Какой уровень гемоглобина считается нормальным? | Вечные вопросы | Вопрос-Ответ
Уровень гемоглобина (в переводе с др.-греч. αἷμα — кровь и лат. globus — шар) в крови является одним из важнейших показателей состояния здоровья человека. Именно с помощью этого сложного белка происходит насыщение каждой клеточки тела кислородом (O2), что позволяет нашему организму в буквальном смысле дышать и функционировать. Также это вещество переносит углекислый газ (CO2) от клеток в легкие, очищая тем самым внутренние органы от продуктов распада.
Что входит в его состав?
Гемоглобин состоит из двух частей: белка (глобин) и соединения железа (гема), делающего кровь красной при соединении с кислородом. Когда кровь омывает легкие, ионы железа притягивают кислород и, насыщая кровь этим живительным газом, разносят его по всем органам. Здесь же происходит обратный процесс: ионы железа отдают кислород, а к работе приступает белок глобин — он образует соединение с углекислым газом (кровь становится темной) и через легкие выводит его из организма.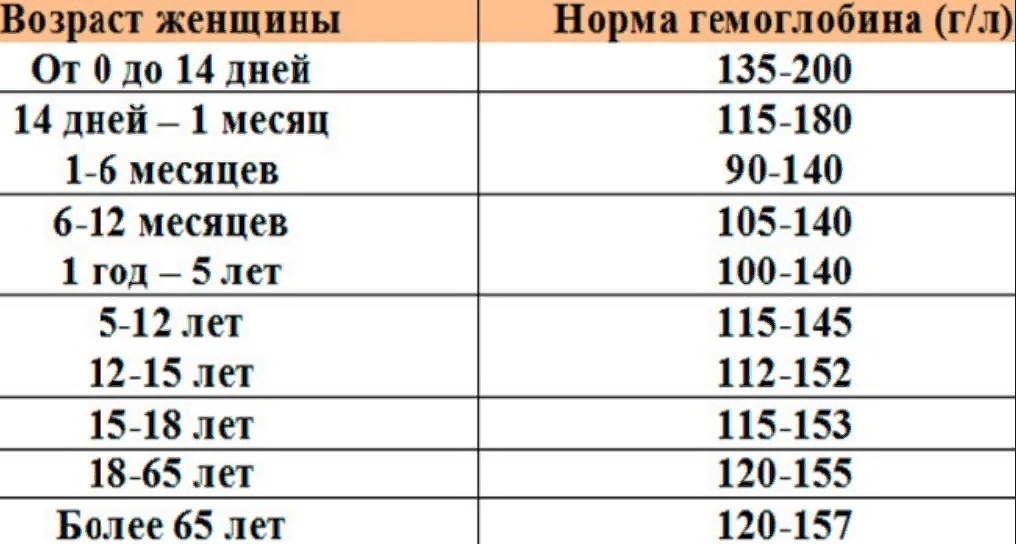
Норма содержания гемоглобина в крови человека
В раннем возрасте уровень гемоглобина у детей меняется очень часто. Максимальная концентрация гемоглобина в крови ребенка наблюдается в первые дни его жизни. Потом она постепенно снижается сначала от недели к неделе, затем от месяца к месяцу до полугода. Этот уровень сохраняется вплоть до пяти лет, после чего вновь несколько возрастает.
|
Возраст |
Гемоглобин г/л (грамм на литр) |
|
Новорожденный – первые три дня |
145-225 |
|
Первые семь дней |
135-215 |
|
Первые четырнадцать дней |
125-205 |
|
Месячный ребенок |
110-175 |
|
Два месяца |
90-140 |
|
От 3 до 6 месяцев |
90-135 |
|
От 6 месяцев до 5 лет |
105-140 |
|
От 5 до 12 лет |
115-145 |
По достижении ребенком 12 возраста судить об уровне его гемоглобина следует уже с учетом половой принадлежности.
|
Возраст |
Гемоглобин г/л (грамм на литр) у мальчиков |
Гемоглобин г/л (грамм на литр) у девочек |
|
От 12 до 15 лет |
120-160 |
112-152 |
|
От 15 до 18 лет |
117-160 |
115-153 |
Нормальные показатели гемоглобина в крови совершеннолетних людей остаются примерно на одном и том же уровне, вплоть до преклонного возраста.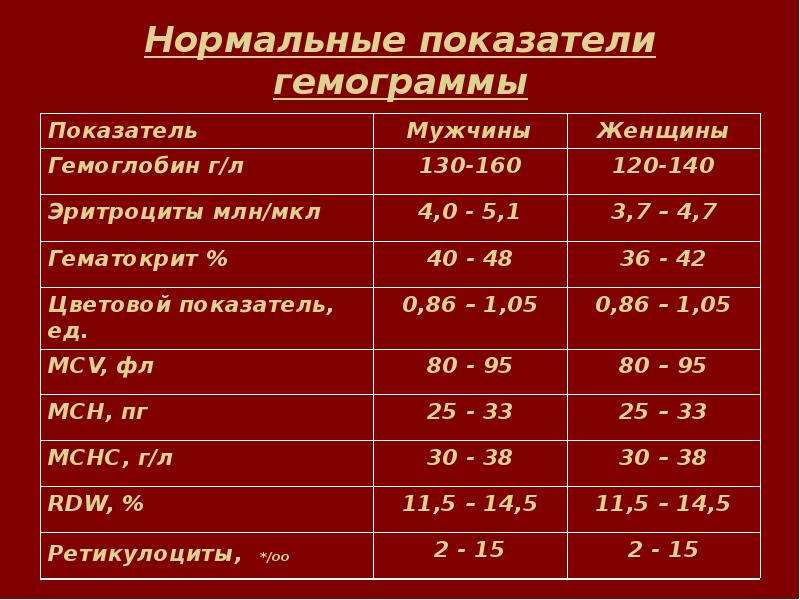 По данным Всемирной организации здравоохранения (ВОЗ), содержание гемоглобина в крови у мужчин выше, чем у женщин. По отдельной шкале медики оценивают уровень гемоглобина в крови беременных женщин. Нижней границей нормы в I и III триместре беременности считается 110 г/л, во II — 105 г/л. Верхнее значение на протяжении всего периода ожидания ребенка не должно превышать 155 г/л.
По данным Всемирной организации здравоохранения (ВОЗ), содержание гемоглобина в крови у мужчин выше, чем у женщин. По отдельной шкале медики оценивают уровень гемоглобина в крови беременных женщин. Нижней границей нормы в I и III триместре беременности считается 110 г/л, во II — 105 г/л. Верхнее значение на протяжении всего периода ожидания ребенка не должно превышать 155 г/л.
|
Пол |
Гемоглобин г/л (грамм на литр) |
|
Мужчины |
130-160 (нижний предел — 120, верхний предел — 180) |
|
Женщины |
120-150 |
|
Беременные женщины |
110-155 |
К чему приводит недостаток или избыток гемоглобина в крови человека?
Недостаток гемоглобина свидетельствует о таком заболевании, как анемия или малокровие. При наличии данного недуга организм человека испытывает кислородное голодание — гипоксию. Это выражается в общей слабости, доходящей до обмороков, дистрофических явлениях, отмирании тканей, которые не может напитать кислородом жидкая кровь. Порой анемия может выступать симптомом более серьезного заболевания или говорить о проблемах с желудочно-кишечным трактом. Основные причины понижения гемоглобина: дефицит меди и железа в продуктах питания, интенсивный рост организма, кровопотери или переливание крови.
При наличии данного недуга организм человека испытывает кислородное голодание — гипоксию. Это выражается в общей слабости, доходящей до обмороков, дистрофических явлениях, отмирании тканей, которые не может напитать кислородом жидкая кровь. Порой анемия может выступать симптомом более серьезного заболевания или говорить о проблемах с желудочно-кишечным трактом. Основные причины понижения гемоглобина: дефицит меди и железа в продуктах питания, интенсивный рост организма, кровопотери или переливание крови.
Как поднять себе уровень гемоглобина, читайте в справке >>
Повышенное содержание гемоглобина также может стать симптом многих достаточно серьезных заболеваний. Например, эритроцитоза — увеличения объема эритроцитов в крови, которое без лечения может вызывает её загустение. В этих случаях у человека развиваются тромбы, инсульты и инфаркты. Причинами повышения гемоглобина могут стать некоторые заболевания крови, легочная недостаточность, болезни сердца или обезвоживание организма.
Как узнать свой уровень гемоглобина?
Чтобы узнать в норме ли у вас гемоглобин, необходимо сдать общий анализ крови. Перед сдачей крови врачи не рекомендуют принимать пищу. Как минимум за 12 часов до посещения больницы нужно избегать активных физических нагрузок, а также ни в коем случае не употреблять алкоголь. Иначе результаты лабораторных исследований будут искажены.
Смотрите также:
Гемоглобин — одна из мишеней коронавируса COVID-19
Перевод на русский язык компании Logrus Global: https://logrusglobal.ru
COVID-19: атакует 1-бета-цепь гемоглобина и захватывает порфирин, чтобы ингибировать метаболизм человеческого гема.
14 апреля 2020 г.
Скачать PDF-версию
Авторы: Вэньчжун Лю1,2,*, Хуалань Ли2
1 Факультет информатики и инженерии, Сычуаньский инженерно-технический университет, Цзыгун, 643002, Китай;
2 Факультет медико-биологической и пищевой промышленности, Ибиньский университет, Ибинь, 644000, Китай;
* Адрес для переписки: liuwz@suse.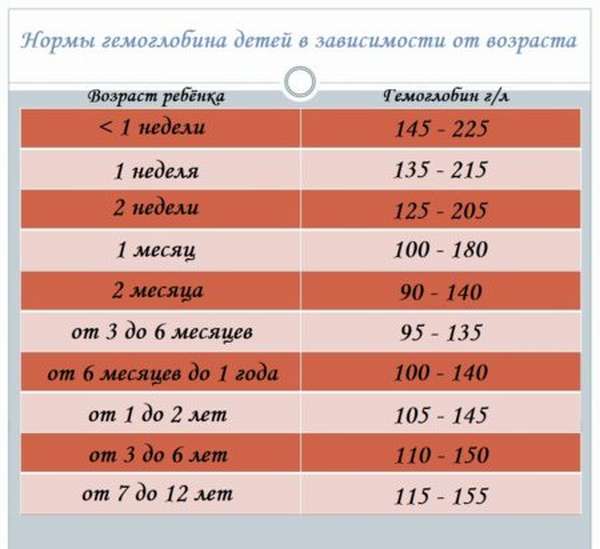 edu.cn.
edu.cn.
Конспект
Новая коронавирусная пневмония (COVID-19) представляет собой контагиозную острую респираторную инфекцию, вызванную новым коронавирусом. Этот вирус представляет собой РНК-вирус с позитивной полярностью цепи, имеющий высокую степень гомологии с коронавирусом летучей мыши. В этом исследовании для сравнения биологических ролей некоторых белков нового коронавируса использовали анализ консервативных доменов, гомологическое моделирование и молекулярную стыковку. Результаты показали, что белок ORF8 и поверхностный гликопротеин могут связываться с порфирином. В то же время белки orf1ab, ORF10 и ORF3a могут координированно атаковать гем, находящийся на 1-бета-цепи гемоглобина, что приводит к отщеплению железа с образованием порфирина. В результате такой атаки количество гемоглобина, который может переносить кислород и углекислый газ, становится все меньше и меньше. Клетки легких испытывают чрезвычайно сильное отравление и воспаление из-за невозможности обеспечения интенсивного обмена углекислым газом и кислородом; в конечном итоге изображения ткани легких принимают вид матового стекла.

Ключевые слова: новый коронавирус; респираторный дистресс; вид матового стекла изображения легкого; гликопротеин E2; ОRF8; оrf1ab; хлорохин; кровь; диабетический; флуоресцентный резонансный энергоперенос; древний вирус; цитокиновый шторм.
1. Введение
Новая коронавирусная пневмония (COVID-19) — контагиозное острое респираторное инфекционное заболевание. Пациенты с коронавирусной пневмонией страдают от лихорадки с температурой выше 38 градусов с такими симптомами, как сухой кашель, усталость, одышка, затрудненное дыхание, при визуализации легких они имеют вид матового стекла1-3. При морфологическом исследовании образцов тканей можно обнаружить большое количество слизи без явных вирусных включений. Эта пневмония была впервые обнаружена в декабре 2019 года на южнокитайском рынке морепродуктов провинции Хубэй, Китай4. Инфекция имеет высокую контагиозность5,6. Сейчас количество инфицированных людей достигло десятков тысяч по всему миру, и распространение инфекции не ограничено расой и границами. Исследователи провели тесты на выделение вирусов и секвенирование нуклеиновых кислот, чтобы подтвердить, что заболевание было вызвано новым коронавирусом7,8. Отмечено, что нуклеиновая кислота нового коронавируса представляет собой РНК с позитивной полярностью цепи8. Его структурные белки включают: белок-шип (S), белок оболочки (E), мембранный белок (M) и нуклеокапсидный фосфопротеин. Транскрибируемые неструктурные белки включают: orf1ab, ORF3a, ORF6, ORF7a, ORF10 и ORF8. Новый коронавирус высоко гомологичен коронавирусу летучих мышей9,10 и обладает значительной гомологией по отношению к вирусу SARS11,12. Исследователи изучили функцию структурных белков и некоторых неструктурных белков нового коронавируса13,14. Но новый коронавирус обладает целым набором потенциальных геномных характеристик, часть которых в основном является причиной вспышки в человеческой популяции15,16. Например, CoV EIC (белок оболочки коронавируса с функцией ионного канала) участвует в модулировании высвобождения вириона и взаимодействия «CoV — хозяин»17.
Исследователи провели тесты на выделение вирусов и секвенирование нуклеиновых кислот, чтобы подтвердить, что заболевание было вызвано новым коронавирусом7,8. Отмечено, что нуклеиновая кислота нового коронавируса представляет собой РНК с позитивной полярностью цепи8. Его структурные белки включают: белок-шип (S), белок оболочки (E), мембранный белок (M) и нуклеокапсидный фосфопротеин. Транскрибируемые неструктурные белки включают: orf1ab, ORF3a, ORF6, ORF7a, ORF10 и ORF8. Новый коронавирус высоко гомологичен коронавирусу летучих мышей9,10 и обладает значительной гомологией по отношению к вирусу SARS11,12. Исследователи изучили функцию структурных белков и некоторых неструктурных белков нового коронавируса13,14. Но новый коронавирус обладает целым набором потенциальных геномных характеристик, часть которых в основном является причиной вспышки в человеческой популяции15,16. Например, CoV EIC (белок оболочки коронавируса с функцией ионного канала) участвует в модулировании высвобождения вириона и взаимодействия «CoV — хозяин»17.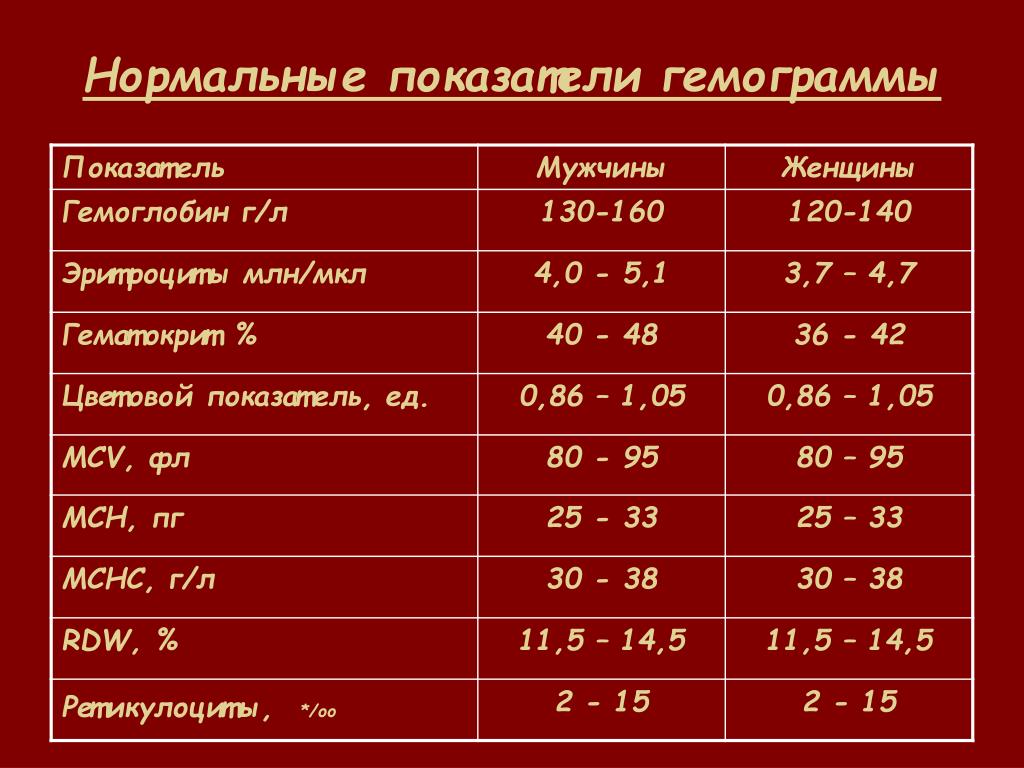 Шип-белки, белки ORF8 и ORF3a значительно отличаются от белков других известных SARS-подобных коронавирусов, и они могут вызывать более серьезные различия в патогенности и передаче по сравнению с известными для SARS-CoV18. Более ранние исследования показали, что новый коронавирус проникает в эпителиальные клетки с использованием шип-белка, взаимодействующего с рецепторным белком ACE2 человека на поверхности клетки, что и вызывает инфекцию у людей. Однако структурный анализ шип-белка (S) нового коронавируса показывает, что белок S лишь слабо связывается с рецептором ACE2 по сравнению с коронавирусом SARS19. Из-за ограничений существующих экспериментальных методов специфические функции вирусных белков, таких как ORF8 и поверхностный гликопротеин, до сих пор неясны. Механизм патогенности нового коронавируса остается загадочным20.
Шип-белки, белки ORF8 и ORF3a значительно отличаются от белков других известных SARS-подобных коронавирусов, и они могут вызывать более серьезные различия в патогенности и передаче по сравнению с известными для SARS-CoV18. Более ранние исследования показали, что новый коронавирус проникает в эпителиальные клетки с использованием шип-белка, взаимодействующего с рецепторным белком ACE2 человека на поверхности клетки, что и вызывает инфекцию у людей. Однако структурный анализ шип-белка (S) нового коронавируса показывает, что белок S лишь слабо связывается с рецептором ACE2 по сравнению с коронавирусом SARS19. Из-за ограничений существующих экспериментальных методов специфические функции вирусных белков, таких как ORF8 и поверхностный гликопротеин, до сих пор неясны. Механизм патогенности нового коронавируса остается загадочным20.
В литературе21 описаны показатели биохимического исследования 99 пациентов с вызванной новым коронавирусом пневмонией, и в этом отчете отражены аномалии связанных с гемоглобином показателей биохимических анализов пациентов. Согласно отчету, количество гемоглобина и нейтрофилов у большинства пациентов снизилось, а индексные значения сывороточного ферритина, скорости оседания эритроцитов, С-реактивного белка, альбумина и лактатдегидрогеназы у многих пациентов значительно возросли. Эти изменения предполагают, что содержание гемоглобина у пациента уменьшается, а гема увеличивается, и организм будет накапливать слишком много вредных ионов железа, что вызовет воспаление в организме и увеличит уровни С-реактивного белка и альбумина. Клетки реагируют на стресс, вызванный воспалением, производя большое количество сывороточного ферритина для связывания свободных ионов железа, чтобы уменьшить повреждения. Гемоглобин состоит из четырех субъединиц, 2-α и 2-β, и каждая субъединица имеет железосодержащий гем22,23. Гем является важным компонентом гемоглобина. Это порфирин, содержащий железо. Структура без железа называется порфирином. Когда железо находится в двухвалентном состоянии, гемоглобин может отщеплять углекислый газ и связывать атомы кислорода в альвеолярных клетках, при этом железо окисляется до трехвалентного уровня.
Согласно отчету, количество гемоглобина и нейтрофилов у большинства пациентов снизилось, а индексные значения сывороточного ферритина, скорости оседания эритроцитов, С-реактивного белка, альбумина и лактатдегидрогеназы у многих пациентов значительно возросли. Эти изменения предполагают, что содержание гемоглобина у пациента уменьшается, а гема увеличивается, и организм будет накапливать слишком много вредных ионов железа, что вызовет воспаление в организме и увеличит уровни С-реактивного белка и альбумина. Клетки реагируют на стресс, вызванный воспалением, производя большое количество сывороточного ферритина для связывания свободных ионов железа, чтобы уменьшить повреждения. Гемоглобин состоит из четырех субъединиц, 2-α и 2-β, и каждая субъединица имеет железосодержащий гем22,23. Гем является важным компонентом гемоглобина. Это порфирин, содержащий железо. Структура без железа называется порфирином. Когда железо находится в двухвалентном состоянии, гемоглобин может отщеплять углекислый газ и связывать атомы кислорода в альвеолярных клетках, при этом железо окисляется до трехвалентного уровня. Когда гемоглобин становится доступен другим клеткам организма через кровь, он может высвобождать атомы кислорода и присоединять углекислый газ, а железо восстанавливается до двухвалентного.
Когда гемоглобин становится доступен другим клеткам организма через кровь, он может высвобождать атомы кислорода и присоединять углекислый газ, а железо восстанавливается до двухвалентного.
Особо эффективных лекарств и вакцин для борьбы с болезнью, вызванной новым коронавирусом, не существует24. Однако в недавних поисках клинических методов лечения было обнаружено несколько старых препаратов, которые могут подавлять некоторые функции вируса, например, хлорохина фосфат оказывает определенное влияние на новую коронавирусную пневмонию25. Хлорохина фосфат — это противомалярийный препарат, который применяется в клинике уже более 70 лет. Эксперименты показывают, что эритроциты, инфицированные возбудителем малярии, могут накапливать большое количество хлорохина. Препарат приводит к потере фермента гемоглобина и смерти паразита из-за недостаточности аминокислот для его роста и развития. Предполагается, что терапевтический эффект хлорохина фосфата в отношении новой коронавирусной пневмонии может быть тесно связан с аномальным метаболизмом гемоглобина у человека. Между тем мы можем отметить, что хлорохин также широко используется для лечения порфирии26,27.
Между тем мы можем отметить, что хлорохин также широко используется для лечения порфирии26,27.
Поэтому мы предположили, что присоединение вирусных белков к порфиринам вызовет ряд патологических реакций у человека, таких как снижение уровня гемоглобина. Из-за тяжелой эпидемии и существующих условий с ограниченными экспериментальными методами тестирования функций белков большое научное значение имеет анализ функции белков нового коронавируса методами биоинформатики.
В этом исследовании для анализа функций белков, связанных с вирусом, использовались методы прогнозирования консервативных доменов, гомологического моделирования и молекулярной стыковки. Это исследование показало, что белок ORF8 и поверхностный гликопротеин способны объединяться с порфирином с образованием комплекса, в то время как белки orf1ab, ORF10, ORF3a скоординировано атакуют гем на 1-бета-цепи гемоглобина и отщепляют железо с образованием порфирина. Этот механизм вируса подавляет нормальный метаболический путь гема и приводит к проявлению у людей симптомов заболевания. Основываясь на результатах вышеупомянутых исследований, с помощью технологии молекулярной стыковки мы также проверили то, каким образом хлорохина фосфат и фавипиравир могут быть полезны в клинической практике.
Основываясь на результатах вышеупомянутых исследований, с помощью технологии молекулярной стыковки мы также проверили то, каким образом хлорохина фосфат и фавипиравир могут быть полезны в клинической практике.
2. Материалы и методы
2.1. Набор данных
Следующие последовательности белка загружали из NCBI: все белки нового коронавируса Ухань, гем-связывающий белок; гемоксидаза; для анализа консервативного домена использовали белковые последовательности.
Все белки нового коронавируса Ухань также использовали для конструирования трехмерных структур путем гомологического моделирования.
В то же время следующие файлы были загружены из базы данных PDB: кристаллическая структура MERS-CoV nsp10_nsp16 комплекс—5yn5, гем, оксигемоглобин человека 6bb5; дезоксигемоглобин человека 1a3n; 0TX; Rp. Комплекс MERS-CoV nsp10_nsp16—5yn5 использовался для гомологического моделирования. Гем, 0TX и 1RP использовались для молекулярной стыковки. Два оксигемоглобин был использован для стыковки белков.
2.2. Блок-схема биоинформационного анализа
На основе опубликованных в данном исследовании биологических белковых последовательностей была проведена серия биоинформационного анализа. Этапы показаны на рисунке 1:1. Консервативные домены вирусных белков анализируются28-30 онлайн-сервером МЕМЕ. Консервативные домены использовались для прогнозирования функциональных различий вирусных белков и белков человека. 2. Трехмерная структура вирусных белков была построена путем гомологического моделирования в средстве Swiss-model31,32. Если длина последовательности превышала 5000 нуклеотидов, использовался инструмент гомологического моделирования Discovery-Studio 2016. 3. Использование технологии молекулярной стыковки (инструмент LibDock) Discovery-Studio 201633 позволило смоделировать рецептор-лигандное соединение вирусных белков с гемом человека (или порфиринами). С учетом результатов биоинформационного анализа была построена модель жизненного цикла вируса и предложена соответствующая молекулярная картина заболевания.
Рисунок 1. Блок-схема биоинформационного анализа.
Рабочий процесс основан на эволюционных принципах. Хотя биологическая последовательность, характерная для развитых форм жизни и вируса, отличается, молекулы с аналогичными структурами всегда могут играть аналогичные биологические роли. В методе гомологического моделирования используется принцип, согласно которому аналогичная первичная структура белковых последовательностей имеет аналогичную пространственную структуру. Метод молекулярной стыковки построен на гомологическом моделировании реальных трехмерных молекул.
2.3. Анализ консервативного домена
MEME Suite — это онлайн-сайт, который объединяет множество инструментов прогнозирования и описания мотивов. Алгоритм максимального ожидания (EM) является основой для идентификации мотива на сайте MEMЕ. Мотив представляет собой консервативный домен небольшой последовательности в белке. Модели, основанные на мотивах, помогают оценить надежность филогенетического анализа. После открытия онлайн-инструмента MEME интересующие белковые последовательности объединяют в текстовый файл, при этом сохраняется формат файла .fasta. Затем выбирают нужное количество мотивов и нажимают кнопку «Перейти». В конце анализа консервативные домены отображаются после нажатия на ссылку.
После открытия онлайн-инструмента MEME интересующие белковые последовательности объединяют в текстовый файл, при этом сохраняется формат файла .fasta. Затем выбирают нужное количество мотивов и нажимают кнопку «Перейти». В конце анализа консервативные домены отображаются после нажатия на ссылку.
2.4. Гомологическое моделирование
SWISS-MODEL — это полностью автоматический сервер гомологического моделирования структуры белка, доступ к которому можно получить через веб-сервер. Первый шаг — войти на сервер SWISS-MODEL, ввести последовательность и нажать Search Template («Поиск шаблона»), чтобы выполнить простой поиск шаблона. После завершения поиска можно выбрать шаблон для моделирования. Поиск шаблонов выполняется нажатием кнопки Build Model, и модель шаблона выбирается автоматически. Как видно, было найдено несколько шаблонов, а затем построено множество моделей. Здесь выбирается только модель. Модель в формате PDB загружается и визуализируется в VMD. SWISS-MODEL моделирует только белковые модели, соответствующие последовательностям менее 5000 нуклеотидных оснований.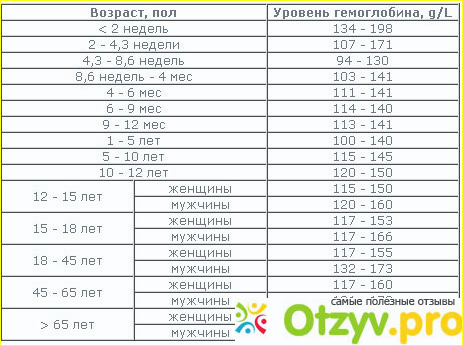 Для моделирования белка, соответствующего последовательности более 5000 нуклеотидов, можно использовать инструмент гомологического моделирования Discovery-Studio.
Для моделирования белка, соответствующего последовательности более 5000 нуклеотидов, можно использовать инструмент гомологического моделирования Discovery-Studio.
Перед использованием Discovery-Studio для гомологического моделирования неизвестного белка (такого, как orf1ab) файл структуры pdb матричного белка, такого как MERS-CoVnsp10_nsp16 комплекс 5yn5, должен быть загружен из базы данных PDB. Затем для сопоставления гомологичных последовательностей белков 5yn5 и orf1ab был применен инструмент сопоставления последовательностей Discovery-Studio. Затем был построен файл пространственной структуры orf1ab на основе матричного белка 5yn5.
2.5. Технология молекулярной стыковки
Молекулярная стыковка — это процесс нахождения наилучшего соответствия между двумя или более молекулами посредством определения геометрического и энергетического соответствия. Этапы использования молекулярной стыковки LibDock с Discovery-Studio следующие:
1. Подготовка модели лиганда. Откройте файл лиганда, например, гема, и нажмите кнопку Prepare Ligands («Подготовка лигандов») в подменю Dock Ligands («Док-лиганды») меню Receptor-Ligand Interactions («Взаимодействие рецептор-лиганд»), чтобы создать модель лиганда гема для стыковки. Сначала удалите FE (атом железа) из гема, а затем нажмите кнопку Prepare Ligands («Подготовка лигандов»), после чего будет сгенерирована модель лиганда порфирина. При открытии 0 XT снова нажмите кнопку Prepare Ligands («Подготовка лигандов»), чтобы получить модель хлорохинового лиганда.
Откройте файл лиганда, например, гема, и нажмите кнопку Prepare Ligands («Подготовка лигандов») в подменю Dock Ligands («Док-лиганды») меню Receptor-Ligand Interactions («Взаимодействие рецептор-лиганд»), чтобы создать модель лиганда гема для стыковки. Сначала удалите FE (атом железа) из гема, а затем нажмите кнопку Prepare Ligands («Подготовка лигандов»), после чего будет сгенерирована модель лиганда порфирина. При открытии 0 XT снова нажмите кнопку Prepare Ligands («Подготовка лигандов»), чтобы получить модель хлорохинового лиганда.
2. Подготовьте модель белкового рецептора. Откройте файл pdb белка (сгенерированный с помощью гомологического моделирования) и нажмите Prepare protein («Подготовка белка») в подменю Dock Ligands («Док-лиганды») меню Receptor-Ligand Interactions («Взаимодействие рецептора с лигандом»), чтобы создать модель рецептора белка для стыковки.
3. Установите параметры стыковки для ее достижения. Выберите модель генерируемого белкового рецептора. В подменю Define and Edit Binding Site («Определение и редактирование сайта связывания») в меню Receptor-Ligand Interactions («Взаимодействие рецептора с лигандом») нажмите кнопку From receptor Cavities («Из полостей рецептора»). На диаграмме модели рецептора белка появляется красная сфера. После щелчка правой кнопкой мыши по красному шару можно изменить его радиус. Затем в меню Receptor-Ligand Interactions («Взаимодействие рецептора с лигандом») выберите Dock Ligands (LibDock) («Док-лиганды LibDock») в подменю Dock Ligands («Док-лиганды»). Во всплывающем окне выберите лиганд в качестве вновь созданной модели лиганда (ALL) и выберите рецептор в качестве вновь созданной модели рецептора (ALL), а для сфер сайтов задайте только что установленные координаты сфер. Наконец, нажмите RUN («Выполнить»), чтобы начать стыковку.
На диаграмме модели рецептора белка появляется красная сфера. После щелчка правой кнопкой мыши по красному шару можно изменить его радиус. Затем в меню Receptor-Ligand Interactions («Взаимодействие рецептора с лигандом») выберите Dock Ligands (LibDock) («Док-лиганды LibDock») в подменю Dock Ligands («Док-лиганды»). Во всплывающем окне выберите лиганд в качестве вновь созданной модели лиганда (ALL) и выберите рецептор в качестве вновь созданной модели рецептора (ALL), а для сфер сайтов задайте только что установленные координаты сфер. Наконец, нажмите RUN («Выполнить»), чтобы начать стыковку.
4. Рассчитайте энергию связывания и выберите положение с наибольшей энергией связывания. После завершения стыковки будет отображено множество местоположений лиганда. Откройте окно стыковки и нажмите кнопку Caculate Binding Energies («Рассчитать энергии связывания») в подменю Dock Ligands («Док-лиганды») меню Receptor-Ligand Interactions («Взаимодействие рецептора с лигандом»). Во всплывающем окне выберите рецептор в качестве значения по умолчанию, лиганд в качестве стыкуемой модели (ALL), а затем запустите вычисление энергии связывания. Наконец, сравните энергию связывания и выберите положение с наибольшей энергией связывания. Чем выше стабильность комплекса, тем больше энергия связывания.
Наконец, сравните энергию связывания и выберите положение с наибольшей энергией связывания. Чем выше стабильность комплекса, тем больше энергия связывания.
5. Экспортируйте вид совместного сечения. Для вида в состоянии стыковки после установки стиля отображения области связывания нажмите кнопку Show 2D Map («Показать 2D-карту») в подменю View Interaction («Просмотр взаимодействия») меню Receptor-Ligand Interaction («Взаимодействие между рецептором и лигандом»), чтобы открыть вид участка связывания. Это представление может быть сохранено в виде файла изображения.
2.6. Технология стыковки белков
ZDOCK от Discovery-Studio — это еще один инструмент молекулярной стыковки для изучения взаимодействий белков. Мы использовали его для изучения атаки гемоглобина вирусными неструктурными белками. Ниже приведено описание стыковки orf1ab и гемоглобина, при изучении стыковки с другими неструктурными белками вируса применяли аналогичные методы стыковки. После открытия PBD-файлов человеческого оксигемоглобина 6bb5 и белка orf1ab нажмите кнопку Dock proteins (ZDOCK) в меню Dock and Analyze Protein Comlexes («Стыковка и анализ белковых комплексов»). Во всплывающем интерфейсе выберите человеческий оксигемоглобин 6bb5 в качестве рецептора, а orf1a в качестве лиганда, а затем нажмите кнопку Run («Выполнить»). После того как компьютер закончит вычисления, нажмите на интерфейс proteinpose («положение белка») и выберите положение и кластер с самым высоким баллом ZDOCK. Так можно получить положение белка orf1ab на человеческом оксигемоглобине 6bb5. Дезоксигемоглобин человека 1a3n имеет сходную схему стыковки с белком orf1ab.
Во всплывающем интерфейсе выберите человеческий оксигемоглобин 6bb5 в качестве рецептора, а orf1a в качестве лиганда, а затем нажмите кнопку Run («Выполнить»). После того как компьютер закончит вычисления, нажмите на интерфейс proteinpose («положение белка») и выберите положение и кластер с самым высоким баллом ZDOCK. Так можно получить положение белка orf1ab на человеческом оксигемоглобине 6bb5. Дезоксигемоглобин человека 1a3n имеет сходную схему стыковки с белком orf1ab.
3. РЕЗУЛЬТАТЫ
3.1. Вирусные структурные белки, связывающие порфирин
У человека гемоглобин может разлагаться на глобин и гем. Гем состоит из порфирина и иона железа, при этом ион железа находится в середине порфирина. Гем нерастворим в воде и может быть объединен с гем-связывающими белками с образованием комплекса и транспортироваться в печень. Порфирин разлагается до билирубина и выводится через желчный проток, а железо, содержащееся в молекуле, может повторно использоваться организмом. Если вирусные белки могут связываться с порфирином гема, они должны обладать связывающей способностью, аналогичной гем-связывающему белку человека, то есть вирусные белки и гем-связывающие белки должны иметь аналогичные консервативные домены. Для изучения связывания структурных белков вируса и порфирина в настоящей работе были применены следующие методы биоинформатики.
Если вирусные белки могут связываться с порфирином гема, они должны обладать связывающей способностью, аналогичной гем-связывающему белку человека, то есть вирусные белки и гем-связывающие белки должны иметь аналогичные консервативные домены. Для изучения связывания структурных белков вируса и порфирина в настоящей работе были применены следующие методы биоинформатики.
Сначала на онлайн-сервере MEME был выполнен поиск консервативных доменов в каждом структурном белке вируса и человеческом гем-связывающем белке (ID:NP_057071.2 гем-связывающий белок 1, ID: EAW47917.1 гем-связывающий белок 2). На рисунке 2 показано, что три вирусных белка (поверхностный гликопротеин, белок оболочки и нуклеокапсидный фосфопротеин) и белки связывания гема имеют консервативные домены, но мембранный гликопротеин не имеет консервативных доменов, p-значения малы, различия статистически значимы. Домены в трех вирусных белках различны, что позволяет предположить некоторое различие способностей структурных белков связывать порфирин. Мембранный гликопротеин не может связываться с порфирином.
Мембранный гликопротеин не может связываться с порфирином.
Рисунок 2. Консервативные домены в структурных белках и гем-связывающих белках человека. A. Консервативные домены поверхностного гликопротеина. B. Консервативные домены белка оболочки. C. Консервативные домены мембранного гликопротеина. D. Консервативные домены нуклеокапсидного фосфопротеина.
Затем онлайн-сервер Swiss-model смоделировал поверхностные гликопротеины для получения трехмерной структуры, и были выбраны два вида файлов на основе шаблонов Spike и E2. 3D-структурный файл гема был загружен из базы данных PDB.
В конце концов Discovery-Studio реализовала молекулярную стыковку поверхностных гликопротеинов и порфирина. Сначала не удалось стыковать белок-шип с гемом (и порфирином). Гликопротеин Е2 (рисунок 3.A) получен из матриц 1zva.1.A. Стыковка гликопротеина Е2 и гема также была безуспешной. Когда удалили ион железа и гем стал порфирином, удалось выполнить множество видов стыковки между гликопротеином E2 и порфирином.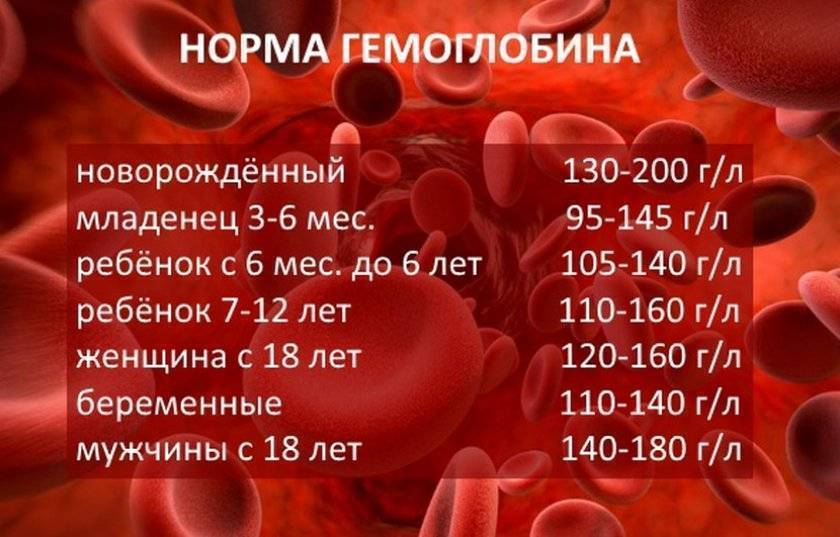 После вычисления энергии связывания за результат была принята позиция стыковки с самой высокой энергией связывания (7 530 186 265,80 ккал/моль). Результат стыковки показан на рисунке 4.A-1, где представлена молекулярная модель связывания гликопротеина E2 с порфирином.
После вычисления энергии связывания за результат была принята позиция стыковки с самой высокой энергией связывания (7 530 186 265,80 ккал/моль). Результат стыковки показан на рисунке 4.A-1, где представлена молекулярная модель связывания гликопротеина E2 с порфирином.
На рисунке 4.A-2 представлен двухмерный вид участка связывания, в котором 18 аминокислот гликопротеина Е2 взаимодействуют с порфирином.
При анализе белка оболочки использовались те же методы. Шаблон 5×29.1. A был выбран в качестве шаблона 3D-структуры белка оболочки (рисунок 3.B). Discovery-Studio обнаружила несколько видов стыковки белка оболочки и порфирина, где было выбрано положение стыковки с самой высокой энергией связывания (219 317,76 ккал/моль). На рисунке 4.B-1 показан результат стыковки, представляющий собой молекулярную модель связывания белка оболочки с порфирином. Рисунок 4.В-2 представляет двухмерный вид участка связывания, в котором 18 аминокислот белка оболочки взаимодействуют с порфирином.
Те же методы использовались для анализа нуклеокапсидного фосфопротеина. В качестве шаблона фосфопротеина нуклеокапсида использовали 1ssk.1.А (рисунок 3.С). Discovery-Studio позволила выявить вариант стыковки между нуклеокапсидным фосфопротеином и порфирином с самой высокой энергией связывания (15 532 506,53 ккал/моль). На рисунке 4.С-1 показан результат стыковки, представляющий собой молекулярную модель связывания нуклеокапсидного фосфопротеина с порфирином. На рисунке 4.C-2 представлен двухмерный вид связывающего участка, где 22 аминокислоты нуклеокапсидного фосфопротеина связаны с порфирином. Мембранный белок получен из шаблонов 1zva.1.A. Состыковать мембранный белок с гемом (и порфирином) не удалось. Полученные результаты свидетельствуют, что поверхностный гликопротеин, белок оболочки и нуклеокапсидный фосфопротеин могут связываться с порфирином с образованием комплекса.
Было обнаружено, что энергия связывания белка оболочки была самой низкой, энергия связывания гликопротеина Е2 была самой высокой, а энергия связывания нуклеокапсидного фосфопротеина была средней.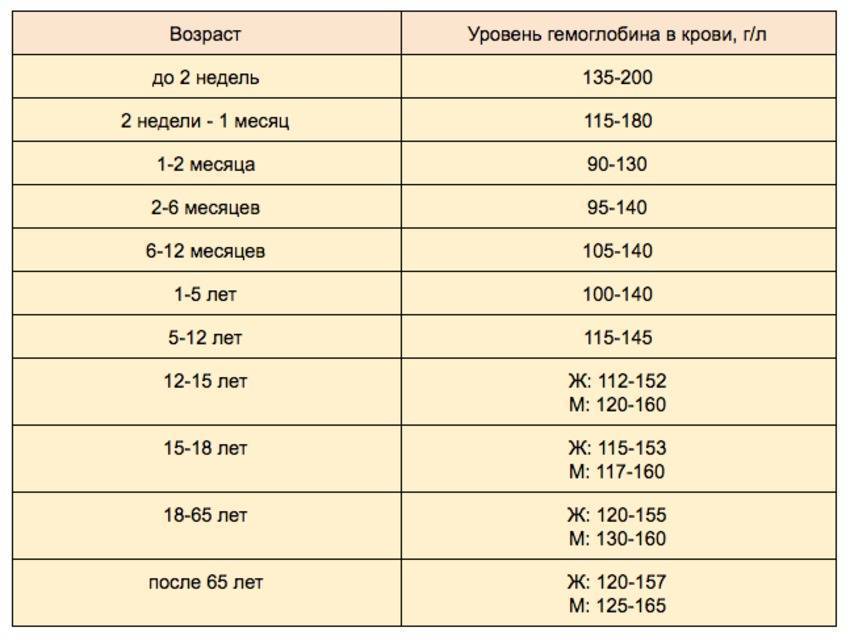 Это означает, что связывание гликопротеина Е2 с порфирином является наиболее стабильным, связывание нуклеокапсидного фосфопротеина с порфирином является неустойчивым, а связывание белка оболочки с порфирином является наиболее неустойчивым.
Это означает, что связывание гликопротеина Е2 с порфирином является наиболее стабильным, связывание нуклеокапсидного фосфопротеина с порфирином является неустойчивым, а связывание белка оболочки с порфирином является наиболее неустойчивым.
После этого был проведен следующий анализ, чтобы выяснить, атакуют ли структурные белки гем с отщеплением атома железа и образованием порфиринов. Гем имеет оксидазу, называемую гемоксидазой, которая окисляет гем и отщепляет ион железа. Если структурные белки могут атаковать гем и отщеплять ионы железа, они должны иметь такой же консервативный домен, как гемоксидаза. Онлайн-сервер MEME был использован для поиска консервативных доменов структурных белков и белков гемоксидазы (NP_002124.1: гемоксигеназы-1; BAA04789.1: гемоксигеназы-2; AAB22110.2: гемоксигеназы-2). В результате консервативных доменов структурных белков обнаружено не было (рисунок 5). Объединяя этот результат с результатом предыдущего анализа, можно предположить, что структурные белки могут объединяться только с порфирином.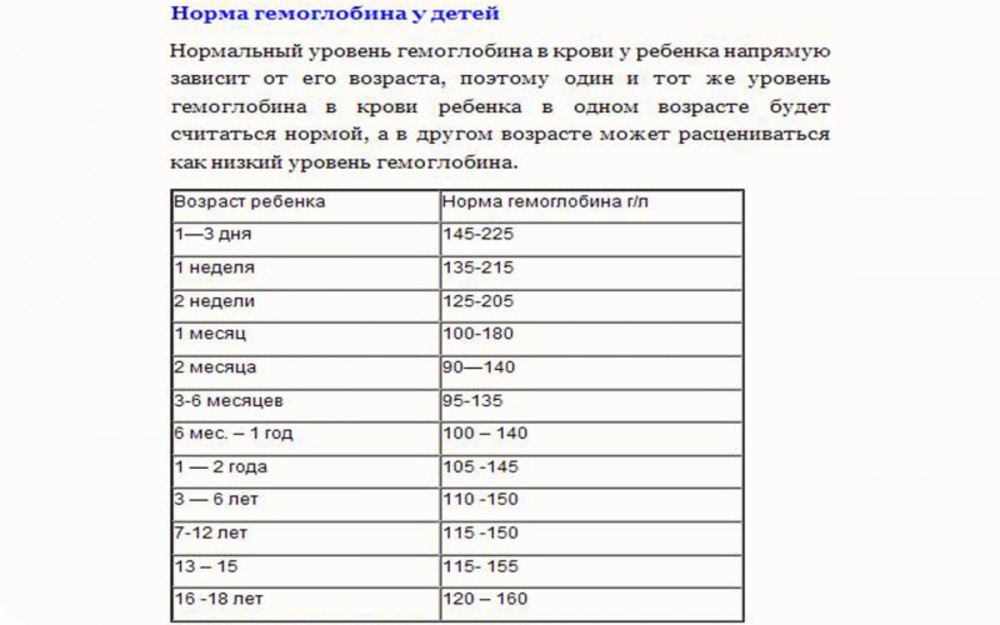 Можно сделать вывод, что структурные белки не атакуют гем, вызывая диссоциацию атома железа с образованием порфирина.
Можно сделать вывод, что структурные белки не атакуют гем, вызывая диссоциацию атома железа с образованием порфирина.
Рисунок 3. Трехмерные структурные схемы новых белков коронавируса, полученные с помощью гомологического моделирования. A. Гликопротеин E2 поверхностного гликопротеина. B. Белок оболочки. C. Нуклеокапсидный фосфопротеин. D. Белок orf1ab. E. Белок ORF8. F. Белок ORF7a.
Рисунок 4. Результаты молекулярной стыковки структурных белков вируса и порфирина (красная структура). A. Результаты молекулярной стыковки гликопротеина E2 и порфирина. B. Результаты молекулярной стыковки белка оболочки и порфирина. C. Результаты молекулярной стыковки нуклеокапсидного фосфопротеина и порфирина. 1. Структурные белки вируса. 2. Вид участков связывания.
Рисунок 5. Консервативные домены структурных белков и белков гемоксигеназы человека. A. Консервативные домены поверхностного гликопротеина. B. Консервативные домены белка оболочки. C. Консервативные домены мембраны. D. Консервативные домены нуклеокапсидного фосфопротеина.
A. Консервативные домены поверхностного гликопротеина. B. Консервативные домены белка оболочки. C. Консервативные домены мембраны. D. Консервативные домены нуклеокапсидного фосфопротеина.
3.2. Неструктурные белки вируса, связывающие порфирин
Сначала на онлайн-сервере MEME был выполнен поиск консервативных доменов в каждом структурном белке вируса и человеческом гем-связывающем белке (ID:NP_057071.2 гем-связывающий белок 1, ID: EAW47917.1 гем-связывающий белок 2). На рисунке 2 показано, что три вирусных белка (поверхностный гликопротеин, белок оболочки и нуклеокапсидный фосфопротеин) и белки связывания гема имеют консервативные домены, но мембранный гликопротеин не имеет консервативных доменов, p-значения малы, различия статистически значимы. Домены в трех вирусных белках различны, что позволяет предположить некоторое различие способностей структурных белков связывать порфирин. Мембранный гликопротеин не может связываться с порфирином.
Рисунок 6. Консервативные домены в неструктурных белках и гем-связывающих белках человека. A. Консервативные домены orf1ab. B. Консервативные домены ORF3a. C. Консервативные домены ORF6. D. Консервативные домены ORF7a. E. Консервативные домены ORF8. F. Консервативные домены ORF10.
Гомологическое моделирование и технология молекулярной стыковки были применены для изучения способности белка orf1ab связывать гем. Поскольку Swiss-model не может моделировать 3D-структуру белковой последовательности orf1ab из-за ограничения на длину кодирующей последовательности (не более 5000 нуклеотидов), для гомологического моделирования использовалась программа Discovery-Studio. Кристаллическая структура комплекса MERS-CoV nsp10_nsp16 5yn5 и гема была загружена из базы данных PDB. В этом исследовании кристаллическая структура комплекса MERS-CoV nsp10_nsp16 5yn5 была взята в качестве матрицы для создания гомологичной структуры белка orf1ab. В качестве 3D-структуры белка orf1ab была выбрана гомологичная структура по умолчанию (рисунок 3.D). Затем в программе Discovery-Studio была проведена молекулярная стыковка белка orf1ab и порфирина. Белок orf1ab и гем не удалось состыковать, но после удаления ионов железа и превращения гема в порфирин радиус действия увеличился и несколько типов стыковки удалось довести до конца. Путем вычисления энергии связывания была выбрана модель стыковки с наибольшей энергией связывания (561 571,10 ккал/моль). Результат стыковки показан на рисунке 7.A-1, где представлена молекулярная модель связывания белка orf1ab с порфирином. Связывающая часть белка orf1ab действует как зажим. Именно этот зажим захватывает порфирин без иона железа. На рисунке 7.A-2 показан двухмерный вид участка связывания. Видно, что 18 аминокислот белка orf1ab связаны с порфирином.
В качестве 3D-структуры белка orf1ab была выбрана гомологичная структура по умолчанию (рисунок 3.D). Затем в программе Discovery-Studio была проведена молекулярная стыковка белка orf1ab и порфирина. Белок orf1ab и гем не удалось состыковать, но после удаления ионов железа и превращения гема в порфирин радиус действия увеличился и несколько типов стыковки удалось довести до конца. Путем вычисления энергии связывания была выбрана модель стыковки с наибольшей энергией связывания (561 571,10 ккал/моль). Результат стыковки показан на рисунке 7.A-1, где представлена молекулярная модель связывания белка orf1ab с порфирином. Связывающая часть белка orf1ab действует как зажим. Именно этот зажим захватывает порфирин без иона железа. На рисунке 7.A-2 показан двухмерный вид участка связывания. Видно, что 18 аминокислот белка orf1ab связаны с порфирином.
Для изучения свойств связывания белка ORF8 с гемом использовались те же этапы анализа, что и для структурного белкового метода. Файл структуры был создан на основе шаблона ORF7 (рисунок 3. E). Было обнаружено несколько видов стыковки белка ORF8 и порфирина, из которых выбрано стыковочное положение, имеющее наибольшую энергию связывания (12 804 859,25 ккал/моль). Результат стыковки (рисунок 7.В-1) представляет собой молекулярную модель связывания белка ORF8 с порфирином. Рисунок 7.В-2 представляет собой двухмерный вид участка связывания, где 18 аминокислот ORF8 связаны с порфирином.
E). Было обнаружено несколько видов стыковки белка ORF8 и порфирина, из которых выбрано стыковочное положение, имеющее наибольшую энергию связывания (12 804 859,25 ккал/моль). Результат стыковки (рисунок 7.В-1) представляет собой молекулярную модель связывания белка ORF8 с порфирином. Рисунок 7.В-2 представляет собой двухмерный вид участка связывания, где 18 аминокислот ORF8 связаны с порфирином.
Для анализа белка ORF7a использовались те же методы, что и при анализе белка ORF8. Шаблон ORF7a — 1yo4.1.A (рис. 3.F). Белок ORF7a и порфирин имели наивысшую энергию связывания (37 123,79 ккал/моль). На рисунке 7.С-1 показана молекулярная модель связи ORF7a с порфирином. Пятнадцать аминокислот ORF7a связаны с порфирином (рис. 7.C-2). Связывающая часть белка ORF7a также действует как зажим.
Swiss-модель не может предоставить шаблон для ORF10. ORF6a и ORF3a получены из шаблонов 3h08.1.A и 2m6n.1.A соответственно, но состыковать ORF6a (ORF3a) с гемом и порфирином не удалось.
Рисунок 7. Результаты молекулярной стыковки неструктурных белков вируса и порфирина (красный). A. Результаты молекулярной стыковки белка orf1ab и порфирина. B. Результаты молекулярной стыковки для белка ORF8 и порфирина. C. Результаты молекулярной стыковки белка ORF7a и порфирина. 1. Неструктурные белки вируса. 2. Вид участков связывания.
Результаты молекулярной стыковки неструктурных белков вируса и порфирина (красный). A. Результаты молекулярной стыковки белка orf1ab и порфирина. B. Результаты молекулярной стыковки для белка ORF8 и порфирина. C. Результаты молекулярной стыковки белка ORF7a и порфирина. 1. Неструктурные белки вируса. 2. Вид участков связывания.
Наконец, был проведен следующий анализ, чтобы выяснить, могли ли неструктурные белки атаковать гем и отщеплять атом железа с образованием порфиринов. Здесь для анализа консервативных доменов неструктурных белков и белков гемоксидазы использовался тот же метод, что и для предыдущего структурного белка — онлайн-сервер MEME (NP_002124.1: гемоксигеназа-1; BAA04789.1: гемоксигеназа-2; AAB22110.2: гемоксигеназа-2). Как показано на рисунке 8, ORF10, orf1ab и ORF3a имеют консервативные домены. Учитывая результаты предыдущего анализа, можно сказать, что неструктурные белки ORF10, orf1ab и ORF3a могут атаковать гем и отщеплять атом железа с образованием порфирина. Однако р-значение для orf1ab и ORF3a больше, чем 0,1 %. Поэтому ORF10 может быть основным белком, атакующим гем, тогда как orf1ab и ORF3a захватывают гем или порфирин.
Результаты показали, что orf1ab, ORF7a и ORF8 могут связываться с порфирином, в то время как ORF10, ORF3a и ORF6 не могут связываться с гемом (и порфирином). ORF10, ORF1ab и ORF3a также обладают способностью атаковать гем с образованием порфирина. Энергии связывания orf1ab, ORF7a, ORF8 и порфирина сравнивали между собой. Было обнаружено, что энергия связывания ORF7a была самой низкой, энергия связывания ORF8 была самой высокой, а энергия связывания orf1ab была средней. Это означает, что связывание ORF8 с порфирином является наиболее стабильным, связывание orf1ab с порфирином является неустойчивым, а связывание ORF7a с порфирином является наиболее неустойчивым. Последовательности ORF10 и ORF6 короткие, поэтому они должны быть короткими сигнальными пептидами. Следовательно, механизм, с помощью которого неструктурные белки атакуют гем, может быть такой: ORF10, ORF1ab и ORF3a атакуют гем и образуют порфирин; ORF6 и ORF7a отправляют порфирин в ORF8; и ORF8 и порфирин образуют стабильный комплекс.
Рисунок 8. Консервативные домены неструктурных белков и белков гемоксигеназы человека. A. Консервативные домены orf1ab. B. Консервативные домены ORF3a. C. Консервативные домены ORF6. D. Консервативные домены ORF7a. E. Консервативные домены ORF8. F. Консервативные домены ORF10.
3.3. Вирусный неструктурный белок атакует гем на бета-цепи гемоглобина
Порфирины в организме человека — это в основном железосодержащие порфирины, то есть гем. Большая часть молекул гема не свободна, а связана в составе гемоглобина. Для выживания вирусов им требуется большое количество порфиринов. Поэтому новый коронавирус нацелен на гемоглобин, атакует гем и охотится на порфирины. Результаты предыдущего анализа показали, что ORF1ab, ORF3a и ORF10 имеют домены, сходные с гемоксигеназой, но только ORF1ab может связываться с порфирином. Чтобы изучить атакующее поведение белков orf1ab, ORF3a и ORF10, мы использовали технологию молекулярной стыковки ZDOCK. Технология молекулярной стыковки ZDOCK позволяет анализировать взаимодействия белков и находить приблизительные положения этих трех белков на гемоглобине.
Технология молекулярной стыковки ZDOCK позволяет анализировать взаимодействия белков и находить приблизительные положения этих трех белков на гемоглобине.
Сначала мы загрузили гемоксигеназу 2 (5UC8) из PDB и использовали ее в качестве шаблона, а затем использовали инструмент гомологического моделирования Discovery-Studio для создания трехмерной структуры ORF10 (рисунок 9). Поскольку гемоглобин имеет две формы: окисленную и восстановленную, в приведенном ниже анализе выполнена молекулярная стыковка белков в этих двух случаях, а в качестве результата принята позиция с наивысшей оценкой ZDOCK.
Рисунок 9. Моделирование гомологии ORF10.
На дезоксигемоглобине orf1ab располагается в нижне-среднем участке 1-альфа- и 2-альфа-цепи вблизи 2-альфа-цепи (рисунок 10.A). ORF3a располагается в нижне-среднем участке 1-альфа и 2-альфа-цепи вблизи 2-альфа цепи (рисунок 10.B). ORF10 располагается в нижне-средней части 1-бета- и 2-бета-цепи вблизи 1-бета-цепи (рисунок 10.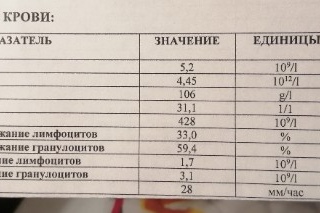 C). Предполагается следующий механизм: orf1ab атакует 2-альфа-цепь, вызывая изменения конформации белка глобина. Связывание ORF3A с цепью 2-альфа приводит к атаке ею цепи 1-бета, открывающей гем. ORF10 быстро присоединяется к 1-бета-цепи и непосредственно воздействует на гем 1-бета-цепи. Когда атом железа отщепляется, гем превращается в порфирин, и orf1ab получает возможность захватить порфирин. Белок orf1ab играет критически важную роль на протяжении всей атаки.
C). Предполагается следующий механизм: orf1ab атакует 2-альфа-цепь, вызывая изменения конформации белка глобина. Связывание ORF3A с цепью 2-альфа приводит к атаке ею цепи 1-бета, открывающей гем. ORF10 быстро присоединяется к 1-бета-цепи и непосредственно воздействует на гем 1-бета-цепи. Когда атом железа отщепляется, гем превращается в порфирин, и orf1ab получает возможность захватить порфирин. Белок orf1ab играет критически важную роль на протяжении всей атаки.
Рисунок 10. Вирусный неструктурный белок атакует гемоглобин. A. orf1ab атакует дезоксигемоглобин. B. ORF3a атакует дезоксигемоглобин. C. ORF10 атакует дезоксигемоглобин. D. orf1ab атакует окисленный гемоглобин. E. ORF10 атакует окисленный гемоглобин. F. ORF3a атакует окисленный гемоглобин.
На окисленном гемоглобине orf1ab располагается в нижне-средней части альфа- и бета-цепи вблизи альфа-цепи (рисунок 10.A). ORF10 располагается в нижней части бета-цепи, ближе к внешней (рисунок 10.B). ORF3a располагается в нижне-средней части альфа- и бета-цепи и приближен к бета-цепи (рисунок 10.C). Возможный механизм состоит в том, что orf1ab связывается с альфа-цепью и атакует бета-цепь, вызывая конфигурационные изменения в альфа- и бета-цепях; ORF3 атакует бета-цепь и обнажает гем. ORF10 быстро прикрепляется к бета-цепи и непосредственно влияет на атомы железа в геме бета-цепи. Гем после отщепления железа превращается в порфирин, и orf1ab получает возможность захватить порфирин. Белок orf1ab играет ключевую роль на протяжении всей атаки.
Атака вирусных белков на оксигемоглобин приводит к прогрессирующему уменьшению количества гемоглобина, который может переносить кислород. Влияние вирусных белков на дезоксигемоглобин будет еще сильнее уменьшать количество гемоглобина, доступного для переноса диоксида углерода и глюкозы крови. Люди с диабетом могут иметь нестабильный уровень глюкозы крови. Состояние пациента дополнительно ухудшается от отравления диоксидом углерода. Клетки легких испытывают чрезвычайно сильное воспаление из-за невозможности обеспечения интенсивного обмена углекислым газом и кислородом; в конечном итоге изображения ткани легких принимают вид матового стекла. Состояние пациентов с респираторными расстройствами ухудшится.
3.4. Валидация воздействия хлорохина фосфата
Химические компоненты хлорохина фосфата конкурируют с порфирином и связываются с вирусным белком, тем самым ингибируя атаку вирусного белка на гем или связывание с порфирином. Для проверки влияния хлорохина фосфата на молекулярный механизм действия вируса была принята технология молекулярной стыковки. Структурный файл 0TX (хлорохин) был загружен из базы данных PDB. Затем была использована технология молекулярной стыковки Discovery-Studio 2016 для тестирования эффектов вирусных белков и хлорохина.
Рисунок 11.A-1 представляет собой схему связывания хлорохина с поверхностным гликопротеином вируса. На рисунке 11.A-2 показана область связывания вирусного поверхностного гликопротеина. В связывании участвуют 13 аминокислот. Энергия связывания хлорохина с гликопротеином Е2 вируса составляет 3 325 322 829,64 ккал/моль, что составляет около половины энергии связывания гликопротеина Е2 и порфирина. Согласно результатам рис. 4.А-2, дальнейший анализ показал, что некоторые аминокислоты (например, VAL A:952, ALA A:956, ALA B:956, ASN A:955 и др.) гликопротеина Е2 могут связываться не только с хлорохин-фосфатом, но и с порфиринами. Другими словами, хлорохин имеет одну треть шансов ингибировать вирусный гликопротеин E2 и уменьшить симптомы у пациента.
Вид связывания хлорохина и белка оболочки показан на рисунке 11.В-1. Энергия связывания хлорохина и белка оболочки 7852,58 ккал/моль, что эквивалентно лишь 4 % энергии связывания белка оболочки и порфирина. Участок связывания показан на рисунке 11.B-2. На рисунках 4.В-2 и 11.В-2 представлены некоторые аминокислоты (такие, как LEV E:28, PHE: D:20, VAL E:25) белка оболочки, которые связываются не только с хлорохин-фосфатом, но и с порфирином.
Рисунок 11.С-1 представляет собой схему связывания хлорохина с фосфопротеином нуклеокапсида. Энергия связывания хлорохина с нуклеокапсидным фосфопротеином составляет 198 815,22 ккал/моль, что эквивалентно лишь 1,4 % энергии связывания нуклеокапсидного фосфопротеина и порфирина. ALA A:50 и т. д. нуклеокапсида фосфопротеина участвуют в связывании (рисунок 12.C-2). Рисунки 4.C-2 и 11.C-2 свидетельствуют о том, что аминокислоты нуклеокапсидного фосфопротеина могут связывать порфирин, но не могут связывать хлорохин. Стыковка мембранного белка с хлорохином не произошла.
Рисунок 11. Результаты молекулярной стыковки структурных белков вируса и хлорохина (красный). A. Результаты молекулярной стыковки гликопротеина E2 и порфирина. B. Результаты молекулярной стыковки белка оболочки и порфирина. C. Результаты молекулярной стыковки нуклеокапсидного фосфопротеина и порфирина. 1. Структурные белки вируса. 2. Вид участков связывания.
Принципиальная схема связывания хлорохина с белком orf1ab показана на рисунке 12.A-1. Участок связывания белка orf1ab представлен на рисунке 12.A-2. Энергия связывания хлорохина и белка orf1ab составляет 4 584 302,64 ккал/моль, что в 8 раз больше энергии связывания между orf1ab и порфирином. Согласно результатам на рисунке 7.A-2, было показано, что некоторые аминокислоты, такие как MET 7045, PHE 7043, LYS 6836 белка orf1ab, могут быть связаны не только с фосфатом хлорохина, но и с порфирином.
Принципиальная схема связывания хлорохина с белком ORF8 показана на рисунке 12.B-1. На рисунке 12.B-2 показан участок связывания ORF8. Энергия связывания хлорохина с белком ORF8 составляет 4 707 657,39 ккал/моль, что эквивалентно лишь 37 % энергии связывания белка ORF8 с порфирином. Согласно результату, показанному на рисунке 7.B-2, аминокислоты, такие как ILE A: 74, ASP A:75, LYS A: 53 ORF8, могут связываться не только с фосфатом хлорохина, но и с порфирином.
Принципиальная схема связывания хлорохина с белком ORF7a показана на рисунке 12.C-1. На рисунке 12.C-2 представлен вид участка связывания. Энергия связывания хлорохина с белком ORF7a составляет 497 154,45 ккал/моль, что в 13 раз превышает энергию связи белка ORF7a с порфирином. Согласно результатам, показанным на рисунке 7.C-2, аминокислоты, такие как GLN A:94, ARG A:78 и LEU A:96 белка ORF7 могут связываться не только с фосфатом хлорохина, но и с порфирином.
Стыковка белков ORF3a, ORF6 и ORF10 с хлорохином не удалась.
Эти результаты показали, что хлорохин может в определенной степени ингибировать связывание E2 и ORF8 с порфирином с образованием комплекса. Кроме того, хлорохин может предотвратить атаку orf1ab, ORF3a и ORF10 на гем с образованием порфирина.
Рисунок 12. Результаты молекулярной стыковки вирусных неструктурных белков и хлорохина (красная структура). A. Результаты молекулярной стыковки белка orf1ab и хлорохина. B. Результаты молекулярной стыковки белка ORF8 и хлорохина. C. Результаты молекулярной стыковки белка ORF7a и хлорохина. 1. Неструктурные белки вируса. 2. Вид участков связывания.
3.5. Валидация эффекта фавипиравира
Рисунок 11.С-1 представляет собой схему связывания хлорохина с фосфопротеином нуклеокапсида. Энергия связывания хлорохина с нуклеокапсидным фосфопротеином составляет 198 815,22 ккал/моль, что эквивалентно лишь 1,4 % энергии связывания нуклеокапсидного фосфопротеина и порфирина. ALA A:50 и т. д. нуклеокапсида фосфопротеина участвуют в связывании (рисунок 12.C-2). Рисунки 4.C-2 и 11.C-2 свидетельствуют о том, что аминокислоты нуклеокапсидного фосфопротеина могут связывать порфирин, но не могут связывать хлорохин. Стыковка мембранного белка с хлорохином не произошла.
Таблица 1. Эффект фавипиравира
| Белок вируса | Порфирин (ккал/моль) |
Фавипиравир (ккал/моль) |
Имеет идентичные остатки |
Мишень | Отношение связывания с мишенью (фавипиравир/ порфирин) |
|---|---|---|---|---|---|
| Гликопротеин E2 | 7,530,186,265.80 | — | — | — | — |
| Белок оболочки | 219,317.76 | 597,814,480.55 | Да | Да | 2,725.79 |
| Нуклеокапсид | 15,532,506.53 | — | — | — | — |
| orf1ab | 561,571.10 | 1,052,489.88 | Да | Да | 1.87 |
| ORF8 | 12,804,859.25 | 348,589.80 | Да | — | — |
| ORF7a | 37,123.79 | 17,034,560.60 | Да | Да | 458.86 |
4. Обсуждение
4.1. Новый коронавирус произошел от древнего вируса
Для самых примитивных форм жизни, коими являются вирусы, не так-то просто увидеть их роль в связывании порфирина. Соединения порфирина широко распространены в фотосинтезирующих и нефотосинтезирующих организмах и связаны с критическими физиологическими процессами, такими как катализ, перенос кислорода и энергии. Порфирин также является древним соединением, широко распространенным на Земле. Порфирин впервые обнаружен в сырой нефти и асфальтовой породе в 1934 году. Порфирин обладает уникальными фотоэлектронными свойствами, отличной термостабильностью и имеет широкие перспективы применения в химии материалов, медицине, биохимии и аналитической химии. Его характеристики отлично подходят для применений, связанных с двухфотонным поглощением, флуоресценцией, передачей энергии и других. Перенос энергии флуоресцентного резонанса (FRET) — это безызлучательный процесс, при котором донор в возбужденном состоянии передает энергию реципиенту в основном состоянии посредством дипольного эффекта дальнего действия. FRET-характеристики порфирина могут быть основой способа выживания, на который опирался исходный вирус.
Существует множество теорий о происхождении вирусов, одна из которых называется теорией совместной эволюции, в которой вирусы могут эволюционировать из комплексов белка и нуклеиновой кислоты. Различные методы не объясняют, как вирус выжил независимо от не существовавших в начале жизни клеток, поэтому происхождение вирусов остается загадкой. В этой статье предполагается, что вирус может связываться с порфирином, что может объяснить проблему выживания оригинального вируса. Поскольку порфирин обладает характеристикой передачи энергии флуоресцентного резонанса, вирусы, которые связываются с порфиринами, могут получить энергию с помощью этого светоиндуцированного метода. Вирус, получивший энергию, может использовать ее для минимального перемещения, для выхода из состояния гибернации или перехода в него из активного состояния. Согласно результатам нашего исследования, новый коронавирус был формой жизни, зависящей от порфирина. Поэтому мы можем предположить, что новый коронавирус происходит от древнего вируса, который мог развиваться у летучих мышей на протяжении бесчисленных поколений.
4.2. Более высокая проницаемость порфиринов сквозь клеточные мембраны обуславливает большую инфекционность
Быстрая эволюция нового коронавируса также сопровождается некоторыми парадоксальными особенностями. Нынешняя теория предполагает, что новый коронавирус связывается с рецептором ACE2 человека через белок-шип. Он попадает в клетки человека по механизму фагоцитоза. Модели инфекционных заболеваний показали, что новая коронавирусная пневмония очень контагиозна. Следовательно, способность связывания белка-шипа и белка ACE2 человека должна быть большой, но в литературе имеются сообщения о том, что эта способность связывания является слабой. Что вызывает высокую инфекционность нового коронавируса? Мы считаем, что в дополнение к методу инвазии через взаимодействие шип-ACE2 вирус должен обладать оригинальным механизмом инвазии.
Медицинские работники обнаружили новый коронавирус в моче, слюне, кале и крови. Жизнеспособный вирус также может обнаруживаться в биологических жидкостях. В таких средах порфирин является доминирующим веществом. Порфириновые соединения относятся к классу азотсодержащих полимеров, и существующие исследования показали, что они обладают выраженной способностью обнаруживать клеточные мембраны и проникать сквозь них. В начале жизни молекулы вирусов с порфиринами непосредственно перемещались в исходную мембранную структуру за счет проницаемости порфирина. Это исследование показало, что гликопротеин E2 и белок оболочки нового коронавируса могут хорошо связываться с порфиринами. Поэтому коронавирус в связи с порфирином может также напрямую проникать через клеточную мембрану человека, что делает процесс инфицирования надежным. Наш валидационный анализ показал, что фавипиравир может предотвратить связывание только белка оболочки и порфирина. В то же время хлорохин может предотвращать связывание гликопротеина Е2 с порфирином лишь в определенной степени. Следовательно, инфекционность новой коронавирусной пневмонии не предотвращается этими препаратами полностью, так как связывание гликопротеина E2 и порфирина ингибируется не полностью.
4.3. Сложность индивидуального иммунитета
В некоторых теориях предполагается, что иммунный ответ возникает в организме после того, как у пациента разовьется заболевание. У некоторых пациентов после выздоровления вырабатываются иммунные антитела. Согласно нашему исследованию, гликопротеин E2, белок оболочки, нуклеокапсидный фосфопротеин, orf1ab, ORF7a и ORF8 вируса могут связываться с порфирином. Но из текущего исследования неясно, какие иммунные антитела возникали против вирусных белков.
Кроме того, некоторые пациенты могут погибнуть в результате цитокинового шторма. По сравнению с пациентами с атипичной пневмонией, анатомические характеристики умерших отличаются. Комплекс вирусных белков и порфирина может быть малорастворимым. Избыток слизи в тканях умерших пациентов был причиной избытка муцинового белка. Муцин может превратить слабо соединенные клетки в плотно соединенные и увеличить смазку между ними. Можно предположить, что действующее соединение приводит к уменьшению связи между клетками, в результате чего клетки начинают нуждаться в муцине для укрепления связи между собой в пределах тканей и для обеспечения смазывающего эффекта. Кроме того, когда пациент вступает в тяжелый инфекционный период, вирусные структурные белки, в основном, используются для сборки вирусов. Поэтому мы не можем обнаружить заметных вирусных включений в клетках тканей при аутопсии умерших пациентов.
4.4. Иммунные клетки заражаются и секретируют антитела и вирусные белки
Иммунные клетки, такие как плазматические клетки, также известны как эффекторные В-клетки. Плазматические клетки в основном наблюдаются в соединительной ткани слизистой оболочки как в пищеварительном тракте, так и в дыхательных путях. Это клетки, секретирующие антитела. Плазматические клетки выполняют функцию синтеза и хранения антител, а именно иммуноглобулинов, и участвуют в гуморальных иммунных ответах. В зависимости от источника выработки антител выделяют естественные антитела, такие как антитела анти-А и анти-В в системе групп крови ABO. По способности к участию в процессе агглютинации в ходе антигенной реакции антитела делят на полные антитела IgM и неполные антитела IgG. Обнаружение IgM и IgG в крови помогает определить, является ли организм человека инфицированным вирусом. В крови пациентов с подозрением на новую коронавирусную пневмонию содержится большое количество IgM. При лечении количество IgM у пациента снижается, а количество IgG повышается, указывая на то, что его организм вырабатывает резистентность и иммунитет. Имеются сообщения о том, что плазматические клетки также имеют рецептор ACE2, то есть для них существует путь инфекции шип-ACE2. Учитывая сообщения о том, что селезенка, костный мозг и лимфатические узлы тяжелых пациентов также сильно повреждены, мы предполагаем, что плазматические клетки также тесно связаны с инфекцией и выздоровлением пациентов с коронавирусом.
Плазматические клетки могут секретировать различные антитела, что также объясняет высвобождение вирусных белков в организме. Вирусные белки orf1ab, ORF3a и ORF10 синтезировались в клетках и атаковали гемоглобин и гем вне клеток. Вирусные белки могли покидать клетки через механизмы секреции белков. К числу секретируемых белков в основном относятся пищеварительные ферменты, антитела и некоторые гормоны. Исходя из вышеизложенной точки зрения, что инфицирование было связано с плазматическими клетками, мы полагали, что вирусные белки секретировались главным образом изнутри клетки наружу по механизму секреции антител. Один из возможных путей заключается в том, что после инфицирования плазматической клетки в ней запускаются процессы вирусной транскрипции и трансляции, а затем из клетки секретируются вирусные белки, такие как orf1ab, ORF3a и ORF10. Однако неясно, секретируются ли вирусные белки за пределы клетки путем связывания с антителами группы крови.
Мы планировали смоделировать этот механизм, но объем вычислений оказался слишком велик. После того, как мы ввели «антитела крови» в поисковую строку базы данных PDB, веб-страница показала почти 160 000 записей и почти 47 000 записей, связанных с человеком. Кроме того, моделирование молекулярной стыковки антител и белков, таких как orf1ab, представляет собой стыковку белков, процесс расчета которой является очень сложным. Поэтому мы не можем смоделировать этот механизм. Мы предлагаем другим лабораториям использовать суперкомпьютеры для моделирования этого механизма.
4.5. Вирусный белок атакует гемоглобин, высвобожденный за счет иммунного гемолиза эритроцитов
Эритроциты в основном содержат гемоглобин. Во время гемолиза гемоглобин выходит из клеток и растворяется в плазме. В этот момент способность гемоглобина переносить кислород теряется. Гемолиз происходит из-за разрыва мембран эритроцитов и растворения матрикса. Либо может происходить расширение пор мембраны эритроцита до степени, позволяющей гемоглобину покидать клетку, оставляя за собой двояковогнутую дискообразную клеточную мембрану — «гематоцит». Иммунный гемолиз — это специфический гемолиз, вызванный реакцией «антиген-антитело». Неспецифический гемолиз вызывается физическими, химическими или биологическими факторами. После гемолиза эритроцитов вирусные белки могут атаковать гемоглобин. Учитывая, что некоторые исследователи подсчитали, что люди с кровью типа O хуже заражаются COVID-19, мы предполагаем, что иммунный гемолиз может быть основным методом обеспечения атаки гемоглобина вирусными белками. Вирусные белки атакуют гемоглобин после заражения. Из-за ограниченных возможностей вычислительных инструментов мы не можем смоделировать, атакуют ли вирусные белки гемоглобин снаружи или внутри эритроцитов.
4.6. Более высокий уровень гемоглобина вызывает более высокую болезненность
Показано, что терапевтический эффект хлорохина фосфата в отношении новой коронавирусной пневмонии может быть тесно связан с аномальным метаболизмом гемоглобина у человека. Количество гемоглобина является основным биохимическим показателем крови, и его содержание различается в зависимости от пола. В норме у мужчин его уровень значимо выше, чем у женщин, что также может быть причиной того, почему мужчины заражаются новой коронавирусной пневмонией чаще, чем женщины. Кроме того, большинство пациентов с новой коронавирусной пневмонией составляют людей среднего и старшего возраста. Многие из этих пациентов имеют сопутствующие заболевания, такие как сахарный диабет. Пациенты с диабетом имеют более высокий уровень гликированного гемоглобина. Гликированный гемоглобин представляет собой дезоксигемоглобин. Гликированный гемоглобин представляет собой комбинацию гемоглобина и глюкозы крови, что является еще одной причиной высокого уровня инфицирования среди пожилых людей.
Это исследование подтвердило, что белки orf1ab, ORF3a и ORF10 могут скоординированно атаковать гем на бета-цепи гемоглобина. Атаке подвергаются как оксигенированный, так и дезоксигенированный гемоглобин. Во время атаки позиции orf1ab, ORF3 и ORF10 немного отличаются. Было показано, что, чем выше содержание гемоглобина, тем выше риск заболевания. Однако нет уверенности в том, что частота заболеваний, вызванных аномальным гемоглобином (структурным), относительно невелика. Гемоглобин пациентов и выздоравливающих должен быть объектом дальнейших исследований и лечения.
4.7. Ингибирование анаболического пути гема и развитие заболевания
В данной статье рассматривалось непосредственное вмешательство вируса в сборку гемоглобина человека. Основной причиной был слишком низкий уровень нормального гема. Гем участвует в критических биологических процессах, таких как регуляция экспрессии генов и трансляции белка. Порфирин является важным материалом для синтеза гема. Поскольку существующие данные показывают, что в организме оказывается слишком много свободного железа, это должно быть следствием того, что вирус-продуцирующая молекула конкурирует с железом за порфирин. Ингибирование анаболического пути гема и возникновение симптомов у человека.
Неясно, является ли пространственная молекулярная структура гема и порфирина у пациентов с порфирией такой же, как и у здоровых людей. При наличии аномальной структуры неясно, может ли такой порфирин связываться с вирусным белком с образованием комплекса, и может ли вирусный белок атаковать подобный гем. Эти вопросы должны быть рассмотрены в клинических и экспериментальных исследованиях.
5. Выводы
С момента возникновения эпидемии использование методов биоинформатики имеет большое научное значение для анализа ролей белков нового коронавируса (таких, как ORF8 и поверхностные гликопротеины). В этом исследовании методы прогнозирования доменов применялись для поиска консервативных доменов. Структуру белковых молекул, таких как ORF8 и поверхностных гликопротеинов, получали с помощью методов гомологического моделирования. Технология молекулярной стыковки использовалась для анализа взаимодействия связывающей части вирусных белков с гемом и порфирином. Результаты исследования показывают, что ORF8 и поверхностные гликопротеины могут объединяться с порфирином с образованием комплекса. В то же время белки orf1ab, ORF10 и ORF3a могут координированно атаковать гем, находящийся на 1-бета-цепи гемоглобина, что приводит к отщеплению железа с образованием порфирина. В результате такой атаки количество гемоглобина, который может переносить кислород и углекислый газ, уменьшается. Клетки легких испытывают чрезвычайно сильное воспаление из-за невозможности обеспечения интенсивного обмена углекислым газом и кислородом; в конечном итоге изображения ткани легких принимают вид матового стекла. Состояние пациентов с респираторными расстройствами ухудшится. Пациенты с диабетом и пожилые люди имеют более высокий уровень гликированного гемоглобина. Уровень гликированного гемоглобина снижается в результате вирусной атаки, что делает уровень глюкозы в крови пациентов нестабильным. Поскольку порфириновые комплексы вируса, продуцируемого в организме человека, ингибировали анаболический путь гема, они вызывали широкий спектр инфекций и заболеваний.
С учетом этих выводов дальнейший анализ показал, что хлорохин может предотвратить атаку orf1ab, ORF3a и ORF10 на гем с образованием порфирина и в определенной степени ингибировать связывание ORF8 и поверхностных гликопротеинов с порфиринами, эффективно облегчая симптомы респираторного дистресса. Поскольку способность хлорохина ингибировать структурные белки не слишком велика, терапевтический эффект для разных людей может быть различным. Фавипиравир может ингибировать связывание белка оболочки и белка ORF7a с порфирином, предотвращать проникновение вируса в клетки-хозяева и может связывать свободный порфирин. В связи с побочными действиями таких препаратов, как хлорохин, и возможностью аллергических реакций на них обращайтесь к квалифицированному врачу для получения подробной информации о лечении и не принимайте препарат самостоятельно.
На основании компьютерного моделирования и дискуссионного анализа этого исследования мы выдвинули предположение об основном механизме патогенности этого вируса. Вирус может сначала инфицировать клетки с рецепторами ACE2, включая иммунные клетки. Иммунные клетки производят антитела и вирусные белки. Антитела действуют на эритроциты, вызывая иммунный гемолиз. Гемоглобин высвобождается и подвергается атаке. Вирус захватывает порфирин и ингибирует метаболизм гема. Поэтому мы считаем, что поражение организма человека вирусом носит системный характер, а не ограничивается дыхательной системой.
Данная работа предназначена только для научного обсуждения, правильность выводов должна быть подтверждена другими лабораториями. Мы с нетерпением ожидаем сообщений от лабораторий, которые смогут доказать, является ли эта теория неправильной или правильной из следующих экспериментов: 1) используйте рентгеноструктурный анализ для определения структуры гемоглобина у тяжелобольных пациентов, чтобы выяснить, есть ли какие-либо отклонения; 2) в эксперименте с вирусами должны быть показаны следующие этапы: вирусные белки могут связывать порфирин; вирусные белки могут атаковать гем; вирусные белки могут атаковать гемоглобин в крови.
Заявления/h5>
Согласие этического комитета и согласие на участие
Согласие на публикацию
Доступность данных и материалов
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Финансирование
Эта работа финансировалась за счет гранта Фонда естественных наук для проекта по внедрению талантов Сычуанского университета науки и техники (номер награды 2018RCL20, грантополучатель WZL).
Вклад авторов
Финансирование получил WZL. Дизайн, анализ, написание: WZL. Курирование данных, проверка рукописи: HLL. Все авторы прочитали и согласились с опубликованной версией рукописи.
Благодарности
Об авторах
1 Факультет информатики и инженерии, Сычуаньский инженерно-технический университет, Цзыгун, 643002, Китай;
2 Факультет медико-биологических наук и технологии пищевых продуктов, Ибиньский университет, Ибинь, 644000, Китай.
Список литературы
- Дяо, K., Хань, P., Пан, T., Ли, И. и Ян, Ц. Характерные особенности визуализации HRCT в репрезентативных случаях завозной инфекции новой коронавирусной пневмонии 2019 г. (Diao, K., Han, P., Pang, T., Li, Y. & Yang, Z. HRCT Imaging Features in Representative Imported Cases of 2019 Novel Coronavirus Pneumonia). Precision Clinical Medicine (2020).
- Чан, Д. и др., Эпидемиологические и клинические характеристики новой коронавирусной инфекции на примере 13 пациентов за пределами Уханя. Китай (Chang, D. et al. Epidemiologic and clinical characteristics of novel coronavirus infections involving 13 patients outside Wuhan, China). JAMA (2020).
- Хуан С. и др. Клинические характеристики пациентов, инфицированных новым коронавирусом 2019 в Ухане, Китай (Huang, C. et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China). The Lancet (2020).
- Ли, С., Цай, Цз., Ван, С. и Ли, Ю. Возможность крупномасштабного переноса инфекции 2019 nCov от человека к человеку в первом поколении (Li, X., Zai, J., Wang, X. & Li, Y. Potential of large ‘first generation’human‐to‐human transmission of2019‐nCoV). Journal of Medical Virology (2020).
- Ван, Д. и др., Клинические характеристики 138 госпитализированных пациентов с вирусной пневмонией, вызванной новым коронавирусом 2019, Ухань, Китай (Wang, D. et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus–infected pneumonia in Wuhan, China). Jama (2020).
- Ли, Ц. и др. Ранняя динамика передачи пневмонии, вызванной новой коронавирусной инфекцией, в Ухане, Китай (Li, Q. et al. Early transmission dynamics in Wuhan, China, of novel coronavirus–infected pneumonia). New England Journal of Medicine (2020).
- Чжу, Н. и др. Новый коронавирус, выделенный у пациентов с пневмонией в Китае в 2019 г. (Zhu, N. et al. A novel coronavirus from patients with pneumonia in China, 2019). New England Journal of Medicine (2020).
- Ву, Ф. и др. Новый коронавирус, ассоциируемый с респираторными заболеваниями людей в Китае (Wu, F. et al. A novel coronavirus associated with human respiratory disease in China). Nature, 1-8 (2020).
- Лу, Х., Страттон, С. У. и Тан, И. В. Вспышка пневмонии неизвестной этиологии в Ухане, Китай: загадка и чудо (Lu, H., Stratton, C. W. & Tang, Y. W. Outbreak of Pneumonia of Unknown Etiology in Wuhan China: the Mystery and the Miracle). Journal of Medical Virology.
- Чу, Н. и др. Исследовательская группа по новому китайскому коронавирусу. Новый коронавирус, выделенный у пациентов с пневмонией в Китае (Zhu, N. et al. China Novel Coronavirus Investigating and Research Team. A novel coronavirus from patients with pneumonia in China, 2019). N Engl J Med (2020).
- Лу, Р. и др. Геномная характеристика и эпидемиология нового коронавируса 2019: заключения по поводу происхождения вируса и связывания его с рецепторами (Lu, R. et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding). The Lancet (2020).
- Ван, М. и др. Прецизионно-медицинский подход в отношении лечения коронавирусной уханьской пневмонии (Wang, M. et al. A precision medicine approach to managing Wuhan Coronavirus pneumonia). Precision Clinical Medicine (2020).
- Шехер, С. Р., Пекош, А. в сборнике «Молекулярная биология коронавируса SARS» (Schaecher, S. R. & Pekosz, A. in Molecular Biology of the SARS-Coronavirus) 153-166 (Springer, 2010).
- МакБрайд, Р. и Филдинг, Б. Ч. Роль вспомогательных белков вируса тяжелого острого респираторного синдрома (ТОРС) в патогенезе вируса (McBride, R. & Fielding, B. C. The role of severe acute respiratory syndrome (SARS)-coronavirus accessory proteins in virus pathogenesis). Viruses 4, 2902-2923 (2012).
- Ву, А. и др. Состав и дивергенция генома нового коронавируса (2019-nCoV) родом из Китая (Wu, A. et al. Genome Composition and Divergence of the Novel Coronavirus (2019-nCoV) Originating in China). Cell Host & Microbe (2020).
- Параскевис, Д. и др. Полногеномный эволюционный анализ нового коронавируса (2019-nCoV) позволяет отклонить гипотезу о его появлении в результате недавнего события рекомбинации (Paraskevis, D. et al. Full-genome evolutionary analysis of the novel corona virus (2019-nCoV) rejects the hypothesis of emergence as a result of a recent recombination event). Infection, Genetics and Evolution, 104212 (2020).
- Ли, С. и др. Регуляция отклика на стресс со стороны эндоплазматического ретикулума активностью ионных каналов, образуемых белком оболочки коронавируса, вызывающего инфекционный бронхит, модуляцией выброса вирионов, влиянием на апоптоз, репликативную способность и патогенез (Li, S. et al. Regulation of the ER Stress Response by the Ion Channel Activity of the Infectious Bronchitis Coronavirus Envelope Protein Modulates Virion Release, Apoptosis, and Pathogenesis). Frontiers in Microbiology 10, 3022 (2020).
- То, К.-К. В. и др. Постоянное выявление нового коронавируса 2019 в слюне (To, K. K.-W. et al. Consistent detection of 2019 novel coronavirus in saliva). Clinical Infectious Diseases (2020).
- Дон, Н. и др. Анализ моделей генома и белковой структуры отображает происхождение и патогенность вируса 2019-nCoV, нового коронавируса, вызвавшего вспышку пневмонии в Ухане, Китай (Dong, N. Et al Genomic and protein structure modelling analysis depicts the origin and pathogenicity of 2019-nCoV, a novel coronavirus which caused a pneumonia outbreak in Wuhan, China). F1000Research 9, 121 (2020).
- Роте, К. и др. Передача инфекции 2019-nCoV при контакте с бессимптомным носителем в Германии (Rothe, C. et al. Transmission of 2019-nCoV infection from an asymptomatic contact in Germany). New England Journal of Medicine (2020).
- Чен, Н. и др., Эпидемиологические и клинические характеристики 99 случаев пневмонии, вызванной новым коронавирусом 2019 года в Ухане, Китай: дескриптивное исследование (Chen, N. et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study). The Lancet (2020).
- Дас, Р. и Шарма, П., в сборнике «Клиническая молекулярная медицина» (Das, R. & Sharma, P. in Clinical Molecular Medicine) 327-339 (Elsevier, 2020).
- Казазян-мл., Х. Х. и Вудхэд, А. П. Синтез гемоглобина А в развивающемся плоде (Kazazian Jr, H. H. & Woodhead, A. P. Hemoglobin A synthesis in the developing fetus). New England Journal of Medicine 289, 58-62 (1973).
- Лю, Дж. и др. Общие и отличающиеся аспекты патологии и патогенеза новых патогенных для человека коронавирусных инфекций SARS‐CoV, MERS‐CoV и 2019‐nCoV. (Liu, J. et al. Overlapping and discrete aspects of the pathology and pathogenesis of the emerging human pathogenic coronaviruses SARS‐CoV, MERS‐CoV, and 2019‐nCoV). Journal of Medical Virology (2020).
- Ван, М. и др. Ремдесивир и хлорохин эффективно ингибируют недавно появившийся коронавирус (2019-n-CoV) in vitro (Wang, M. et al. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro). Cell Research, 1-3 (2020).
- Бернардо-Сейсдедос, Г., Джил, Д., Блуэн, Ж-М., Ришар, Э. и Милле, О. Заболевания белкового гомеостаза (Bernardo-Seisdedos, G., Gil, D., Blouin, J.-M., Richard, E. & Millet, O. in Protein Homeostasis Diseases) 389-413 (Elsevier, 2020).
- Ламеда, И. Л. и Кох, Т. Р в сборнике «Заболевания печени» (Lameda, I. L. P. & Koch, T. R. in Liver Diseases) 107-116 (Springer, 2020).
- Бэйли, Т. Л., Джонсон, Дж., Грант, К. Э., Нобл, У. С. Программа МЕМЕ SUITE (Bailey, T. L., Johnson, J., Grant, C. E. & Noble, W. S. The MEME suite). Nucleic acids research 43, W39-W49 (2015).
- Бэйли, Т. Л. и др. ПРОГРАММА МЕМЕ SUITE: средство поиска известных мотивов и открытия новых (Bailey, T. L. et al. MEME SUITE: tools for motif discovery and searching). Nucleic acids research 37, W202-W208 (2009).
- Бэйли, Т. Л., Уильямс, Н., Мислех, С. и Ли, У. У. МЕМЕ: обнаружение и анализ мотивов с последовательностях ДНК и белков (Bailey, T. L., Williams, N., Misleh, C. & Li, W. W. MEME: discovering and analyzing DNA and protein sequence motifs). Nucleic acids research 34, W369-W373 (2006).
- Шведе, Т., Копп, Й., Гуэ, Н. и Петиш, М. SWISS-MODEL: автоматизированный сервер гомологического моделирования белков (Schwede, T., Kopp, J., Guex, N. & Peitsch, M. C. SWISS-MODEL: an automated protein homology-modeling server). Nucleic acids research 31, 3381-3385 (2003).
- Бьязини, М. и др. SWISS-MODEL: моделирование третичной и четвертичной структуры белков и использованием эволюционной информации (Biasini, M. et al. SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information). Nucleic acids research 42, W252-W258 (2014).
- Studio, D. Discovery Studio. Accelrys [2.1] (2008).
Препараты для лечения анемии (при снижении уровня гемоглобина)
{{/if}} {{each list}} ${this} {{if isGorzdrav}}Удалить
{{/if}} {{/each}} {{/if}}На сайте «Горздрав» вы можете заказать препараты для поднятия гемоглобина в крови у взрослых по доступной цене. На сайте предусмотрена возможность сравнения стоимости во всех пунктах сети для удобного подбора подходящего варианта. Представленные лекарства при низком гемоглобине прошли обязательную сертификацию и сопровождаются подробной инструкцией по применению. Чтобы приобрести средства с повышенным содержанием железа в составе, достаточно заказать товар на сайте. Оплата и выдача покупки проводятся в кассе аптеки.
Заказать препараты для повышения гемоглобина
В сеть аптек «Горздрав» входит более 800 отделений в Москве и Санкт-Петербурге и в их прилегающих областях. При заказе онлайн вы можете посмотреть расположение всех аптек и подобрать удобную аптеку для получения товара. В каталоге представлены только препараты для повышения гемоглобина, прошедшие сертификацию, которые можно посмотреть на странице интересующего лекарства. Благодаря продуманной сортировке по действующему веществу на сайте можно ознакомиться с аналогами препаратов.
Применение препаратов при низком гемоглобине
Гемоглобин обеспечивает транспортировку кислорода ко всем внутренним органам. При его недостатке снижается трудоспособность человека, ухудшается общее состояние, появляется слабость, головокружения, развивается анемия. Это заболевание наблюдается у 10-30% взрослого населения. Причинами снижения гемоглобина и появления анемии могут быть:
- Несбалансированное питание;
- Хронические кровопотери;
- Язвы желудка и кишечника;
- Глистные инвазии;
- Грыжи.
На организм также влияют курение, алкоголь, стресс, переутомления. При низком уровне гемоглобина назначается специальная диета и курс препаратов железа. Лекарства принимают не менее 1,5-2 месяцев, а после нормализации гемоглобина продолжают прием в половинной дозе еще 4-6 недель[1].
Формы выпуска
Лекарства для повышения уровня гемоглобина в крови выпускаются в различных формах, что не влияет на их действие:
- Таблетки,
- Капсулы,
- Сиропы,
- Растворы,
- Ампулы.
Для кого
Препараты железа при низком гемоглобине могут назначаться как взрослым, так и детям:
- У женщин анемия наблюдается чаще, чем у мужчин, поэтому женщинам следует регулярно сдавать анализ крови и, при необходимости, принимать железосодержащие препараты.
- Для беременных разработаны сбалансированные лекарственные комплексы для поднятия уровня гемоглобина.
- Анемия может появиться и у детей. Для нормализации уровня железа в крови прописываются специальные детские лекарства.
Противопоказания
Средства для поднятия уровня гемоглобина обладают определенными противопоказаниями. Перед покупкой обязательно ознакомьтесь с инструкцией. Особое внимание стоит уделить людям с:
- Индивидуальной непереносимостью компонентов;
- Нарушениями механизма усваивания железа;
- Анемией не железодефицитного характера.
ПЕРЕД ПРИМЕНЕНИЕМ ПРЕПАРАТОВ НЕОБХОДИМО ОЗНАКОМИТЬСЯ С ИНСТРУКЦИЕЙ ПО ПРИМЕНЕНИЮ ИЛИ ПРОКОНСУЛЬТИРОВАТЬСЯ СО СПЕЦИАЛИСТОМ.
Список литературы:
- [1] Руководство по первичной медико-санитарной помощи. А. А. Баранов, Денисов И.Н., Чучалин А.Г. ГЭОТАР-Медиа, 2006. — 1584 с.
Узнаем как ой должен быть гемоглобин у мужчин? Норма гемоглобина у мужчин по возрасту
В расшифровке общего анализа крови есть показатель, отражающий уровень гемоглобина, обозначается Hb. Что это означает? Гемоглобин играет весьма значительную роль в работе организма. При любых отклонениях от нормы нужно предпринимать меры, ведь от его уровня зависит жизнь человека. В данной статье разберем подробнее, какой должен быть гемоглобин у мужчин. Содержание этого «кислородного белка» у сильной половины отличается от показателей у женщин.
Какую роль в организме играет гемоглобин
Гемоглобин находится в красных кровеносных тельцах. По сути, это железосодержащий белок, который имеет способность связываться с кислородом, что позволяет доносить во все ткани организма О2 и выводить СО2. Процесс этот является самым главным в организме. Гемоглобин, снабжая головной мозг и все органы кислородом, обеспечивает нормальную жизнедеятельность в целом.
Основные функции гемоглобина крови:
- Транспортная. Снабжение тканей и органов кислородом и выведение из организма человека углекислого газа.
- Поддержка оптимального уровня Ph в организме. Железо, входящее в состав гемоглобина, нейтрализует кислоты.
- Взаимосвязь количества эритроцитов и гемоглобина. Они должны быть в равном соотношении. Насыщенность цвета крови напрямую зависит от содержания железа. Когда кровь жидкой консистенции, алая — это говорит о пониженном гемоглобине и наоборот, если цвет темный, кровь густая — это свидетельство повышенного уровня Hb.
Для оценки уровня гемоглобина существуют определенные нормы. Какой должен быть гемоглобин у мужчин? В среднем 132-170 г/л. У женщин данный показатель ниже — в норме от 112 до 157 г/л.
Гемоглобин в крови у мужчин
Почему у мужчин уровень гемоглобина на порядок выше, чем у женщин? Ответ прост. Наличие тестостерона в мужском организме требует дополнительного кислорода, именно он подпитывает все клетки при дополнительных нагрузках, которые часто возникают у сильной половины в процессе жизнедеятельности. Обычно мужчины переносят более тяжелые физические нагрузки, так установлено природой, поэтому и потребность в кислороде у них выше. Однако не только от пола зависит уровень такого жизненно важного компонента крови как гемоглобин. Норма у мужчин по возрасту (таблица приводит эти показатели) тоже разная.
Возраст, лет | Норма гемоглобина г/л |
10–13 | 120–150 |
13–16 | 120–160 |
16–18 | 118–154 |
18–45 | 135–175 |
45–66 | 132–173 |
>66 | 127–175 |
Узнать уровень гемоглобина можно только путем лабораторных исследований, сдав общий анализ крови. Отмирание старых клеток и появление новых — непрерывный процесс. Так, за один час в крови человека отмирает и вновь рождается миллиард клеток. Существует прямая связь между количеством эритроцитов и уровнем гемоглобина крови. В случае, если по каким-то причинам эритроцитов вырабатывается больше, происходит увеличение гемоглобина, и наоборот. Влияет на это множество факторов, в том числе и образ жизни человека, неправильное питание.
Задумываетесь над тем, какой должен быть гемоглобин у мужчин зрелого возраста? В норме он не должен быть меньше 130 г/л, но и не превышать 170 г/л. С годами этот показатель начинает меняться в меньшую сторону. С угасанием уровня тестостерона уменьшается и количество гемоглобина. Если показатели, не соответствуют приведенным в таблице по возрасту на +-5 единиц, необходимо проконсультироваться у доктора о причинах и начать лечение.
Низкий гемоглобин
Мы разобрались, какой должен быть гемоглобин у мужчин в норме. Если цифры ниже приведенных, почему так бывает? Снизиться гемоглобин в мужском организме может по ряду причин, вот основные:
- Недостаток железа в организме. Это может произойти при несбалансированном питании, исключении из рациона мясных блюд, их недостатке, переходе на вегетарианство
- Избыток или недостаток витаминов. И здесь первопричина — неправильное питание, бедное витаминами, полезными веществами и микроэлементами. Стоит отметить, что передозировка витамина С может привести к тому, что организм не будет усваивать витамин В12. Это также негативно сказывается на организме в целом.
- Хронические формы воспалений кишечника. При нарушении структуры тканей органов ЖКТ нарушается усвоение железа.
- Различные инфекции, при которых происходит распад клеток крови.
- Большая кровопотеря при операциях, травмах.
- Вредные привычки -курение, алкоголизм.
Определить наличие анемии можно по следующим симптомам: бледная тусклая кожа, повышенная утомляемость, головокружение, учащенный пульс, выпадение волос.
Как повысить гемоглобин крови
Вы сдали анализ крови на гемоглобин. Норма у мужчин по возрасту (таблица поможет вам в этом) определена. Если показатель ниже нормы, доктор назначит лечение, чтобы повысить уровень Hb. Лечение анемии проходит не только посредством приема лекарственных препаратов железа, грамотный диетолог должен составить вам правильный рацион питания. Из продуктов мы получаем железо двух видов: гемовое (животного происхождения), оно усваивается в организме на 30% и негемовое (растительного), усвоение идет всего лишь на 7%. Что посоветует диетолог включить в свое ежедневное меню для получения организмом железа? Чернослив, курагу, зерновые мюсли, свежевыжатые соки, свежую зелень, гречку, овсянку, «Бородинский» хлеб и, конечно же, красное мясо и печень.
Однако получение чистого железа не будет залогом лечения анемии у мужчин, проходит оно длительно. Необходимо употреблять витамины, которые будут способствовать усвоению железа. Важна для этого группа витаминов С, В (в особенности В12). Каждый прием пищи нужно завершать долькой яблока или граната. Исключить из рациона продукты, которые задерживают всасывание железа — это шоколад, кофе, кисломолочные напитки, твердый сыр, сгущенка, сдобный хлеб.
Чем опасен повышенный гемоглобин
С низким уровнем Hb мы разобрались, а о чем говорит повышенный гемоглобин у мужчин? Причинами отклонения от нормы в большую сторону могут служить следующие факторы:
- Тяжелые физические нагрузки или интенсивные тренировки.
- Опухоль.
- Курение.
- Сбои в сердечно-сосудистой системе.
- Непроходимость кишечника.
- Сахарный диабет.
Симптомы повышенного гемоглобина: общая вялость, ухудшение зрение, бледность кожи, отсутствие аппетита, воспаления мочеполовой системы. Впоследствии повышенный уровень гемоглобина может привести к изменению давления в высокую сторону, к проблемам с сердечно-сосудистой системой, образованию тромбов.
Зная, какой нормальный уровень гемоглобина у мужчин, и при этом обнаружив у себя завышенные цифры, не стоит сразу паниковать. В некоторых случаях это считается нормой.
- У альпинистов гемоглобин всегда превышает норму в 170 г/л. Связано это непосредственно с их профессиональной деятельностью.
- У жителей горных районов гемоглобин всегда повышен, так как у них идет постоянное перенасыщение крови кислородом.
- Во время интенсивных занятий спортом необходимо большее количество О2 мышцам, поэтому повышается уровень гемоглобина крови.
Концентрации гемоглобина у коренных гренландцев ниже, чем у датчан: оценка гемоглобина у 234 гренландцев и у 2804 датчан
Задача: Сравнить концентрации гемоглобина у гренландцев и датчан.
Методы: Гемоглобин измерялся в ходе обследования населения в 1993–1994 годах, в котором участвовали 234 коренных гренландских индейца (115 мужчин) в возрасте 19–82 года.и в округе Копенгаген в 1983–1984 годах, где проживало 2804 кавказских датчанина (1444 мужчины) в возрасте 30–60 лет. Гренландские участники были жителями столицы Нуук (n = 70), города Илулиссат (n = 74) и четырех поселений в районе Уумманнак (n = 90). Достоверность различий оценивали по t-критерию Стьюдента и xi2-критерию. Корреляции оценивали по коэффициенту корреляции Спирмена (rs).
Полученные результаты: Гренландцы: уровни гемоглобина не коррелировали с возрастом или потреблением традиционных продуктов и не различались существенно в трех жилых районах.Средний гемоглобин был выше у мужчин, 146 +/- 9,6 (SD) г / л, чем у женщин, 132 +/- 9,6 г / л (p <0,0001). Средний гемоглобин у богатых железом мужчин с сывороточным ферритином> 32 мкг / л (n = 104) составлял 146 +/- 9,3 г / л, а у женщин с высоким содержанием железа (n = 68) 133 +/- 10,4 г / л ( р <0,0001). 5-й процентиль гемоглобина у мужчин с высоким содержанием железа составлял 133 г / л (8,3 ммоль / л), а у женщин - 118 г / л (7,3 ммоль / л). Распространенность железодефицитной анемии (т.е. ферритин <13 мкг / л и Hb <5-й перцентиль для мужчин и женщин с высоким содержанием железа) составляла 0% у мужчин, 2.78% у женщин младше 50 лет и 0% у женщин старше 50 лет. Датчане: средний гемоглобин у мужчин составлял 154 +/- 10,0 г / л, а у женщин 138 +/- 10,4 г / л (p <0,0001). Гемоглобин у мужчин с высоким содержанием железа (n = 1379) (т.е. ферритин в сыворотке> 32 мкг / л) составлял 154 +/- 10,7 г / л, а у женщин с высоким содержанием железа (n = 1003) 140 +/- 9,6 г / л. (р <0,0001). Средний гемоглобин был ниже в пременопаузе, чем у женщин в постменопаузе (p <0,0001). 5-й процентиль гемоглобина у мужчин с высоким содержанием железа составил 137 г / л (8,5 ммоль / л), а у женщин - 124 г / л (7.7 ммоль / л). Распространенность железодефицитной анемии (т.е. ферритин <13 мкг / л и Hb <5-й процентиль для мужчин и женщин с высоким содержанием железа) составляла 0% у мужчин, 1,92% у женщин <или = 50 лет и 0% у женщин> 50. лет возраста.
Заключение: Концентрации гемоглобина у гренландцев были значительно ниже, чем у датчан, как у мужчин (p <0,0001), так и у женщин (p <0.0001). Дельта (средний гемоглобин) у мужчин составляла 8,0 г / л (0,5 ммоль / л), а у женщин - 6,2 г / л (0,4 ммоль / л). Вариации уровня гемоглобина могут быть связаны с генетическими различиями.
Продолжительность ночного сна и риск возникновения анемии у населения Китая: проспективное когортное исследование
Coresh, J., Astor, B. & Sarnak, M. J. Доказательства повышенного риска сердечно-сосудистых заболеваний у пациентов с хронической болезнью почек. Текущее мнение в нефрологии и гипертонии 13 , 73–81 (2004).
Артикул PubMed Google ученый
Каллтон Б.Ф. и др. . Влияние анемии на госпитализацию и смертность пожилых людей. Кровь 107 , 3841–3846, https://doi.org/10.1182/blood-2005-10-4308 (2006).
CAS Статья PubMed Google ученый
Донг, X. и др. . Популяционное исследование гемоглобина, расы и смертности у пожилых людей. Журналы по геронтологии . Серия A, Биологические и медицинские науки 63 , 873–878 (2008).
Артикул Google ученый
Пеннинкс, Б. У., Пахор, М., Вудман, Р. С. и Гуральник, Дж. М. Анемия в пожилом возрасте связана с повышенной смертностью и госпитализацией. Журналы по геронтологии . Серия A, Биологические и медицинские науки 61 , 474–479 (2006).
Артикул Google ученый
Закай Н.А. и др. . Проспективное исследование статуса анемии, концентрации гемоглобина и смертности в когорте пожилых людей: Cardiovascular Health Study. Архив внутренней медицины 165 , 2214–2220, https://doi.org/10.1001/archinte.165.19.2214 (2005).
ADS Статья PubMed Google ученый
Ананд И. и др. . Анемия и ее связь с клиническим исходом при сердечной недостаточности. Тираж 110 , 149–154, https://doi.org/10.1161/01.CIR.0000134279.79571.73 (2004).
Артикул PubMed Google ученый
Ли, У. К. и др. . Анемия: значительный риск сердечно-сосудистой смертности после инфаркта миокарда с подъемом сегмента ST, осложненного сопутствующими заболеваниями артериальной гипертензии и заболевания почек. PloS one 12 , e0180165, https://doi.org/10.1371/journal.pone.0180165 (2017).
Артикул PubMed PubMed Central Google ученый
Спенс Р. К. Экономическое бремя анемии при сердечной недостаточности. Клиники сердечной недостаточности 6 , 373–383, https://doi.org/10.1016/j.hfc.2010.02.003 (2010).
Артикул PubMed Google ученый
Танимура, М. и др. . Влияние анемии на сердечно-сосудистую гемодинамику, терапевтическую стратегию и клинические результаты у пациентов с сердечной недостаточностью и гемодинамическими застойными явлениями. Журнал циркуляции: официальный журнал Японского общества циркуляции 81 , 1670–1677, https://doi.org/10.1253/circj.CJ-17-0171 (2017).
Артикул Google ученый
Штауфер М. Э. и Фан Т. Распространенность анемии при хронической болезни почек в США. PloS one 9 , e84943, https://doi.org/10.1371/journal.pone.0084943 (2014).
ADS Статья PubMed PubMed Central Google ученый
Portoles, J. et al. . Развитие анемии связано с плохим прогнозом при хронической болезни почек 3 стадии NKF / KDOQI. BMC нефрология 14 , 2, https://doi.org/10.1186/1471-2369-14-2 (2013).
CAS Статья PubMed PubMed Central Google ученый
Гаучи Р., Хантер М., Брюс Д. Г., Дэвис В. А. и Дэвис Т. М. Э. Анемия, осложняющая диабет 2 типа: распространенность, факторы риска и прогноз. Журнал диабета и его осложнений 31 , 1169–1174, https://doi.org/10.1016/j.jdiacomp.2017.04.002 (2017).
Артикул PubMed Google ученый
Gu, L., Lou, Q., Wu, H., Ouyang, X. & Bian, R. Отсутствие связи между анемией и прогрессированием почечной недостаточности у китайских пациентов с диабетом 2 типа. Журнал исследований диабета 7 , 42–47, https://doi.org/10.1111/jdi.12368 (2016).
Артикул PubMed Google ученый
Пейрано, П. Д. и др. . Нарушения сна и железодефицитная анемия в младенчестве. Лекарство от сна 11 , 637–642, https://doi.org/10.1016/j.sleep.2010.03.014 (2010).
Артикул PubMed PubMed Central Google ученый
Пейрано, П., Алгарин, К., Гарридо, М., Алгарин, Д. и Лозофф, Б. Железодефицитная анемия связана с измененными характеристиками спинных веретен во сне NREM в младенчестве. Нейрохимические исследования 32 , 1665–1672, https://doi.org/10.1007/s11064-007-9396-8 (2007).
CAS Статья PubMed PubMed Central Google ученый
Ферри, Дж. Э. и др. . Проспективное исследование изменения продолжительности сна: ассоциации со смертностью в когорте Уайтхолла II. Сон 30 , 1659–1666 (2007).
Артикул PubMed PubMed Central Google ученый
Gangwisch, J. E. et al. . Короткая продолжительность сна как фактор риска гипертонии: анализ первого Национального исследования здоровья и питания. Гипертония 47 , 833–839, https://doi.org/10.1161/01.HYP.0000217362.34748.e0 (2006).
CAS Статья PubMed Google ученый
Икехара, С. и др. . Связь продолжительности сна со смертностью от сердечно-сосудистых заболеваний и других причин для японских мужчин и женщин: исследование JACC. Сон 32 , 295–301 (2009).
Артикул PubMed PubMed Central Google ученый
Пател С. Р. и др. . Проспективное исследование продолжительности сна и риска смерти у женщин. Сон 27 , 440–444 (2004).
Артикул PubMed Google ученый
Song, Q. et al . Длительная продолжительность сна и риск ишемического инсульта и геморрагического инсульта: проспективное исследование Кайлуан. Научные отчеты 6 , 33664, https://doi.org/10.1038/srep33664 (2016).
ADS CAS Статья PubMed PubMed Central Google ученый
Сун, К., Лю, X., Чжоу, В., Ван, X. и Ву, С. Изменения продолжительности сна и риска метаболического синдрома: проспективное исследование Кайлуан. Научные отчеты 6 , 36861, https://doi.org/10.1038/srep36861 (2016).
ADS CAS Статья PubMed PubMed Central Google ученый
Ван, X., Лю, X., Сун, Q. & Wu, S. Продолжительность сна и риск инфаркта миокарда и смерти от всех причин в китайской популяции: исследование Кайлуань. Лекарство от сна 19 , 13–16, https://doi.org/10.1016/j.sleep.2015.09.027 (2016).
Артикул PubMed Google ученый
Lipsic, E. et al. . Уровни гемоглобина и 30-дневная смертность у пациентов после инфаркта миокарда. Международный кардиологический журнал 100 , 289–292, https://doi.org/10.1016/j.ijcard.2004.10.043 (2005).
Артикул PubMed Google ученый
Sabatine, M. S. et al. . Связь уровня гемоглобина с клиническими исходами при острых коронарных синдромах. Тираж 111 , 2042–2049, https://doi.org/10.1161/01.CIR.0000162477.70955.5F (2005).
CAS Статья PubMed Google ученый
Джаковска М., Кумари М. и Степто А. Сон и биомаркеры в Английском продольном исследовании старения: ассоциации с С-реактивным белком, фибриногеном, дегидроэпиандростерона сульфатом и гемоглобином. Психонейроэндокринология 38 , 1484–1493, https://doi.org/10.1016/j.psyneuen.2012.12.015 (2013).
CAS Статья PubMed Google ученый
Капуччио, Ф. П. и др. . Гендерные ассоциации короткой продолжительности сна с распространенной и эпизодической артериальной гипертензией: исследование Whitehall II. Гипертония 50 , 693–700, https://doi.org/10.1161/HYPERTENSIONAHA.107.095471 (2007).
CAS Статья PubMed PubMed Central Google ученый
Kim, S. J. et al. . Генетическая связь короткой продолжительности сна с заболеваемостью гипертонией — 6-летнее наблюдение в корейском геномном и эпидемиологическом исследовании. Журнал циркуляции: официальный журнал Японского общества циркуляции 76 , 907–913, JST.JSTAGE / circj / CJ-11-0713 (2012).
Ву, С. и др. . Распространенность идеального здоровья сердечно-сосудистой системы и ее связь с четырехлетними сердечно-сосудистыми событиями в промышленном городе на севере Китая. Тираж. Качество сердечно-сосудистой системы и исходы 5 , 487–493, https://doi.org/10.1161/CIRCOUTCOMES.111.963694 (2012).
Артикул PubMed Google ученый
Song, Q. et al . Продолжительность сна является независимым фактором риска развития фибрилляции предсердий у населения Китая: проспективное когортное исследование. Научные отчеты 7 , 3679, https://doi.org/10.1038/s41598-017-04034-8 (2017).
ADS Статья PubMed PubMed Central Google ученый
Фоли, Д. Дж. Эпидемиологический взгляд на один рассказ о двусторонней гипотезе. Обзоры медицины сна 8 , 155–157; обсуждение 175–156, https://doi.org/10.1016/j.smrv.2004.02.002 (2004).
Розен, К.Л. и др. . Обструктивное апноэ во сне и серповидноклеточная анемия. Педиатрия 134 , 273–281, https://doi.org/10.1542/peds.2013-4223 (2014).
Артикул PubMed PubMed Central Google ученый
Хан, А. М., Ашизава, С., Хлебович, В. и Аппель, Д. В. Анемия старения и синдром обструктивного апноэ во сне. Сон и дыхание = Schlaf & Atmung 15 , 29–34, https: // doi.org / 10.1007 / s11325-010-0326-7 (2011).
Артикул Google ученый
Grandner, M. A. et al. . Чрезмерная продолжительность сна и повышенный уровень С-реактивного белка: влияние пола и этнорасовой группы. Sleep 36 , 769–779E, https://doi.org/10.5665/sleep.2646 (2013).
Артикул PubMed PubMed Central Google ученый
Патель, С. Р. и др. . Продолжительность сна и биомаркеры воспаления. Сон 32 , 200–204 (2009).
Артикул PubMed PubMed Central Google ученый
Мейер-Эверт, Х. К. и др. . Влияние потери сна на С-реактивный белок, воспалительный маркер сердечно-сосудистого риска. Журнал Американского кардиологического колледжа 43 , 678–683, https: // doi.org / 10.1016 / j.jacc.2003.07.050 (2004).
CAS Статья PubMed Google ученый
Пенджаби, Н. М. Эпидемиология обструктивного апноэ сна у взрослых. Труды Американского торакального общества 5 , 136–143, https://doi.org/10.1513/pats.200709-155MG (2008).
Артикул PubMed PubMed Central Google ученый
Сонг, К., Лю, X., Ван, X. и Ву, С. Возрастные и гендерные связи между продолжительностью сна и эпизодической гипертонией в китайском населении: исследование Кайлуань. Журнал гипертонии человека 30 , 503–507, https://doi.org/10.1038/jhh.2015.118 (2016).
CAS Статья PubMed Google ученый
Zhang, Q. et al. . Идеальные показатели здоровья сердечно-сосудистой системы и риски ишемического и внутримозгового геморрагического инсульта. Инсульт 44 , 2451–2456, https://doi.org/10.1161/STROKEAHA.113.678839 (2013).
Артикул PubMed Google ученый
Zhu, J. et al. . Обновление 2016 г. Китайского руководства по профилактике и лечению дислипидемии у взрослых. Китайский журнал обращения 31 , 937–953 (2016).
Google ученый
Cai, J. et al. . Оценка эффективности содержания гемоглобина ретикулоцитов в диагностике железодефицитной анемии у взрослых китайцев. Питательные вещества 9 , https://doi.org/10.3390/nu9050450 (2017).
Распространенность и факторы риска тяжести и типа анемии у малавийских мужчин и женщин: городские и сельские различия | Показатели здоровья населения
Анемия имеет умеренное значение для общественного здравоохранения среди женщин и умеренное значение для здоровья мужчин в этих малавийских сельских и городских популяциях, в соответствии с классификацией ВОЗ, основанной на распространенности [28].Многопараметрический анализ показывает, что для мужчин увеличение возраста и ВИЧ-инфекция были независимыми факторами риска как легкой, так и средней или тяжелой анемии. Недостаточный вес также был связан с легкой анемией. Среди женщин в возрасте от 35 до 54 лет беременность, первородство и ВИЧ-инфекция были факторами риска анемии любой степени тяжести. Кроме того, у женщин со средним или высшим образованием, а также у женщин с шестью и более беременностями выше шансы на легкую анемию.
Распространенность анемии среди небеременных и беременных женщин, о которых здесь сообщается, ниже, чем по данным DHS Малави.DHS может исключать относительно более здоровых участников, которых нет дома, или состав населения может отличаться [24]. Различия в распространенности анемии между мужчинами и женщинами были минимальными после 55 лет, а после 65 лет распространенность была выше у мужчин. Это, возможно, указывает на большую биологическую и социальную уязвимость в репродуктивном возрасте у женщин и пожилых мужчин [31–33]. Также вероятно, что вмешательства, нацеленные на женщин репродуктивного возраста, могут защитить их от анемии в более позднем возрасте.Это также подтверждается более высоким риском анемии среди никогда не беременных женщин, хотя это может быть связано с их пригодностью для воспроизводства.
Более высокая распространенность анемии в сельских регионах объясняется неравенством в предоставлении медицинских услуг и доступе к ним, риском заболеваний, предпочтениях в отношении фертильности и генетических состояниях, таких как серповидно-клеточная анемия [16, 34–36]. Хотя городское / сельское проживание не было признано независимым фактором риска анемии любой степени тяжести у мужчин или женщин, общие показатели анемии были выше у городских женщин, чем у сельских женщин, но выше у сельских, чем у городских мужчин.Существовали также различия между городскими и сельскими районами по типам анемии. Микроцитарная анемия была более распространена среди городских жителей. Дефицит железа может быть менее распространен в сельской местности, поскольку сельские жители имеют лучший доступ к разнообразным и более питательным источникам пищи; продукты, богатые железом, такие как яйца, зеленые листовые овощи и рыба, обитающая на берегу озера; и фрукты, содержащие витамин С, улучшающие усвоение железа. Сельские женщины могут иметь больше шансов получить доступ к богатым минералами глиняным гранулам, традиционно потребляемым во время беременности [37].
Анемия обратно пропорциональна экономическому развитию на уровне страны / региона, благосостоянию на уровне домохозяйства и доходу и образованию на индивидуальном уровне [16, 24, 38]. Хотя исследования показали связь между низким социально-экономическим статусом и всеми уровнями анемии, наши результаты показывают, что у женщин с более высоким уровнем образования вероятность развития легкой анемии выше [32, 34, 39]. Мы наблюдали гендерные различия в распространенности анемии среди различных профессиональных групп. Среди мужчин наиболее распространены фермеры / рыбаки, а среди женщин — наемные рабочие.Это указывает на то, что различия в драйверах анемии у мужчин связаны с факторами, которые влияют на выработку, а также на распад или потерю эритроцитов. Распространенность микроцитарной анемии увеличивалась с повышением уровня образования и была самой низкой среди фермеров. Вполне вероятно, что в рационе более образованных женщин из более обеспеченных семей, как и у городских жителей, возможно, отошли от традиционных диет и добавок, что повысило их риск развития анемии, связанной с питанием. И наоборот, сельское хозяйство / рыболовство могут увеличить риск других инфекционных заболеваний, вызывающих анемию, таких как шистосомоз и анкилостомоз, и такие контакты могут быть более распространены среди мужчин.
Повторные беременности и короткие интервалы между родами истощают запасы железа, что приводит к анемии, что объясняет связь более высокого паритета с легкой анемией [7, 40, 41]. Напротив, более высокий паритет не был связан с анемией средней или тяжелой степени. Вмешательства по борьбе с анемией обычно нацелены на беременных женщин, поэтому повторнородящие женщины постоянно обращаются к вмешательствам по борьбе с анемией, таким как добавка железа / фолиевой кислоты, обработанные инсектицидами сетки (ITN), химиопрофилактика малярии, дегельминтизация, тестирование на ВИЧ и последующее лечение.Это может смягчить эффект паритета на анемию и поддерживать уровень гемоглобина на легкой стадии анемии. Высокий риск легкой и средней или тяжелой анемии у первородящих женщин может быть связан с их физической подготовкой к воспроизводству и сопутствующими сопутствующими заболеваниями. Хотя ВИЧ-инфекция и статус лечения были скорректированы, остаточное искажение все еще возможно, исходя из степени тяжести / стадии ВИЧ, которые не учитываются в этом анализе.
Низкий ИМТ (<18,5 кг / м 2 ) был связан с легкой анемией у мужчин.Многие исследования показали обратную связь между ИМТ и анемией [31, 32, 34, 42]. Многострановой многоуровневый анализ уровней гемоглобина у африканских женщин показал более сильную связь анемии с социально-экономическими и контекстными факторами, чем с ИМТ [11]. Было показано, что дефицит питательных микроэлементов играет разную роль в этиологии анемии различной степени тяжести [17]. Дефицит железа и фолиевой кислоты был связан с легкой анемией, в то время как дефицит витаминов A и B12, как было показано, играет важную роль в этиологии тяжелой анемии [6, 17].Хотя высокий ИМТ не обязательно означает лучшее потребление питательных микроэлементов, люди с недостаточным весом более склонны к другим сопутствующим сопутствующим заболеваниям и, следовательно, к худшему питанию.
Наши результаты показывают, что зарегистрированное употребление алкоголя значительно снижает вероятность умеренной и тяжелой анемии среди мужчин, даже после поправки на возраст, образование, место жительства, род занятий, ИМТ, курение и ВИЧ-статус. Есть некоторые свидетельства того, что традиционно сваренное пиво в SSA имеет достаточно высокое содержание железа, чтобы защищать его от дефицита железа [43, 44].Однако из-за социальной природы употребления алкоголя нельзя исключать роль обратной причинно-следственной связи.
Наблюдалась сильная связь со всеми степенями тяжести анемии и ВИЧ. Анемия — частое осложнение ВИЧ-инфекции, и было показано, что она усиливается по мере прогрессирования заболевания [45, 46]. Было показано, что использование АРТ снижает распространенность анемии среди ВИЧ-инфицированных [47, 48], но наши результаты показывают, что в этих условиях, когда анемия не может активно лечиться у посетителей клиник АРТ, это преимущество может быть замечено только через несколько лет лечение.И наоборот, использование некоторых АРТ (зидовудина) может быть связано с повышенным риском анемии из-за миелосупрессии [47, 49]. Несмотря на умеренную распространенность ВИЧ в этом исследовании, почти 20% тяжелой анемии среди населения были связаны с ВИЧ.
Бремя и распространение анемии в этой группе населения имеет несколько последствий. Поскольку было показано, что анемия отрицательно влияет на физическую работоспособность и снижает индивидуальную продуктивность, вероятным последствием этого является снижение доходов домохозяйств и продовольственная безопасность, особенно для таких групп населения, как наше, которые в основном полагаются на натуральное сельское хозяйство / рыболовство и другие часто сезонные источники доход [16, 50].Анемия может способствовать плохим исходам беременности и родов у женщин репродуктивного возраста, что может ухудшить и без того плохие материнские и перинатальные индексы [16] и может противодействовать улучшенным исходам, которые обычно наблюдаются у образованных городских женщин. Анемия среди ВИЧ-инфицированных связана с более низким качеством жизни, прогрессированием заболевания и повышенной смертностью; и, следовательно, может ухудшить прогноз [45, 51], но не может регулярно проходить скрининг во всех службах ВИЧ.
Результаты нашего исследования следует рассматривать в свете нескольких ограничений.Поперечный характер опроса затрудняет изучение временных отношений. Хотя неправильная классификация переменных воздействия, вероятно, будет минимальной, воздействие, измеренное во время обследования, может не отражать воздействие в прошлом, которое может быть важным для хронического состояния, такого как анемия. Предыдущие исследования показали, что место постоянного проживания в детстве может влиять на риск анемии больше, чем место текущего проживания, и полное влияние SES на здоровье не может быть зафиксировано путем измерения SES в определенный момент времени [32, 52, 53] .Наконец, большое количество отсутствующих наблюдений за ВИЧ и анемией могло внести систематическую ошибку в выборку. Уровень отклика среди городских мужчин был низким, что могло привести к смещению отбора, поскольку трудоустроенным мужчинам было сложно набирать на работу [27]. Чтобы свести к минимуму недостающие данные о ВИЧ, мы оценивали ВИЧ-статус из разных источников — тестирования из текущего или предыдущего исследования и самоотчета. Хотя маловероятно, что субъекты будут ложно сообщать о своем ВИЧ-положительном статусе, вполне возможно, что участники, не давшие согласия на тестирование на ВИЧ, с большей вероятностью будут ВИЧ-положительными.Таким образом, мы могли недооценить распространенность анемии и влияние ВИЧ. Кроме того, мы могли переоценить распространенность анемии, если бы у более здоровых участников с более низким риском анемии отсутствовали значения, потому что они были на работе и, следовательно, были недоступны для тестирования на ВИЧ. Также вероятно, что некоторые из женщин, которые сами сообщили, что они небеременны, на самом деле могли находиться на ранних сроках беременности, и это могло привести к переоценке распространенности анемии среди небеременных женщин и недооценке последствий беременности.Основным ограничением нашего исследования является отсутствие данных о других сопутствующих заболеваниях, вызывающих анемию. Основное наличие инфекционных (малярия, шистосомоз и анкилостомоз) и неинфекционных (хроническая болезнь почек, меноррагия, гастрит, язвенная болезнь) заболеваний, которые важны для этиологии анемии в популяциях SSA, могло привести к некоторым остаточным смешанным факторам. Например, поскольку беременные женщины подвергаются особому риску заражения малярией, некоторые из наблюдаемых эффектов беременности могут быть искажены малярией.Бремя некоторых хронических заболеваний, вызывающих анемию, таких как хроническая болезнь почек и язвенная болезнь, может быть выше среди городского населения и может способствовать наблюдаемому более высокому риску анемии среди городских женщин.
СУНДУК
Если вы не помните свой пароль, вы можете сбросить его, введя свой адрес электронной почты и нажав кнопку «Сбросить пароль».