки на единицу из более крупных и более многочисленных клеток на диета с высоким содержанием жиров, чем для увеличения такой же величины за счет меньшего и меньшего количества клеток на диете (рис.S1). Было высказано предположение, что когда ожирение прогрессирует, рано возникает гипертрофия жировых клеток, а затем запускается гиперплазия [8]. Наше исследование показало, что рекрутирование новых клеток экспоненциально увеличивается с увеличением массы жировой ткани. Гипертрофия жировых клеток является основным фактором увеличения массы жировой прослойки, тогда как гиперплазия не способствует этому увеличению, потому что она возникает в небольших клетках, в которых хранится гораздо меньший объем жира. Таким образом, наша модель естественным образом воплощает идею о том, что на гиперплазию влияет гипертрофический рост клеток.С другой стороны, сообщалось, что гиперплазия жировых клеток возникает только на ранних стадиях развития; следовательно, не следует ожидать рекрутирования новых клеток на поздних стадиях даже в условиях ожирения [6], [7]. Возможно, возраст животных в нашем исследовании (6 недель) допускает возникновение гиперплазии.
Модель, разработанная здесь, может дать микроскопическое представление о росте жировых клеток, зависящем от размера, который не может быть рассмотрен статическими поперечными исследованиями.Например, мы обнаружили следующие специфические свойства роста клеток, зависящие от размера: нижний критический размер, инициирующий накопление липидов, не зависел от диеты у двух линий мышей, тогда как верхний критический размер ограничивал рост клеток до достижения необходимого уровня. необычайный размер, был увеличен на диете с высоким содержанием жиров. Эта зависимость роста клеток от размера является проверяемой гипотезой. Далее, параметр флуктуации размера клеток различался при обычном питании и рационе с высоким содержанием жиров; оно больше при диете с высоким содержанием жиров, когда оно преобразуется в единицы, соответствующие изменению единицы времени, а не единице изменения массы жировой подушечки.Таким образом, случайный процесс высвобождения и поглощения жира жировыми клетками происходит более активно при диете с высоким содержанием жиров, чем при обычной диете. Может быть интересно посмотреть, можно ли распространить эти результаты на другие штаммы и организмы.
По сравнению с исследованиями, в которых наблюдали единственный пик в распределении размеров клеток жировых клеток [11], [15] — [17], [28], мы наблюдали бимодальные распределения клеток по размерам, как сообщалось другими [12], [13], [29] — [32]. В большинстве исследований [12], [13], [31], [32] по наблюдению за бимодальностью использовалась технология счетчика Коултера, которая имеет несколько преимуществ для оценки всего распределения размеров ячеек [31]: Во-первых, можно доказать, что анализируемые ячейки быть подлинными жировыми клетками на основе морфологии и флотации; во-вторых, объем каждой клетки оценивается независимо от формы и без артефактов смещения центра, что является правилом, а не исключением, с использованием гистологических подходов; наконец, можно подсчитать и измерить достаточное количество частиц, чтобы получить статистически значимые комплексные кривые.Напротив, микроскопические методы гистологии могут не наблюдать мелкие клетки из-за влияния увеличения микроскопа [30], небольшого количества образцов и систематической ошибки выборки. Однако при использовании счетчика Коултера контаминация, не связанная с адипоцитами, может способствовать распределению клеток по размерам, особенно при малых размерах, хотя наш минимальный диаметр клеток, 22 µ м, превышает возможные диапазоны контаминации, от 10 до 20 µ м, упомянутый Mersmann и MacNeil [31]. Чтобы быть уверенным, мы снова проанализировали модифицированные данные, используя только распределения размеров клеток выше 35 µ м в диаметре с моделью, и пришли к тем же выводам (данные не показаны).Надир в распределении размеров клеток () может разделять две популяции клеток. ДеМартини и Франсендез определили маленькие клетки с диаметром менее 35 µ м как «очень маленькие жировые клетки» [29]. Согласно нашей модели, эти клетки имеют незначительный рост в зависимости от размера, поскольку их размер меньше нижнего критического размера, с l = 37 µ m. Следовательно, механизм роста, зависящий от размера, может естественным образом объяснить происхождение бимодальности в распределении жировых клеток по размеру.Клетки с размером только выше могут расти в зависимости от размера, но клетки с размером меньше могут расти случайным образом с флуктуацией размера за счет липидного обмена. Это разделение вызывает накопление клеток ниже размера, что дает более низкий пик в распределении размеров клеток. Эта популяция клеток может служить потенциальным резервуаром для зрелых жировых клеток. Процесс их созревания можно интерпретировать следующим образом: жировые клетки, достигающие критического размера, вследствие случайных колебаний размера могут расти с механизмом роста, зависящим от размера.Как упоминалось выше, изменение размера происходит более активно при диете с высоким содержанием жиров; следовательно, пласт может ускорить процесс созревания в условиях стимуляции.
В модель роста ткани мы включили привлечение новых клеток и рост существующих клеток, но не гибель старых клеток, потому что модель соответствовала данным без апоптоза жировых клеток. Этот результат также согласуется с исследованием, в котором наблюдалось, что жировая ткань придатка яичка мышей C57BL / 6 не демонстрирует значительной гибели клеток до 12 недель при диете с высоким содержанием жиров [33].Однако расширенная диета с высоким содержанием жиров в конечном итоге вызывает апоптоз жировых клеток [33]. Более того, в одном недавнем исследовании сообщается, что жировые клетки человека обновляются в течение десяти лет [7]. Таким образом, наша модель все еще нуждается в улучшении, чтобы ее можно было использовать в различных условиях. Следует учитывать гибель клеток и формально учитывать зависимость параметров модели от диеты. В этом исследовании мы сосредоточились на одном жировом депо, эпидидимальном жире, по нескольким причинам: 1) вес эпидидимальных жировых подушечек может быть определен более точно, чем вес паховых жировых подушечек из-за легкости иссечения; 2) морфология жировых клеток в эпидидимальном жире более однородна, чем в паховом жире, который содержит много клеток, подобных коричневой жировой ткани, особенно у мышей, устойчивых к ожирению, вызванному диетой; и 3) разница между генотипами была более очевидной в росте эпидидимального жира, который достигает плато через 4 недели у мышей FVB, но продолжает расти у мышей C57 на протяжении всего эксперимента, в отличие от пахового жира, который показывает устойчивый рост. у обоих штаммов (рис.S1). Хотя мы не измеряли распределение по размеру клеток других жировых отложений, мы измерили изменение массы паховых и бурых жировых отложений, что показывает аналогичную картину с эпидидимальным жировым депо (рис. S1). Таким образом, представляет интерес применить модель к другим жировым отложениям, которые имеют функциональные различия [34], [35], а также к другим видам, таким как человек, что остается для будущих исследований. Мы ожидаем, что модель может быть применена к таким разнообразным наборам данных, просто настроив параметры модели, потому что модель содержит общие механизмы роста тканей для привлечения новых клеток и их последующего развития.
Наши данные предполагают, что, по крайней мере, три фактора могут объяснить, почему мыши C57 набирают больше жира, чем мыши FVB, при диете с высоким содержанием жиров: во-первых, мыши FVB имеют повышенную скорость метаболизма и повышенную ректальную температуру, скорее всего, из-за повышенного симпатического тонуса. Хотя мы не обнаружили значительных различий в активности между штаммами в нашем исследовании, более подробные измерения поведения показали, что мыши FVB спонтанно гиперактивны по сравнению с мышами C57 [26]. У них также увеличивается частота сердечных сокращений [36], и они реагируют гипергликемией на различные виды лечения [37].Кроме того, они более чувствительны к стрессу, связанному с воздержанием и голоданием [38]. Все эти данные, взятые вместе, предполагают, что активность симпатической нервной системы увеличилась больше у мышей FVB, чем активность у мышей C57. Во-вторых, по сравнению с мышами FVB, мыши C57 очищают циркулирующие триглицериды более эффективно [23], [24], что, по крайней мере, частично можно объяснить более высокой активностью липазы в сыворотке крови [23] и более высокой способностью накапливать триглицериды в печени [23]. ], [24] и жировой ткани (показано в этом исследовании).Хотя молекулярные механизмы клиренса триглицеридов до конца не изучены, жировая ткань явно способствует выведению триглицеридов, поскольку способность выводить циркулирующие триглицериды нарушена у липоатрофных мышей [23]. В частности, сообщалось, что диета с высоким содержанием жиров увеличивает клиренс триглицеридов [39], что может быть связано с индукцией активности липопротеинлипазы в жировой ткани [40]. Наконец, по сравнению с мышами FVB, мыши C57 показали большее количество мелких жировых клеток, особенно при диете с высоким содержанием жиров.Было высказано предположение, что новые жировые клетки могут возникать из клеток-предшественников, которые находятся в депо белого жира взрослых [41], [42], и из других источников, таких как циркулирующие клетки-предшественники костного мозга [43]. Было показано, что рекрутирование обоих типов предшественников стимулируется диетой с высоким содержанием жиров [41], [43]. Возможно, что большее рекрутирование более мелких жировых клеток у мышей C57 может быть вызвано более высоким пулом клеток-предшественников или их более высокой внутренней способностью к дифференцировке адипоцитов.Однако in vitro мезенхимальных стволовых клеток , выделенных из наружного уха мышей FVB и C57, одинаково хорошо дифференцируются в жировые клетки [44]. Попытка сравнить дифференцировку стромальных клеток костного мозга мышей FVB и C57 в жировые клетки не увенчалась успехом из-за очень низкого выхода и плохой пролиферативной способности клеток, выделенных от мышей C57 [45]; однако костный мозг, по-видимому, не является основным источником новых жировых клеток, по крайней мере, у мышей [43]. Наша модель предполагает, что разница между генотипами в рекрутировании малых жировых клеток может быть автономной по отношению к жировым подушечкам, но молекулярный механизм, лежащий в основе этого различия, неясен.Жировая подушечка представляет собой сложный орган, содержащий множество различных типов клеток, включая зрелые жировые, предадипозные и сосудистые клетки, нервы, макрофаги и фибробласты. Количество предшественников адипоцитов и их пролиферация в ответ на внешние сигналы варьируются в зависимости от жировых отложений [35]. Необходимы дальнейшие исследования, чтобы определить, как специфичное для генотипа взаимодействие между разными типами клеток и секретируемыми факторами может влиять на скорость рекрутирования адипоцитов в определенные жировые депо.
Таким образом, мы получили математическую модель, описывающую рост жировой ткани с увеличением количества и размера клеток в зависимости от массы жировой подушечки придатка яичка.Основываясь на этой динамической модели, мы исследовали влияние генетики и диеты на рост жировой ткани. Сравнивая распределение размеров клеток двух штаммов и двух диет, мы пришли к выводу, что изменение размера клеток зависит от диеты, а изменение количества клеток зависит от генетики и диеты, а также от их взаимодействия.
Материалы и методы
Животные
Все процедуры были одобрены Комитетом по уходу и использованию животных Национального института диабета, болезней органов пищеварения и почек.Самцы мышей FVB и C57 были получены из лаборатории Джексона (Бар-Харбор, штат Мэн). Мышей выращивали по четыре в клетке с 12-часовым циклом свет / темнота (свет 06: 00–18: 00). В возрасте 5 недель мышей каждой линии разделили на 2 группы. Половине мышей давали обычную пищу NIH-07 (далее REG; Zeigler Brothers, Inc., Gardners, PA), содержащую 4,08 ккал / г (11% калорий из жира, 62% из углеводов и 26% из белка). Другая половина получала диету с высоким содержанием жиров, F3282 (далее HF; Bio-Serv, Frenchtown, NJ), содержащую 5.45 ккал / г (59% жиров, 26% углеводов и 15% белков). Вода и диета были предоставлены ad libitum . Было проведено пять независимых экспериментов, в каждой из которых использовалось 4 группы мышей: FVB REG, FVB HF, C57 REG и C57 HF. В трех экспериментах мышей содержали на контролируемой диете в течение 12 недель и использовали для анализа состава тела, физиологической характеристики и распределения клеток по размеру. Две дополнительные группы мышей были умерщвлены после 2 недель и 4 недель контрольного кормления с высоким содержанием жира только для определения распределения клеток по размеру.
Эксперименты in vivo
Состав тела, потребление пищи, скорость метаболизма, толерантность к глюкозе, клиренс триглицеридов и окисление жирных кислот в изолированной камбаловидной мышце измеряли, как описано ранее [46]. Окисление жирных кислот всего тела измеряли, как описано в Gautam et al. [47]. Кровь для биохимических анализов брали из хвостовой вены без голодания. Уровни глюкозы измеряли с помощью Glucometer Elite (Bayer, Elkhart, IN). Сывороточный инсулин анализировали с помощью радиоиммуноанализа (Linco Research, St.Чарльз, Миссури). Триглицериды сыворотки, холестерин (Thermo DMA, Луисвилл, Колорадо) и свободная жирная кислота (FFA) (Roche Applied Science, Индианаполис, Индиана) измеряли в соответствии с процедурами производителя.
Измерение размера клеток эпидидимального жира
Распределение размеров клеток эпидидимального жира измеряли через 2, 4 и 12 недель высокожирового и контрольного кормления с использованием Beckman Coulter Multisizer III, как описано ранее [9]. Вкратце, 20–30 мг жировой ткани были взяты из средней части путем рассечения и последующего удаления образца для фиксации из центра разрезанного эпидидимального жира.Образцы тканей немедленно фиксировали в четырехокиси осмия [48], инкубировали на водяной бане при 37 ° C в течение 48 ч, а затем определяли размер жировых клеток на приборе Beckman Coulter Multisizer III с апертурой 400 µ м. Диапазон размеров ячеек, которые можно эффективно измерить с помощью этой апертуры, составляет 20–240 µ м. Прибор был настроен на подсчет 6000 частиц, и суспензия фиксированных клеток была разбавлена так, чтобы совпадающий счет был <10%. После сбора размеров импульсов данные были выражены в виде диаметров частиц и отображены в виде гистограмм отсчетов по диаметру с использованием линейных интервалов и линейной шкалы для оси x ().Распределение клеток по размерам было измерено в четырех образцах из каждой группы, за исключением мышей C57 после 4-недельного воздействия диеты с высоким содержанием жиров, у которых было только три доступных образца. Образец был взят из каждой жировой подушечки и обработан отдельно. Затем каждый образец был посчитан как минимум дважды. Затем кривые для двух образцов усредняются, но только после проверки воспроизводимости между двумя образцами.
Модель роста жировой ткани
Распределение размеров клеток включает всю информацию, относящуюся к размерам клеток в ткани, и его изменения дают статистическое представление о детальном процессе роста каждой клетки.Чтобы изучить рост жировой ткани с точки зрения лежащих в основе микроскопических процессов, мы рассматриваем математическую модель, количественно определяющую процессы, которые изменяют распределение клеток по размерам. Модель может предсказать, сколько новых клеток образуется и как клетки разного размера растут по мере увеличения массы жировой ткани. Плотность количества клеток определенного размера (диаметра) при данной массе жировой подушечки — это конкретное количество, которое необходимо учитывать. Мы рассматриваем, как это распределение клеток по размерам изменяется с постепенным изменением массы жировой ткани.Эволюция распределения клеток по размерам с массой жировой ткани может быть смоделирована уравнением в частных производных:
(1)
Это уравнение включает три основных компонента процесса роста жировой ткани. Во-первых, мы предполагаем, что рекрутирование новых клеток происходит только при минимальном наблюдаемом размере клетки, который математически выражается как дельта-функция. Скорость набора по отношению к массе жировой подушечки определяется экспоненциальной функцией:
(2)
, где — исходное общее количество клеток при данной начальной массе жировой подушечки, а — скорость увеличения количества клеток на единицу изменения. в массе жировой подушечки.Изменение общего числа клеток — это скорость пополнения новых клеток, если гибель клеток незначительна; мы не обнаружили необходимости включать апоптоз в клетки любого размера, чтобы соответствовать этим экспериментальным данным. Следовательно, этот коэффициент пополнения может быть непосредственно получен из экспериментального результата, используя соотношение между общим числом клеток и массой жировой подушечки путем дифференцирования функции по отношению к. Во-вторых, рост клеток зависит от размера клетки. После созревания жировых клеток до определенного размера они могут накапливать жир, вызывая гипертрофию.В дополнение к этой ограничивающей скорости роста маленьких жировых клеток, может также существовать верхний предел роста, потому что большие жировые клетки не могут расти бесконечно в результате этого процесса роста, хотя они могут достигать больших размеров из-за колебаний размера, вызванных обменом липидов. Повышение и снижение скорости роста клеток в зависимости от размера клетки можно описать с помощью общей функциональной формы, умножающей две сигмоидальные функции,
(3)
, где представляет максимальную скорость роста; и — соответственно нижний и верхний критические размеры, которые дают половину максимальной скорости роста; и укажите их масштаб ().Наконец, последний член в формуле. (1) представляет колебания размера клеток с постоянной скоростью, которые отражают обмен липидов, случайным образом происходящий в жировых клетках. Этот липидный оборот является единственным механизмом роста для крупных клеток, размер которых превышает верхний критический размер. Как правило, рост клеток, зависящий от размера, приводит к увеличению их распределения по размерам, в то время как колебания размеров распространяют распределение по размерам.
Метод оптимизации
Чтобы оптимизировать параметры модели, чтобы они могли точно описывать эволюцию распределения размеров ячеек в эксперименте, мы использовали минимизацию «функции стоимости», которая количественно определяет отклонение между моделью и результатами эксперимента.Чтобы определить функцию стоимости, нормализованное распределение размеров клеток при заданной массе жира сравнивалось с данными моделирования с набором параметров:
(6)
, где — общее количество интервалов размера клеток, а — общее количество заданных масса жира. Масштаб функции стоимости был рассчитан на основе внутренней флуктуации экспериментальных данных, которую можно определить как квадрат отклонения между измеренным распределением ячеек по размерам и функцией плавной аппроксимации:
(7)
Эта внутренняя флуктуация численно составляет около 10 процентов квадрата отклонения между экспериментальными и модельными данными.В качестве функции аппроксимации мы использовали сумму двух экспонент и гауссову форму,
(8)
, форму, которая использовалась для аппроксимации распределений размеров жировых клеток [9]. Подбор этих параметров был выполнен с использованием процедуры подбора нелинейной кривой в MATLAB R2007a (Натик, Массачусетс, США). Для процесса оптимизации мы специально использовали метод Монте-Карло с параллельным темперированием, чтобы найти глобальный минимум функции стоимости [50]. Мы использовали 10 равномерно распределенных значений (от 0,1 до 1) для параметра темперирования и запускали десять цепочек параллельно с вероятностью обновления.На каждых 20 шагах случайным образом выбиралась пара смежных имитаций по десяти параметрам отпуска, и их состояния параметров менялись с вероятностью. После достижения равновесия было использовано 20 000 итераций с фиксированным параметром отпуска для оценки оптимальных значений параметров и их стандартных ошибок.
Мы также использовали этот метод для оценки исходного общего количества клеток, и скорости его увеличения, исходя из соотношения между жировой массой и общим количеством клеток (). При минимизации функции аппроксимации и экспериментальных данных мы использовали ограничение, согласно которому начальное количество клеток одинаково независимо от условий диеты, т.е.е., обычные и жирные диеты. Средняя жировая масса четырех контрольных мышей перед регулярной диетой и диетой с высоким содержанием жиров была использована в качестве начальной жировой массы, которая составляет 0,34 г и 0,29 г для мышей FVB и C57, соответственно. Мы оценили погрешности и, распространив 10-процентные статистические флуктуации, наблюдаемые в экспериментальных данных.
Численное решение дифференциального уравнения с частными производными
Мы решили следующую дискретную версию нашей модели, заданную как непрерывное дифференциальное уравнение с частными производными в формуле.(1):
(9)
с интервалом масс δm = 0,1 мг и интервалом размеров δs = 0,73 µ м.
Динамика роста жировой ткани
Abstract
Рост жировой ткани происходит по двум механизмам: гиперплазия (увеличение количества клеток) и гипертрофия (увеличение размера клеток). Генетика и диета влияют на относительный вклад этих двух механизмов в рост жировой ткани при ожирении. В этом исследовании распределение по размеру эпидидимальных жировых клеток от двух линий мышей, устойчивых к ожирению FVB / N и склонных к ожирению C57BL / 6, измеряли через 2, 4 и 12 недель в условиях регулярного кормления и питания с высоким содержанием жира.Общее количество клеток в жировой подушке придатка яичка оценивали по массе жировой подушечки и нормализованному распределению клеток по размеру. Число клеток и средневзвешенный размер клеток увеличиваются в зависимости от массы жировой прослойки. Чтобы точно решить проблему роста жировой ткани, мы разработали математическую модель, описывающую эволюцию распределения размеров жировых клеток как функцию увеличения массы жировой ткани, а не увеличения хронологического времени. Наша модель описывает привлечение новых жировых клеток и их последующее развитие в разных штаммах и с разными режимами питания, с общими механизмами, но с параметрами модели, зависящими от диеты и генетики.По сравнению со штаммом FVB / N, штамм C57BL / 6 имеет большее количество мелких жировых клеток. Гиперплазия усиливается за счет диеты с высоким содержанием жиров в зависимости от штамма, что предполагает синергетическое взаимодействие между генетикой и диетой. Кроме того, кормление с высоким содержанием жиров увеличивает скорость роста жировых клеток, независимо от напряжения, что отражает увеличение количества калорий, требующих хранения. Кроме того, диета с высоким содержанием жиров приводит к резкому увеличению распределения жировых клеток по размерам у обоих штаммов; это означает увеличение колебаний размера жировых клеток за счет липидного обмена.
Авторское резюме
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. Динамическое изменение жировой ткани in vivo невозможно отслеживать в режиме реального времени из-за текущих технических ограничений. Однако мы можем измерить распределение жировых клеток по размеру у отдельных животных. Наша основная цель — извлечь динамические характеристики ремоделирования тканей из снимков распределения клеток по размерам.Мы разрабатываем математическую модель, которая интерполирует измерения распределения клеток по размеру и предсказывает непрерывное изменение распределения клеток по размеру в зависимости от увеличения массы жировых подушечек. Наша модель роста жировой ткани включает три основных компонента: рекрутирование новых клеток, рост клеток в зависимости от размера и колебания размера клеток. В частности, мы сравнили рост жировой ткани у склонных к ожирению и устойчивых к ожирению мышей при стандартной диете или диете с высоким содержанием жиров, чтобы изучить генетическое и диетическое влияние на рост жировой ткани.Применяя нашу модель к этим различным условиям, мы обнаружили, что увеличение размера жировых клеток зависит от диеты. С другой стороны, увеличение количества жировых клеток, вызванное диетой, зависит от деформации, что предполагает синергию между генетикой и диетой.
Введение
Ожирение — это увеличение жировой ткани для хранения избыточного количества потребляемой энергии. Гиперплазия (увеличение числа клеток) и гипертрофия (увеличение размера клеток) — два возможных механизма роста. На фенотип ожирения жировой ткани влияют диета и генетика, а также их взаимодействие [1] — [4].Начиная с исследований Джонсона и Хирша [5], существует обширная литература о росте жировой ткани при нормальном и патологическом развитии, характеризующая состояние ткани с точки зрения среднего размера и количества клеток. Гиперпластический рост появляется только на ранних стадиях развития жировой ткани [6], [7]. Гипертрофия возникает до гиперплазии, чтобы удовлетворить потребность в дополнительных накоплениях жира при прогрессировании ожирения [8]. Однако оказалось трудно понять, как диета и генетика конкретно влияют на гиперплазию и / или гипертрофию жировых клеток из-за ограниченных данных о росте жировой ткани.
Помимо этих исследований, недавно стало возможным точно измерить распределение клеток по размерам. Эта подробная информация, по сравнению со средним размером клеток и общим числом клеток, может использоваться для вычисления многих величин, связанных с размером, которые позволяют более точно охарактеризовать процесс роста жировой ткани. Кумулянты распределения клеток по размерам могут использоваться для вычисления других физиологических величин, таких как средневзвешенный размер клеток. Распределение клеток по размерам можно использовать для оценки общего количества клеток в жировой подушке по ее массе.Более того, считается, что некоторые специфические метаболические свойства, например, резистентность к инсулину [9] и секреция адипокина [10], зависят от точного распределения клеток по размерам, а не от среднего размера клеток. Действительно, в нескольких исследованиях изучается изменение распределения жировых клеток по размерам в различных условиях при развитии куриного эмбриона [11], у худых и страдающих ожирением крыс Zucker [12], [13], крыс линии Wistar с частичной липэктомией [14], биопсии кролика [ 15] и жировой ткани человека [16], [17]. Эти исследования были сосредоточены только на статических различиях между распределениями клеток по размерам в разных условиях.Однако статические распределения размеров клеток в поперечном сечении для ряда снимков развития животных можно использовать для вывода динамики роста жировой ткани, если мы сможем надлежащим образом проанализировать снимки с помощью математического моделирования. Учитывая нынешние технические ограничения, это может быть лучший доступный подход к микроскопическому и продольному пониманию роста жировой ткани in vivo , хотя в недавнем эксперименте были получены микроскопические наблюдения накопления липидов в липидных каплях жировых клеток [18].
Для изучения генетического и диетического влияния на динамический процесс роста жировой ткани мы получили распределения по размерам клеток эпидидимального жира у устойчивых к ожирению линий мышей FVB / N (далее FVB) и склонных к ожирению линий мышей C57BL / 6 (C57) в соответствии с стандартные диеты и диеты с высоким содержанием жиров. Мышь C57 является наиболее охарактеризованной моделью ожирения, вызванного диетой [19], а мышь FVB является предпочтительной моделью для создания трансгенных мышей [20]. Эти два широко используемых инбредных штамма мышей генетически довольно далеки [21], [22] и имеют различные метаболические фенотипы: по сравнению с мышами FVB, мыши C57 имеют низкие уровни циркулирующих триглицеридов [21] и повышенный клиренс триглицеридов [23], [24]; Мыши FVB характеризуются относительно более высокой печеночной инсулинорезистентностью, контррегуляторной реакцией на гипогликемию и сниженной глюкозо-стимулированной секрецией инсулина [25]; Также известно, что мыши FVB являются спонтанно гиперактивными [26] и относительно худыми, поскольку они, по-видимому, менее восприимчивы к диете с высоким содержанием жиров, чем мыши C57 [27].Однако развитие ожирения, вызванного диетой, у этих двух штаммов формально не сравнивалось. В этом исследовании мы разработали математическую модель, прогнозирующую изменение распределения клеток по размерам в зависимости от массы жировой подушечки придатка яичка, для количественного анализа динамических характеристик, зависящих от генетики и / или диеты. Модель роста жировой ткани описывает, сколько новых клеток образуется, как каждая клетка растет в зависимости от ее размера и как липидный обмен приводит к колебаниям размера, которые вызывают расширение распределения клеток по размерам.По мере увеличения массы эпидидимальных жировых подушек распределение клеток по размерам систематически изменяется в зависимости как от генетики, так и от диеты. Сравнивая экспериментальные результаты с теоретической моделью роста, мы обнаружили, что гипертрофия сильно коррелирует с диетой. С другой стороны, гиперплазия зависит от генетики. Изменения при гиперплазии, вызванные диетой, зависят от штамма, что предполагает взаимодействие между диетой и генетикой.
Результаты
Влияние диеты с высоким содержанием жиров на состав тела мышей FVB и C57
В начале эксперимента (в возрасте 5 недель) мыши C57 были значительно легче, чем мыши FVB () из-за разницы в худой масса, хотя общая жировая масса не отличалась ().Когда мышей поддерживали на обычном питании, разница в массе тела исчезла к возрасту 11 недель (6-я неделя эксперимента). В условиях обычной диеты мыши FVB и C57 сохраняли сопоставимую массу жира на протяжении всего эксперимента (). Диета с высоким содержанием жиров вызвала значительное увеличение массы тела и жировой массы у обоих штаммов; однако изменения массы тела и жировой массы были более значительными у мышей C57. У мышей C57 было вдвое больше жира после 12 недель кормления с высоким содержанием жиров ().Общая разница в общей массе жира между мышами FVB и C57 коррелировала с пропорциональными различиями в количестве эпидидимального (внутрибрюшного), пахового (подкожного) и бурого жира (). Потребление калорий и активность были сопоставимы у мышей FVB HF и C57 HF; однако мыши FVB HF имели более высокий уровень покоя и общего потребления кислорода, а также более высокую ректальную температуру, что позволяет предположить, что увеличение расхода энергии, а не снижение потребления калорий, было причиной относительной устойчивости к ожирению, вызванному диетой с высоким содержанием жиров, у мышей FVB.Интересно, что в течение первых 2 недель кормления с высоким содержанием жиров мыши FVB и C57 показали сопоставимое увеличение общей жировой массы (). Мыши C57 HF продолжали быстро увеличивать жировую массу до 10 недели эксперимента, тогда как мыши FVB HF замедляли накопление жира около недели 3. У мышей C57 кормление с высоким содержанием жира вызывало постепенное увеличение как эпидидимальных, так и паховых жировых подушечек; Напротив, у мышей FVB жировая масса придатка яичка увеличилась лишь незначительно после 4 недель кормления с высоким содержанием жира, в то время как паховая жировая подушечка продолжала увеличиваться в размере на протяжении всего эксперимента (рис.S1). Кормление с высоким содержанием жиров вызывало значительное повышение уровня глюкозы и инсулина в крови как у мышей FVB, так и у мышей C57 (). Уровни инсулина и непереносимость глюкозы были выше у мышей C57 HF, чем у мышей FVB HF, что свидетельствует о более тяжелой инсулинорезистентности (). В соответствии с предыдущими сообщениями [23], [24], мыши C57 REG показали пониженные уровни триглицеридов в сыворотке по сравнению с мышами FVB REG без разницы в FFA (). Это происходило не из-за более высокой утилизации жира, так как коэффициент респираторного обмена () и скорость окисления жирных кислот, измеренная in vivo, () и в изолированных скелетных мышцах () были сопоставимы у мышей FVB REG и C57 REG.Более вероятно, что более низкие уровни триглицеридов в сыворотке у мышей C57 REG были вызваны гораздо более эффективным клиренсом циркулирующих триглицеридов, как предполагает тест на клиренс триглицеридов (). Кормление с высоким содержанием жиров снижало уровни циркулирующих триглицеридов как у мышей FVB, так и у мышей C57 и улучшало клиренс триглицеридов у последних штаммов (и). Оба штамма показали сопоставимое снижение коэффициента респираторного обмена, что свидетельствует о сопоставимом увеличении утилизации жирных кислот в условиях диеты с высоким содержанием жиров (). Взятые вместе, эти данные предполагают, что способность эффективно выводить триглицериды из кровотока может способствовать высокой способности накопления жира у мышей C57.
Изменения в составе тела мышей FVB и C57 в условиях стандартного корма (REG) и с высоким содержанием жира (HF).(A) Масса тела. (B) Жировая масса. Состав тела измеряли в указанные моменты времени у 8 мышей в группе с использованием МРТ-анализатора Echo 3-in-1. Высокожировое и контрольное кормление начали в возрасте 5 недель. Ось X показывает недели контролируемого кормления. Показан один из трех независимых экспериментов. Приведенные значения являются средними ± SEM.
Физиологические характеристики мышей FVB и C57.(A) Тест на толерантность к глюкозе был проведен на 10-й неделе контролируемого кормления с пищей и диетой с высоким содержанием жиров. Мышей не кормили в течение ночи и вводили глюкозу (2 мг / г, внутрибрюшинно). Каждая группа была представлена 8 мышами. (B) Окисление олеиновой кислоты в целом организме измеряли у самцов мышей FVB ad C57 в возрасте 10 недель в не голодном состоянии (8 мышей на группу), как описано в Gautam et al. [47]. (C) Окисление пальмитиновой кислоты измеряли в камбаловидной мышце, взятой у 10-недельных мышей FVB и C57, которых содержали на диете (8 мышей на группу).Образцы собирали в 8 часов утра у мышей, которых случайным образом кормили, или мышей, голодавших в течение 18 часов [методы, описанные в Toyoshima et al. [46]]. (D) Клиренс триглицеридов у самцов мышей FVB и C57 после 11 недель контролируемого кормления кормом и диетой с высоким содержанием жиров. Мышей не кормили в течение 4 часов, а затем вводили через желудочный зонд 400 мкл л арахисового масла. Плазму собирали ежечасно в течение 5 часов из хвостовой вены для измерения триглицеридов. Каждая группа была представлена 8 мышами.
Таблица 1
Характеристики мышей FVB и C57 после 12 недель высокожирового и контрольного кормления.
| FVB REG a | FVB HF a | C57 REG a | C57 HF | a 94 996| 912 912 912 912 912 Взаимодействие б | | ||||||||||
| Масса тела (г) | 29,1 ± 0,4 | 35,4 ± 1,4 | 29,1 ± 0,4 | 43,0 ± 1,8 | 0,003 | <0,001 | 0,003 | г)0.47 ± 0,03 | 1,23 ± 0,11 | 0,67 ± 0,08 | 2,00 ± 0,21 | <0,001 | <0,001 | 0,032 | |
| Паховой жир (г) | 0,3412 ± 0,02 | 0,190,3412 ± 0,02 | 0,3412 ± 0,02 | 0,45 ± 0,041,92 ± 0,27 | 0,003 | <0,001 | <0,003 | ||||||||
| Коричневый жир (г) | 0,14 ± 0,01 | 0,27 ± 0,04 | 0,1412 ± 0,01 | 0,01 0,09 0,003<0.001 | 0,003 | ||||||||||
| Потребление пищи (ккал / мышь / день) | 16,6 ± 1,0 | 18,0 ± 1,3 | 13,2 ± 0,8 | 17,7 ± 0,8 | 0,07 | 0,007 92| 3 ns Общее потребление кислорода (мл / г) | 9,5 ± 0,2 | 11,6 ± 0,3 | 9,0 ± 0,2 | 9,4 ± 0,2 | <0,001 | <0,001 | нс | | |
| Потребление кислорода в покое (мл / г) ) | 8.3 ± 0,2 | 9,7 ± 0,3 | 7,3 ± 0,2 | 8,1 ± 0,3 | <0,001 | <0,001 | 0,003 | ||||||||
| Коэффициент респираторного обмена | 0,86 ± 0,01 | 0,75 ± 0,01 0,01 | 0,72 ± 0,00 | нс | <0,001 | 0,006 | |||||||||
| Суммарная активность (торможение луча / мин) | 265 ± 5 | 190 ± 17 | 345 ± 41 | 122011293 нс | <0.001 | нс | |||||||||
| Ходьба (торможение луча / мин) | 105 ± 3 | 53 ± 10 | 144 ± 25 | 38 ± 5 | нс | <0,002 | 91 Ректальная температура (° C) | 36,0 ± 0,2 | 36,6 ± 0,2 | 35,4 ± 0,2 | 35,5 ± 0,2 | <0,001 | 0,057 | нс | |
| Триглицерид сыворотки (мг / дл) | ± 25 | 95 ± 24 | 89 ± 16 | 64 ± 6 | 0.002 | 0,004 | ns | ||||||||
| Свободные жирные кислоты (мМ) | 0,22 ± 0,03 | 0,22 ± 0,01 | 0,26 ± 0,04 | 0,27 ± 0,05 | 1292 ns | ns | ns Глюкоза крови (мг / дл) | 187 ± 16 | 225 ± 11 | 209 ± 11 | 247 ± 15 | нс | 0,009 | нс | |
| Инсулин сыворотки (мг / | 1,1) 912 ± 0,1 | 1,9 ± 0,3 | 2.2 ± 0,8 | 4,9 ± 1,2 | 0,011 | 0,033 | нс | ||||||||
| Клиренс триглицеридов (AUG c ) | 3238 ± 476 | 1651 ± 160 | 48 | 489 | <0,001 | 0,011 | <0,001 | ||||||||
| Толерантность к глюкозе (AUG d ) | 28633 ± 1372 | 39560 ± 3217 | 25947 ± 1212 | 001 | 0,003 |
Гипертрофия и гиперплазия с увеличением жировой массы
Чтобы проверить основной механизм различной скорости накопления жира в эпидидимальном жире мышей FVB и C57, мы измерили распределение массы и размеров клеток в образцах тканей эпидидимальный жир, собранный на 0, 2, 4 и 12 неделях контролируемого кормления (). Поскольку гистологический анализ не позволяет точно определить размер клеток адипоцитов, который будет обсуждаться позже, мы измерили распределение клеток по размеру с помощью счетчика Коултера и рассчитали средневзвешенный размер клеток и общее количество клеток эпидидимальной жировой подушки по этим измерениям, которые аналогичные значения в другом исследовании на мышах [5].Сильная корреляция наблюдалась между массой жировых подушечек и средневзвешенным размером клеток, а также между массой жировых подушечек и общим числом клеток, независимо от напряжения и разницы в диете (). Первая корреляция дала масштабное соотношение между массой жировой подушечки и средневзвешенным по объему размером клеток (). Кроме того, была обнаружена экспоненциальная зависимость между массой жировой подушечки, и общим числом клеток, где начальная масса жировой подушки была получена от контрольных мышей; и начальное количество клеток, и скорость увеличения количества клеток в массе жировой подушечки, были оценены по данным (;).Первоначальное количество клеток у мышей C57 было больше, чем исходное количество клеток у мышей FVB (фиг. S2C и S2D). По мере увеличения массы жировой ткани увеличивается общее количество клеток. Скорость увеличения числа клеток была выше при обычном питании, чем при диете с высоким содержанием жиров, и эта тенденция более очевидна у мышей C57 (;), что указывает на генетическое различие. Соотношение между результатами регулярной диеты и диеты с высоким содержанием жиров составляет 1,42 и 3,22 для мышей FVB и C57 соответственно (). Это может указывать на взаимодействие между генетикой и диетой при увеличении количества клеток.Обратите внимание, что мы также наблюдали аналогичные результаты с массой тела и массой жира, поскольку три величины (масса жировых подушечек придатка яичка, масса жира и масса тела) коррелируют друг с другом. Однако результаты с массой эпидидимальной жировой подушечки были наиболее подходящими: среднеквадратичное отклонение между данными и соответствием составило 9,73, 7,94, 3,56 × 10 5 и 5,54 × 10 6 , соответственно; результат с массой тела составил 11,82, 15,35, 3,65 × 10 5 и 4,82 × 10 6 ; результат с жировой массой 8.58, 8,61, 4,28 × 10 5 и 6,11 × 10 6 .
Нормализованное распределение клеток по размерам в жировой подушке придатка яичка.На графике нанесен один типичный пример контрольных мышей FVB в возрасте пяти недель. Кружки обозначают точки измеренных данных, а линия представляет собой аппроксимирующую кривую, которая представляет собой сумму одной гауссовой и двух экспоненциальных функций.
Таблица 2
Значения параметров модели роста жировой ткани.
Генетические и диетические эффекты на рост жировой ткани
Эти две сильные корреляции между массой жировой подушечки и гипертрофией и между массой жировой подушечки и гиперплазией позволяют предположить, что увеличение жировой ткани можно описать как систематический процесс роста в отношении для увеличения массы жировой прослойки.Мы расположили распределения клеток по размерам, отсортированные по массе эпидидимальных жировых подушечек (). Примечательно, что модель роста жировой ткани в формуле. (1) описывает эволюционирующую картину распределения клеток по размерам в зависимости от увеличения массы жировой прослойки. Модель количественно соответствовала экспериментальным распределениям размеров клеток, несмотря на то, что все распределения представляют собой данные поперечного сечения, полученные от отдельных животных. Различные значения параметров в модели, которые соответствуют каждому индивидуальному распределению размеров клеток для обоих штаммов и обоих режимов питания, дали количественные различия в процессе роста эпидидимальной жировой ткани между штаммами и между диетами ().Во-первых, максимальная зависящая от размера скорость роста и скорость колебаний размера клеток из-за обмена липидов продемонстрировали разницу, вызванную диетой, и меньшую разницу, вызванную деформацией. Зависимый от размера рост и колебания размера, приводящие к гипертрофии, по-видимому, регулируются в основном диетой. В частности, рост, зависящий от размера, сдвинул режим больших клеток в распределении размеров клеток в сторону больших размеров, а флуктуации липидного обмена увеличили разброс распределения вокруг режима больших клеток.Важно отметить, что результаты должны быть тщательно интерпретированы, потому что каждая скорость — это скорость на единицу увеличения массы жировой ткани, а не на единицу увеличения времени. Во-вторых, геометрические параметры (, и), которые определяют форму скорости роста, зависящей от размера, имели практически одинаковые значения независимо от диеты и разницы деформаций, за исключением. Следовательно, нижний критический размер, который дает размер, инициирующий рост клеток в зависимости от размера, и два параметра масштаба могут быть зафиксированы на с l = 37 µ m, η l = 12 µ м и η u = 63 µ м соответственно.С другой стороны, верхний критический размер, ограничивающий зависимый от размера клетки рост больших клеток, зависел от диеты; при кормлении с высоким содержанием жира этот пороговый размер сместился в сторону большего размера (). При диете с высоким содержанием жиров изменения параметров (,, и) увеличивают способность жировых тканей накапливать липиды за счет как гиперплазии, так и гипертрофии. Более низкие уровни триглицеридов в сыворотке при условии диеты с высоким содержанием жиров (и) могут коррелировать с увеличением накопления липидов в увеличенных жировых клетках, потому что не было обнаружено значительных различий в окислении жирных кислот, о чем свидетельствует отсутствие различий в соотношении респираторного обмена ().
Изменения распределения клеток по размерам с увеличением массы эпидидимальной жировой подушечки при кормлении и диетах с высоким содержанием жиров.Начальные распределения размеров клеток пятинедельных контрольных мышей приведены в первой строке каждого столбца пунктирными линиями. Точки показывают экспериментальные результаты. Обратите внимание, что результаты упорядочены по массе эпидидимальной жировой подушечки, а не по хронологическому времени (недели контролируемого кормления), указанному в скобках. Сплошные линии представляют собой нормализованные распределения клеток по размеру, соответствующие данной массе эпидидимальной жировой подушечки, предсказанной моделью роста жировой ткани.
Зависимая от размера скорость роста массы придатка яичка.Здесь для этого графика используются средние значения параметров. Символы показаны для облегчения сравнения строк.
Обсуждение
Наше центральное открытие состоит в том, что гиперплазия и гипертрофия жировых клеток в эпидидимальной жировой подушке является функцией массы жировой подушечки, даже несмотря на то, что у отдельных животных может потребоваться разное время для достижения заданной массы жировой подушечки. Следовательно, рост жировой ткани, представленный как изменения распределения клеток по размерам, можно систематически моделировать как процесс роста по отношению к увеличению массы жировой подушечки; это может отражать корреляцию между массой жировой подушечки и секрецией адипокинов и других сигнальных молекул, контролирующих рост жировой ткани.Соответственно, следует отметить, что нормы в нашей модели — это не обычные нормы увеличения на единицу времени, а нормы увеличения на единицу массы. Таким образом, несколько показателей (, и) в модели имели более высокие значения для животных, соблюдающих диету, чем для животных, соблюдающих диету с высоким содержанием жиров. Однако, если бы эти показатели были преобразованы в обычные нормы увеличения на единицу времени, они имели бы большие значения для диеты с высоким содержанием жиров, потому что требуется меньше времени для увеличения массы жировой подушечки на единицу из более крупных и более многочисленных клеток на диета с высоким содержанием жиров, чем для увеличения такой же величины за счет меньшего и меньшего количества клеток на диете (рис.S1). Было высказано предположение, что когда ожирение прогрессирует, рано возникает гипертрофия жировых клеток, а затем запускается гиперплазия [8]. Наше исследование показало, что рекрутирование новых клеток экспоненциально увеличивается с увеличением массы жировой ткани. Гипертрофия жировых клеток является основным фактором увеличения массы жировой прослойки, тогда как гиперплазия не способствует этому увеличению, потому что она возникает в небольших клетках, в которых хранится гораздо меньший объем жира. Таким образом, наша модель естественным образом воплощает идею о том, что на гиперплазию влияет гипертрофический рост клеток.С другой стороны, сообщалось, что гиперплазия жировых клеток возникает только на ранних стадиях развития; следовательно, не следует ожидать рекрутирования новых клеток на поздних стадиях даже в условиях ожирения [6], [7]. Возможно, возраст животных в нашем исследовании (6 недель) допускает возникновение гиперплазии.
Модель, разработанная здесь, может дать микроскопическое представление о росте жировых клеток, зависящем от размера, который не может быть рассмотрен статическими поперечными исследованиями.Например, мы обнаружили следующие специфические свойства роста клеток, зависящие от размера: нижний критический размер, инициирующий накопление липидов, не зависел от диеты у двух линий мышей, тогда как верхний критический размер ограничивал рост клеток до достижения необходимого уровня. необычайный размер, был увеличен на диете с высоким содержанием жиров. Эта зависимость роста клеток от размера является проверяемой гипотезой. Далее, параметр флуктуации размера клеток различался при обычном питании и рационе с высоким содержанием жиров; оно больше при диете с высоким содержанием жиров, когда оно преобразуется в единицы, соответствующие изменению единицы времени, а не единице изменения массы жировой подушечки.Таким образом, случайный процесс высвобождения и поглощения жира жировыми клетками происходит более активно при диете с высоким содержанием жиров, чем при обычной диете. Может быть интересно посмотреть, можно ли распространить эти результаты на другие штаммы и организмы.
По сравнению с исследованиями, в которых наблюдали единственный пик в распределении размеров клеток жировых клеток [11], [15] — [17], [28], мы наблюдали бимодальные распределения клеток по размерам, как сообщалось другими [12], [13], [29] — [32]. В большинстве исследований [12], [13], [31], [32] по наблюдению за бимодальностью использовалась технология счетчика Коултера, которая имеет несколько преимуществ для оценки всего распределения размеров ячеек [31]: Во-первых, можно доказать, что анализируемые ячейки быть подлинными жировыми клетками на основе морфологии и флотации; во-вторых, объем каждой клетки оценивается независимо от формы и без артефактов смещения центра, что является правилом, а не исключением, с использованием гистологических подходов; наконец, можно подсчитать и измерить достаточное количество частиц, чтобы получить статистически значимые комплексные кривые.Напротив, микроскопические методы гистологии могут не наблюдать мелкие клетки из-за влияния увеличения микроскопа [30], небольшого количества образцов и систематической ошибки выборки. Однако при использовании счетчика Коултера контаминация, не связанная с адипоцитами, может способствовать распределению клеток по размерам, особенно при малых размерах, хотя наш минимальный диаметр клеток, 22 µ м, превышает возможные диапазоны контаминации, от 10 до 20 µ м, упомянутый Mersmann и MacNeil [31]. Чтобы быть уверенным, мы снова проанализировали модифицированные данные, используя только распределения размеров клеток выше 35 µ м в диаметре с моделью, и пришли к тем же выводам (данные не показаны).Надир в распределении размеров клеток () может разделять две популяции клеток. ДеМартини и Франсендез определили маленькие клетки с диаметром менее 35 µ м как «очень маленькие жировые клетки» [29]. Согласно нашей модели, эти клетки имеют незначительный рост в зависимости от размера, поскольку их размер меньше нижнего критического размера, с l = 37 µ m. Следовательно, механизм роста, зависящий от размера, может естественным образом объяснить происхождение бимодальности в распределении жировых клеток по размеру.Клетки с размером только выше могут расти в зависимости от размера, но клетки с размером меньше могут расти случайным образом с флуктуацией размера за счет липидного обмена. Это разделение вызывает накопление клеток ниже размера, что дает более низкий пик в распределении размеров клеток. Эта популяция клеток может служить потенциальным резервуаром для зрелых жировых клеток. Процесс их созревания можно интерпретировать следующим образом: жировые клетки, достигающие критического размера, вследствие случайных колебаний размера могут расти с механизмом роста, зависящим от размера.Как упоминалось выше, изменение размера происходит более активно при диете с высоким содержанием жиров; следовательно, пласт может ускорить процесс созревания в условиях стимуляции.
В модель роста ткани мы включили привлечение новых клеток и рост существующих клеток, но не гибель старых клеток, потому что модель соответствовала данным без апоптоза жировых клеток. Этот результат также согласуется с исследованием, в котором наблюдалось, что жировая ткань придатка яичка мышей C57BL / 6 не демонстрирует значительной гибели клеток до 12 недель при диете с высоким содержанием жиров [33].Однако расширенная диета с высоким содержанием жиров в конечном итоге вызывает апоптоз жировых клеток [33]. Более того, в одном недавнем исследовании сообщается, что жировые клетки человека обновляются в течение десяти лет [7]. Таким образом, наша модель все еще нуждается в улучшении, чтобы ее можно было использовать в различных условиях. Следует учитывать гибель клеток и формально учитывать зависимость параметров модели от диеты. В этом исследовании мы сосредоточились на одном жировом депо, эпидидимальном жире, по нескольким причинам: 1) вес эпидидимальных жировых подушечек может быть определен более точно, чем вес паховых жировых подушечек из-за легкости иссечения; 2) морфология жировых клеток в эпидидимальном жире более однородна, чем в паховом жире, который содержит много клеток, подобных коричневой жировой ткани, особенно у мышей, устойчивых к ожирению, вызванному диетой; и 3) разница между генотипами была более очевидной в росте эпидидимального жира, который достигает плато через 4 недели у мышей FVB, но продолжает расти у мышей C57 на протяжении всего эксперимента, в отличие от пахового жира, который показывает устойчивый рост. у обоих штаммов (рис.S1). Хотя мы не измеряли распределение по размеру клеток других жировых отложений, мы измерили изменение массы паховых и бурых жировых отложений, что показывает аналогичную картину с эпидидимальным жировым депо (рис. S1). Таким образом, представляет интерес применить модель к другим жировым отложениям, которые имеют функциональные различия [34], [35], а также к другим видам, таким как человек, что остается для будущих исследований. Мы ожидаем, что модель может быть применена к таким разнообразным наборам данных, просто настроив параметры модели, потому что модель содержит общие механизмы роста тканей для привлечения новых клеток и их последующего развития.
Наши данные предполагают, что, по крайней мере, три фактора могут объяснить, почему мыши C57 набирают больше жира, чем мыши FVB, при диете с высоким содержанием жиров: во-первых, мыши FVB имеют повышенную скорость метаболизма и повышенную ректальную температуру, скорее всего, из-за повышенного симпатического тонуса. Хотя мы не обнаружили значительных различий в активности между штаммами в нашем исследовании, более подробные измерения поведения показали, что мыши FVB спонтанно гиперактивны по сравнению с мышами C57 [26]. У них также увеличивается частота сердечных сокращений [36], и они реагируют гипергликемией на различные виды лечения [37].Кроме того, они более чувствительны к стрессу, связанному с воздержанием и голоданием [38]. Все эти данные, взятые вместе, предполагают, что активность симпатической нервной системы увеличилась больше у мышей FVB, чем активность у мышей C57. Во-вторых, по сравнению с мышами FVB, мыши C57 очищают циркулирующие триглицериды более эффективно [23], [24], что, по крайней мере, частично можно объяснить более высокой активностью липазы в сыворотке крови [23] и более высокой способностью накапливать триглицериды в печени [23]. ], [24] и жировой ткани (показано в этом исследовании).Хотя молекулярные механизмы клиренса триглицеридов до конца не изучены, жировая ткань явно способствует выведению триглицеридов, поскольку способность выводить циркулирующие триглицериды нарушена у липоатрофных мышей [23]. В частности, сообщалось, что диета с высоким содержанием жиров увеличивает клиренс триглицеридов [39], что может быть связано с индукцией активности липопротеинлипазы в жировой ткани [40]. Наконец, по сравнению с мышами FVB, мыши C57 показали большее количество мелких жировых клеток, особенно при диете с высоким содержанием жиров.Было высказано предположение, что новые жировые клетки могут возникать из клеток-предшественников, которые находятся в депо белого жира взрослых [41], [42], и из других источников, таких как циркулирующие клетки-предшественники костного мозга [43]. Было показано, что рекрутирование обоих типов предшественников стимулируется диетой с высоким содержанием жиров [41], [43]. Возможно, что большее рекрутирование более мелких жировых клеток у мышей C57 может быть вызвано более высоким пулом клеток-предшественников или их более высокой внутренней способностью к дифференцировке адипоцитов.Однако in vitro мезенхимальных стволовых клеток , выделенных из наружного уха мышей FVB и C57, одинаково хорошо дифференцируются в жировые клетки [44]. Попытка сравнить дифференцировку стромальных клеток костного мозга мышей FVB и C57 в жировые клетки не увенчалась успехом из-за очень низкого выхода и плохой пролиферативной способности клеток, выделенных от мышей C57 [45]; однако костный мозг, по-видимому, не является основным источником новых жировых клеток, по крайней мере, у мышей [43]. Наша модель предполагает, что разница между генотипами в рекрутировании малых жировых клеток может быть автономной по отношению к жировым подушечкам, но молекулярный механизм, лежащий в основе этого различия, неясен.Жировая подушечка представляет собой сложный орган, содержащий множество различных типов клеток, включая зрелые жировые, предадипозные и сосудистые клетки, нервы, макрофаги и фибробласты. Количество предшественников адипоцитов и их пролиферация в ответ на внешние сигналы варьируются в зависимости от жировых отложений [35]. Необходимы дальнейшие исследования, чтобы определить, как специфичное для генотипа взаимодействие между разными типами клеток и секретируемыми факторами может влиять на скорость рекрутирования адипоцитов в определенные жировые депо.
Таким образом, мы получили математическую модель, описывающую рост жировой ткани с увеличением количества и размера клеток в зависимости от массы жировой подушечки придатка яичка.Основываясь на этой динамической модели, мы исследовали влияние генетики и диеты на рост жировой ткани. Сравнивая распределение размеров клеток двух штаммов и двух диет, мы пришли к выводу, что изменение размера клеток зависит от диеты, а изменение количества клеток зависит от генетики и диеты, а также от их взаимодействия.
Материалы и методы
Животные
Все процедуры были одобрены Комитетом по уходу и использованию животных Национального института диабета, болезней органов пищеварения и почек.Самцы мышей FVB и C57 были получены из лаборатории Джексона (Бар-Харбор, штат Мэн). Мышей выращивали по четыре в клетке с 12-часовым циклом свет / темнота (свет 06: 00–18: 00). В возрасте 5 недель мышей каждой линии разделили на 2 группы. Половине мышей давали обычную пищу NIH-07 (далее REG; Zeigler Brothers, Inc., Gardners, PA), содержащую 4,08 ккал / г (11% калорий из жира, 62% из углеводов и 26% из белка). Другая половина получала диету с высоким содержанием жиров, F3282 (далее HF; Bio-Serv, Frenchtown, NJ), содержащую 5.45 ккал / г (59% жиров, 26% углеводов и 15% белков). Вода и диета были предоставлены ad libitum . Было проведено пять независимых экспериментов, в каждой из которых использовалось 4 группы мышей: FVB REG, FVB HF, C57 REG и C57 HF. В трех экспериментах мышей содержали на контролируемой диете в течение 12 недель и использовали для анализа состава тела, физиологической характеристики и распределения клеток по размеру. Две дополнительные группы мышей были умерщвлены после 2 недель и 4 недель контрольного кормления с высоким содержанием жира только для определения распределения клеток по размеру.
Эксперименты in vivo
Состав тела, потребление пищи, скорость метаболизма, толерантность к глюкозе, клиренс триглицеридов и окисление жирных кислот в изолированной камбаловидной мышце измеряли, как описано ранее [46]. Окисление жирных кислот всего тела измеряли, как описано в Gautam et al. [47]. Кровь для биохимических анализов брали из хвостовой вены без голодания. Уровни глюкозы измеряли с помощью Glucometer Elite (Bayer, Elkhart, IN). Сывороточный инсулин анализировали с помощью радиоиммуноанализа (Linco Research, St.Чарльз, Миссури). Триглицериды сыворотки, холестерин (Thermo DMA, Луисвилл, Колорадо) и свободная жирная кислота (FFA) (Roche Applied Science, Индианаполис, Индиана) измеряли в соответствии с процедурами производителя.
Измерение размера клеток эпидидимального жира
Распределение размеров клеток эпидидимального жира измеряли через 2, 4 и 12 недель высокожирового и контрольного кормления с использованием Beckman Coulter Multisizer III, как описано ранее [9]. Вкратце, 20–30 мг жировой ткани были взяты из средней части путем рассечения и последующего удаления образца для фиксации из центра разрезанного эпидидимального жира.Образцы тканей немедленно фиксировали в четырехокиси осмия [48], инкубировали на водяной бане при 37 ° C в течение 48 ч, а затем определяли размер жировых клеток на приборе Beckman Coulter Multisizer III с апертурой 400 µ м. Диапазон размеров ячеек, которые можно эффективно измерить с помощью этой апертуры, составляет 20–240 µ м. Прибор был настроен на подсчет 6000 частиц, и суспензия фиксированных клеток была разбавлена так, чтобы совпадающий счет был <10%. После сбора размеров импульсов данные были выражены в виде диаметров частиц и отображены в виде гистограмм отсчетов по диаметру с использованием линейных интервалов и линейной шкалы для оси x ().Распределение клеток по размерам было измерено в четырех образцах из каждой группы, за исключением мышей C57 после 4-недельного воздействия диеты с высоким содержанием жиров, у которых было только три доступных образца. Образец был взят из каждой жировой подушечки и обработан отдельно. Затем каждый образец был посчитан как минимум дважды. Затем кривые для двух образцов усредняются, но только после проверки воспроизводимости между двумя образцами.
Модель роста жировой ткани
Распределение размеров клеток включает всю информацию, относящуюся к размерам клеток в ткани, и его изменения дают статистическое представление о детальном процессе роста каждой клетки.Чтобы изучить рост жировой ткани с точки зрения лежащих в основе микроскопических процессов, мы рассматриваем математическую модель, количественно определяющую процессы, которые изменяют распределение клеток по размерам. Модель может предсказать, сколько новых клеток образуется и как клетки разного размера растут по мере увеличения массы жировой ткани. Плотность количества клеток определенного размера (диаметра) при данной массе жировой подушечки — это конкретное количество, которое необходимо учитывать. Мы рассматриваем, как это распределение клеток по размерам изменяется с постепенным изменением массы жировой ткани.Эволюция распределения клеток по размерам с массой жировой ткани может быть смоделирована уравнением в частных производных:
(1)
Это уравнение включает три основных компонента процесса роста жировой ткани. Во-первых, мы предполагаем, что рекрутирование новых клеток происходит только при минимальном наблюдаемом размере клетки, который математически выражается как дельта-функция. Скорость набора по отношению к массе жировой подушечки определяется экспоненциальной функцией:
(2)
, где — исходное общее количество клеток при данной начальной массе жировой подушечки, а — скорость увеличения количества клеток на единицу изменения. в массе жировой подушечки.Изменение общего числа клеток — это скорость пополнения новых клеток, если гибель клеток незначительна; мы не обнаружили необходимости включать апоптоз в клетки любого размера, чтобы соответствовать этим экспериментальным данным. Следовательно, этот коэффициент пополнения может быть непосредственно получен из экспериментального результата, используя соотношение между общим числом клеток и массой жировой подушечки путем дифференцирования функции по отношению к. Во-вторых, рост клеток зависит от размера клетки. После созревания жировых клеток до определенного размера они могут накапливать жир, вызывая гипертрофию.В дополнение к этой ограничивающей скорости роста маленьких жировых клеток, может также существовать верхний предел роста, потому что большие жировые клетки не могут расти бесконечно в результате этого процесса роста, хотя они могут достигать больших размеров из-за колебаний размера, вызванных обменом липидов. Повышение и снижение скорости роста клеток в зависимости от размера клетки можно описать с помощью общей функциональной формы, умножающей две сигмоидальные функции,
(3)
, где представляет максимальную скорость роста; и — соответственно нижний и верхний критические размеры, которые дают половину максимальной скорости роста; и укажите их масштаб ().Наконец, последний член в формуле. (1) представляет колебания размера клеток с постоянной скоростью, которые отражают обмен липидов, случайным образом происходящий в жировых клетках. Этот липидный оборот является единственным механизмом роста для крупных клеток, размер которых превышает верхний критический размер. Как правило, рост клеток, зависящий от размера, приводит к увеличению их распределения по размерам, в то время как колебания размеров распространяют распределение по размерам.
Метод оптимизации
Чтобы оптимизировать параметры модели, чтобы они могли точно описывать эволюцию распределения размеров ячеек в эксперименте, мы использовали минимизацию «функции стоимости», которая количественно определяет отклонение между моделью и результатами эксперимента.Чтобы определить функцию стоимости, нормализованное распределение размеров клеток при заданной массе жира сравнивалось с данными моделирования с набором параметров:
(6)
, где — общее количество интервалов размера клеток, а — общее количество заданных масса жира. Масштаб функции стоимости был рассчитан на основе внутренней флуктуации экспериментальных данных, которую можно определить как квадрат отклонения между измеренным распределением ячеек по размерам и функцией плавной аппроксимации:
(7)
Эта внутренняя флуктуация численно составляет около 10 процентов квадрата отклонения между экспериментальными и модельными данными.В качестве функции аппроксимации мы использовали сумму двух экспонент и гауссову форму,
(8)
, форму, которая использовалась для аппроксимации распределений размеров жировых клеток [9]. Подбор этих параметров был выполнен с использованием процедуры подбора нелинейной кривой в MATLAB R2007a (Натик, Массачусетс, США). Для процесса оптимизации мы специально использовали метод Монте-Карло с параллельным темперированием, чтобы найти глобальный минимум функции стоимости [50]. Мы использовали 10 равномерно распределенных значений (от 0,1 до 1) для параметра темперирования и запускали десять цепочек параллельно с вероятностью обновления.На каждых 20 шагах случайным образом выбиралась пара смежных имитаций по десяти параметрам отпуска, и их состояния параметров менялись с вероятностью. После достижения равновесия было использовано 20 000 итераций с фиксированным параметром отпуска для оценки оптимальных значений параметров и их стандартных ошибок.
Мы также использовали этот метод для оценки исходного общего количества клеток, и скорости его увеличения, исходя из соотношения между жировой массой и общим количеством клеток (). При минимизации функции аппроксимации и экспериментальных данных мы использовали ограничение, согласно которому начальное количество клеток одинаково независимо от условий диеты, т.е.е., обычные и жирные диеты. Средняя жировая масса четырех контрольных мышей перед регулярной диетой и диетой с высоким содержанием жиров была использована в качестве начальной жировой массы, которая составляет 0,34 г и 0,29 г для мышей FVB и C57, соответственно. Мы оценили погрешности и, распространив 10-процентные статистические флуктуации, наблюдаемые в экспериментальных данных.
Численное решение дифференциального уравнения с частными производными
Мы решили следующую дискретную версию нашей модели, заданную как непрерывное дифференциальное уравнение с частными производными в формуле.(1):
(9)
с интервалом масс δm = 0,1 мг и интервалом размеров δs = 0,73 µ м.
Исследования жировой ткани человека. Размер и количество жировых клеток у пациентов без ожирения и ожирения
Исследован клеточный характер жировой ткани 21 пациента без ожирения и 78 пациентов с ожирением. Размер жировых клеток (липидов на клетку) определяли в трех различных подкожных и глубоких жировых отложениях у каждого пациента, а общее количество жировых клеток в организме оценивали путем деления общего жира тела на различные комбинации размеров жировых клеток в шести разных участках. .Число клеток также оценивалось на основе различных предполагаемых распределений общего жира между подкожными и глубокими жировыми отложениями. Пациенты с ожирением, как группа, имеют более крупные жировые клетки, чем пациенты без ожирения; размер клеток, однако, значительно различается среди жировых отложений людей каждой группы. Различия в размере клеток существуют не только между подкожными и глубокими участками, но и внутри них. Таким образом, оценки общего количества жировых клеток для данного человека, основанные на размере клеток, могут варьироваться на 85%.На основе этих исследований предполагается, что общее количество жировых отложений человека лучше всего и наиболее практически оценивается в настоящее время путем деления общего количества жира в организме на среднее значение размеров жировых клеток по крайней мере трех подкожных участков. НЕ ОТВЕТИТЕЛЬНО ОТ МЕТОДА, ПО КОТОРЫМ ОЦЕНИВАЕТСЯ ОБЩЕЕ ЧИСЛО ЖИРОВЫХ КЛЕТОК, В ОТНОШЕНИИ КЛЕТОЧНОГО ХАРАКТЕРА ЖИРОВОЙ МАССЫ У ЭТИХ ПАЦИЕНТОВ ОБЪЯВЛЯЮТСЯ ДВЕ ОБРАЗЦА ОЖИРЕНИЯ: гиперпластический, с повышенным числом жировых клеток и нормальным или увеличенным размером. только увеличенный размер ячейки.Эти две клеточные модели ожирения не зависят от множества предполагаемых распределений жира между подкожными и глубокими отложениями. Когда эти различные клеточные структуры исследуются с точки зрения различных аспектов размера тела, состава тела, а также степени, продолжительности и возраста начала ожирения, только последнее однозначно отличает гиперпластическое от гипертрофического: гиперпластическое ожирение характеризуется ранним ожирением. возраст начала, гипертрофия, к позднему возрасту начала. Эти исследования показывают, что есть два различных периода в раннем периоде жизни, в течение которых наиболее вероятно возникновение гиперцеллюлярности жировой ткани: очень рано в течение первых нескольких лет и снова в возрасте от 9 до 13 лет.
.