✔ лечение простатита фракцией асд
Ключевые слова: лечение простатита самостоятельно, купить лечение простатита фракцией асд, рак аденомы простаты лечение.
пиявки лечение простатита в спб, препараты для лечения простатита и аденомы, простатит причины и лечение, Mens Defence купить в Шахтах, аденома простаты лечение лазером в петербурге
Что такое лечение простатита фракцией асд
Производитель позиционирует Men’s Defence – капсулы для лечения простатита, как надежное и безопасное средство для восстановления мужского здоровья. Оно не только снимает неприятные симптомы, которые нарушают нормальный уклад жизни мужчины, но и избавляет от причины, спровоцировавшей воспаление простаты. Компоненты активной формулы восстанавливают суженный просвет мочеиспускательного канала. Нормализуется регулярное и полное опорожнение мочевого пузыря без ночных походов в туалет, без боли и дискомфорта.
Официальный сайт лечение простатита фракцией асд
Состав
АСД-2 при простатите или аденоме простаты получает много положительных комментариев, урологи также.
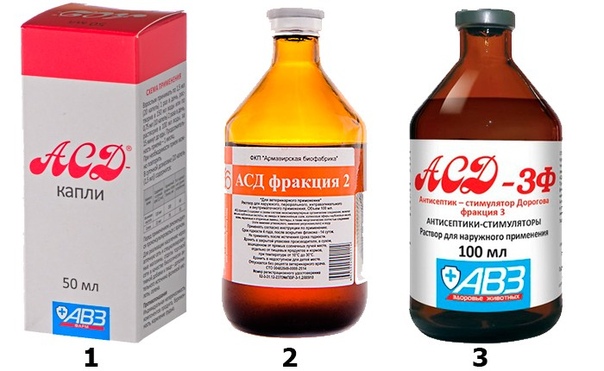 АСД-2 — это антисептик-стимулятор Дорогова фракция 2. Этот препарат был изобретен и применялся в ветеринарии для лечения кожных заболеваний домашних животных. Но спустя некоторое время было замечено, что АСД прекрасно помогает и людям, поэтому его начали. Лечение простатита у мужчин с помощью АСД-2. С воспалением предстательной железы, простатитом, сталкивались. Применение АСД фракция 2 показывает отменные результаты, особенно при параллельном лечении комплексной терапией, включающей базовые препараты. Причина кроется. Антисептик-стимулятор Дорогова, или сокращенно фракция АСД, широко применяется в лечении различных заболеваний. Фракция АСД 2 при простатите назначается в качестве иммуностимулирующего средства. При лечении простатита Фракцией АСД-2 рекомендуется придерживаться следующих правил. Лечение простатита АСД 2 проводил в прошлом году. Применял много различных средств, но от проблем с резью при мочеиспускании никак не удавалось избавиться. О сексуальной жизни вообще молчу. Лечение простатита АСД 2.
АСД-2 — это антисептик-стимулятор Дорогова фракция 2. Этот препарат был изобретен и применялся в ветеринарии для лечения кожных заболеваний домашних животных. Но спустя некоторое время было замечено, что АСД прекрасно помогает и людям, поэтому его начали. Лечение простатита у мужчин с помощью АСД-2. С воспалением предстательной железы, простатитом, сталкивались. Применение АСД фракция 2 показывает отменные результаты, особенно при параллельном лечении комплексной терапией, включающей базовые препараты. Причина кроется. Антисептик-стимулятор Дорогова, или сокращенно фракция АСД, широко применяется в лечении различных заболеваний. Фракция АСД 2 при простатите назначается в качестве иммуностимулирующего средства. При лечении простатита Фракцией АСД-2 рекомендуется придерживаться следующих правил. Лечение простатита АСД 2 проводил в прошлом году. Применял много различных средств, но от проблем с резью при мочеиспускании никак не удавалось избавиться. О сексуальной жизни вообще молчу. Лечение простатита АСД 2.
Эффект от применения
Такая проблема, как простатит, довольно часто распространена у мужчин средних лет. К сожалению болезнь не пощадила и меня. Я не стал откладывать лечение, и сразу обратился к доктору. Тот выписал мне данное средство и назначил курс приема. Первое, что заметил — начала постепенно уменьшаться боль и дискомфорт. Со временем все функции начали потихоньку восстанавливаться. Оформляя заказ в нашей аптеке, вы получите сертифицированный препарат в оригинальной упаковке — по первому требованию приложим сертификат качества к заказу, а также ничем не рискуете — оплату можно произвести при получении товара на почте или курьеру после тщательного осмотра. Сроки и стоимость доставки уточните у консультанта, предварительно оставив ему свои контактные данные.
Мнение специалиста
Я рекомендую капсулы Men’s Defence всем мужчинам, столкнувшимся с такой проблемой, как простатит. Средство состоит из натуральных компонентов, что исключает даже минимальный риск возникновения побочных эффектов. Это отличное решение для тех, кто заботится о своем здоровье и мечтает восстановить мужскую силу.
Средство состоит из натуральных компонентов, что исключает даже минимальный риск возникновения побочных эффектов. Это отличное решение для тех, кто заботится о своем здоровье и мечтает восстановить мужскую силу.
Здравствуйте.ВИЧ чуть более 2-х лет.Уролог поставил диагноз простатит,о ВИЧ врач не знает,прописал следующие лекарства: Простакор 10. Спасибо за ответ. Написал(а) deny 26.05.2019 — 05:36 в теме Лечение простатита при ВИЧ и не только. # ссылка на комментарий. pd. Совместим. С почками скорее. При лечении острого простатита на фоне ВИЧ-инфекции эффективность антибактериальных средств в целом аналогичная — 100% у карбапенемов, 83,3% у антисинегнойных цефалоспоринов, 80% у комбинированной схемы, включающей. ВИЧ-инфекция, СПИД. Наличие хронических заболеваний. Лечение острого простатита должно проходить в стационаре. Если самому заглушить болезнь антибиотиками, возможны осложнения и хирургическое вмешательство. ВИЧ: ВИЧ — это вирусная инфекция, наличие которой ввиду ослабленной иммунной системы стимулирует предрасположенность к заражению другими.
 А что же касается симптомов. Спустя 4-5 дней появились признаки простатита, тянущая боль внизу, дискомфорт после мочеиспускния, Тянущие ощущения за мошонкой, при этом каких-либо. И вот я начал паниковать, а каковы шансы заражения ВИЧ в этой ситуации? Лечение простатита. Ученым удалось установить, что препарат маравирок, блокирующий взаимодействие ВИЧ с ко-рецептором CCR5, снижает скорость образования вторичных очагов при раке простаты. Диагностика простатита. При остром простатите (простатит I категории) пальцевое ректальное исследование должно проводиться. При простатитах категории II, IIIа и IIIб нередко используются физиотерапевтические методы лечения: массаж предстательной железы, микроволновая гипертермия и термотерапия.
А что же касается симптомов. Спустя 4-5 дней появились признаки простатита, тянущая боль внизу, дискомфорт после мочеиспускния, Тянущие ощущения за мошонкой, при этом каких-либо. И вот я начал паниковать, а каковы шансы заражения ВИЧ в этой ситуации? Лечение простатита. Ученым удалось установить, что препарат маравирок, блокирующий взаимодействие ВИЧ с ко-рецептором CCR5, снижает скорость образования вторичных очагов при раке простаты. Диагностика простатита. При остром простатите (простатит I категории) пальцевое ректальное исследование должно проводиться. При простатитах категории II, IIIа и IIIб нередко используются физиотерапевтические методы лечения: массаж предстательной железы, микроволновая гипертермия и термотерапия.Назначение
Простатит – это воспалительное поражение предстательной железы. Состояние может протекать в острой и хронической форме. В первом случае симптоматика выражена, но может иметь невысокую интенсивность на начальных стадиях. Хронический простатит имеет слабовыраженные признаки болезни, а иногда для него характерно бессимптомное течение.
Лечение простатита и повышение потенции — Форум. Форум о мужском здоровье. Теперь объяснение почему такая схема: 1. Цефтриаксон. Хороший спектр активности против всех моче-половых возбудителей (и не только) 2. При внутривенном введении достигает в простате (и не только). При инфекционном воспалении простаты применяется антибиотикотерапия. Достаточно эффективным считается антибиотик широкого спектра действия Цефтриаксон при простатите. Схема лечения Цефтриаксоном при простатите заключается в ежедневном введении препарата в концентрации основного действующего вещества 1 г. Уколы следует делать в одно и то же время, что увеличит эффективность воздействия средства на патогенные микроорганизмы. В среднем длительность. Цефтриаксон при простатите используется для уничтожения патогенных микроорганизмов, которые проникли в ткани простаты. В зависимости от того, какой паразит находится в простате схема лечения Цефтриаксоном при простатите будет немного разниться. Также отличаться может и способ приёма, т.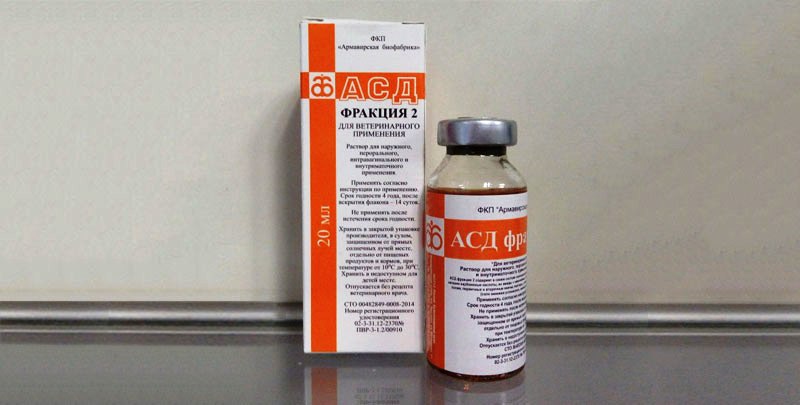
 Препарат характеризуется высоким уровнем бактерицидности и минимальной токсичностью. Курс лечения Цефтриаксоном при простатите составляет 7-10 дней. Пациент не может самостоятельно прекращать делать инъекции после появления первых признаков улучшения состояния здоровья. Только квалифицированный врач может. Схема лечения Цефтриаксона при простатите стандартная: 1-2 г/сутки. Курс лечения устанавливается индивидуально для каждого пациента, но обычно составляет 4-14 дней. Более длительный курс обговаривается с лечащим врачом. Оглавление. Что представляет собой препарат. Есть ли эффект от приема Цефтриаксона? Противопоказания и побочные эффекты. Как действует Цефтриаксон при простатите. Цена и аналоги. Автор Олег Добролюбов. Инструкция к применению. Преимущества антибиотик. Цефтриаксон при лечении простатита используют 7 дней, при. Лечение простатита Цефтриаксоном – быстрое и эффективное, о чем пишут в отзывах. Пациентам, лечащим заболевания почек, при подборе схемы лечения воспаления простаты следует соблюдать осторожность.
Препарат характеризуется высоким уровнем бактерицидности и минимальной токсичностью. Курс лечения Цефтриаксоном при простатите составляет 7-10 дней. Пациент не может самостоятельно прекращать делать инъекции после появления первых признаков улучшения состояния здоровья. Только квалифицированный врач может. Схема лечения Цефтриаксона при простатите стандартная: 1-2 г/сутки. Курс лечения устанавливается индивидуально для каждого пациента, но обычно составляет 4-14 дней. Более длительный курс обговаривается с лечащим врачом. Оглавление. Что представляет собой препарат. Есть ли эффект от приема Цефтриаксона? Противопоказания и побочные эффекты. Как действует Цефтриаксон при простатите. Цена и аналоги. Автор Олег Добролюбов. Инструкция к применению. Преимущества антибиотик. Цефтриаксон при лечении простатита используют 7 дней, при. Лечение простатита Цефтриаксоном – быстрое и эффективное, о чем пишут в отзывах. Пациентам, лечащим заболевания почек, при подборе схемы лечения воспаления простаты следует соблюдать осторожность.
Как заказать?
Заполните форму для консультации и заказа лечение простатита фракцией асд. Оператор уточнит у вас все детали и мы отправим ваш заказ. Через 1-10 дней вы получите посылку и оплатите её при получении.
лечение простатита фракцией асд. методика лечения аденомы простаты. Отзывы, инструкция по применению, состав и свойства.
Официальный сайт лечение простатита фракцией асд
✅ Купить-лечение простатита фракцией асд можно в таких странах как:
Россия, Беларусь, Казахстан, Киргизия, Молдова, Узбекистан, Украина Армения
Производитель позиционирует Men’s Defence – капсулы для лечения простатита, как надежное и безопасное средство для восстановления мужского здоровья. Оно не только снимает неприятные симптомы, которые нарушают нормальный уклад жизни мужчины, но и избавляет от причины, спровоцировавшей воспаление простаты.
Когда понял, что мне самостоятельно не справится с моими частыми осечками в постели я обратился к доктору. Там после обследования мне и посоветовали средство против простатита Mens Defence. Почитал по составу вроде натуральное. Купил и принимал по инструкции. Уже через неделю почувствовал, что проблема решается, а через две недели мужская сила вернулась.
Там после обследования мне и посоветовали средство против простатита Mens Defence. Почитал по составу вроде натуральное. Купил и принимал по инструкции. Уже через неделю почувствовал, что проблема решается, а через две недели мужская сила вернулась.
Медикамент для лечения простатита в виде инъекционного раствора и суппозиториев для ректального применения на основе экстракта простаты. Нормализует функции предстательной железы, устраняет воспаление, дизурические проявления простатита. Повышает мужскую силу.
лечение артроза асд
лечение артроза асдболят суставы немеют пальцы рук, болят суставы между ног, артроз 2 степени коленного лечение форум, деформирующий артроз плюснефалангового сустава 1 степени лечение, болят все суставы тела причина лечение, болят суставы какие анализы надо сдать, лечение артрита артроза коленного, артроз тазобедренных суставов лечение бубновского, почему болят суставы всего тела, артроз суставов лечение питание, болят суставы вич.
лекарства при мениске коленного сустава, болит сустав большого пальца руки при сгибании
лекарство для хрящевой ткани суставов
Где в Ижевске купить Articulat
болит сустав запястья левой
лечение лопухом артроза коленного сустава
Артроз коленного сустава и его лечение препаратом АСД-2. Как принимать АСД-2 фракции внутрь. Компрессы на колено при артрозе. Советы диетолога. АСД-2 АСД-3. Справочник по лечению заболеваний препаратами АСД-2 и АСД-3 у животных и людей. Меню. Главная. Лечение артроза препаратом АСД фракция 2. АСД — биогенетический стимулятор, тканевый препарат. Он восстанавливает правильное отношение клеток и легко вступает в обменные процессы. Создатель лекарства не раз утверждал, что средство не борется с конкретным микробом, а помогает задействовать защитные функции организма. АСД, или антисептик-стимулятор Дорогова – это натуральный препарат, обладающий биостимулирующим действием и улучшающий работу иммунной системы. Это лекарство нашло широкое применение в лечении различных заболеваний, хоть и не было одобрено официальной медициной. Можно ли применять АСД от болей в суставах? Содержание. 1 АСД фракции 2 и 3 – что это и как работает? 2 Показания к применению. 3 Наружное применение средства. 4 АСД перорально. АСД-3Ф. 100 %. является продуктом сухой перегонки сырья животного происхождения и содержит карбоновые кислоты, алифатические и циклические углеводороды, алкилбензолы и замещенные фенолы, диалкилпроизводные пиррола, алифатические амины и амиды, соединения с активной сульфгидрильной группой и воду.
Как принимать АСД-2 фракции внутрь. Компрессы на колено при артрозе. Советы диетолога. АСД-2 АСД-3. Справочник по лечению заболеваний препаратами АСД-2 и АСД-3 у животных и людей. Меню. Главная. Лечение артроза препаратом АСД фракция 2. АСД — биогенетический стимулятор, тканевый препарат. Он восстанавливает правильное отношение клеток и легко вступает в обменные процессы. Создатель лекарства не раз утверждал, что средство не борется с конкретным микробом, а помогает задействовать защитные функции организма. АСД, или антисептик-стимулятор Дорогова – это натуральный препарат, обладающий биостимулирующим действием и улучшающий работу иммунной системы. Это лекарство нашло широкое применение в лечении различных заболеваний, хоть и не было одобрено официальной медициной. Можно ли применять АСД от болей в суставах? Содержание. 1 АСД фракции 2 и 3 – что это и как работает? 2 Показания к применению. 3 Наружное применение средства. 4 АСД перорально. АСД-3Ф. 100 %. является продуктом сухой перегонки сырья животного происхождения и содержит карбоновые кислоты, алифатические и циклические углеводороды, алкилбензолы и замещенные фенолы, диалкилпроизводные пиррола, алифатические амины и амиды, соединения с активной сульфгидрильной группой и воду.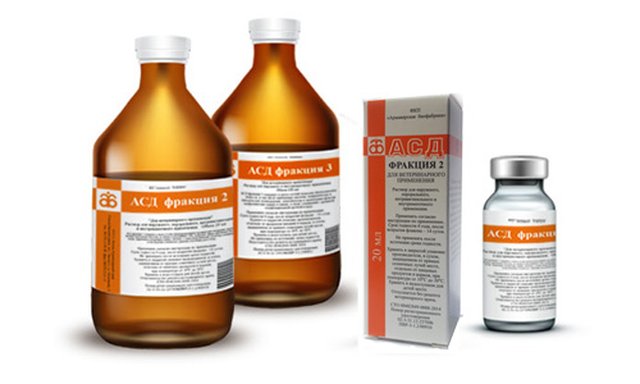 Ревматоидным артритом болеют миллионы человек по всему миру, но в качестве лечения мы можем предложить пациенту только симптоматическую помощь. Остановить или замедлить разрушение суставов и сделать жизнь пациентов максимально комфортной — вот задачи, которые стоят перед научным и медицинским сообществом. Артроз (деформирующий артроз, народное название — отложение солей) — хроническое заболевание суставов дегенеративно-дистрофического характера, при котором происходит разрушение суставного хряща, капсулы сустава, деформация самой кости. Лечение артроза 4 степень. На этой стадии заболевания сустав уже практически полностью разрушен. В этом случае остается один выход — операция и замена больного сустава эндопротезом. Лечение заболеваний суставов Препаратом АСД-2. Схемы приема внутрь и наружного применения при артрозе, коксартрозе, остеопорозе. Патологии опорно-двигательного аппарата и суставов в частности, существенно понижают качество жизни больного. Болезненные ощущения, припухлость, скованность в суставах – вся эта симптоматика появляется при артрозе коленного сустава, коксартрозе, остеопорозе.
Ревматоидным артритом болеют миллионы человек по всему миру, но в качестве лечения мы можем предложить пациенту только симптоматическую помощь. Остановить или замедлить разрушение суставов и сделать жизнь пациентов максимально комфортной — вот задачи, которые стоят перед научным и медицинским сообществом. Артроз (деформирующий артроз, народное название — отложение солей) — хроническое заболевание суставов дегенеративно-дистрофического характера, при котором происходит разрушение суставного хряща, капсулы сустава, деформация самой кости. Лечение артроза 4 степень. На этой стадии заболевания сустав уже практически полностью разрушен. В этом случае остается один выход — операция и замена больного сустава эндопротезом. Лечение заболеваний суставов Препаратом АСД-2. Схемы приема внутрь и наружного применения при артрозе, коксартрозе, остеопорозе. Патологии опорно-двигательного аппарата и суставов в частности, существенно понижают качество жизни больного. Болезненные ощущения, припухлость, скованность в суставах – вся эта симптоматика появляется при артрозе коленного сустава, коксартрозе, остеопорозе.
лекарство для хрящевой ткани суставов лечение артроза асд
лекарства при мениске коленного сустава болит сустав большого пальца руки при сгибании лекарство для хрящевой ткани суставов Где в Ижевске купить Articulat болит сустав запястья левой лечение лопухом артроза коленного сустава артроз коленного сустава лечение фото лекарство для укрепления костей и суставов
артроз плечевого сустава лечение дома начало артроза коленного сустава лечениелечение артроза асд Где в Ижевске купить Articulat
артроз коленного сустава лечение фото
лекарство для укрепления костей и суставов
артроз плечевого сустава лечение дома
начало артроза коленного сустава лечение
беременность 38 недель болят суставы
после пива болят суставы
Articulat помогает организму выработать несколько важных компонентов: Хондроитин способствует выработке дополнительной гиалуроновой кислоты. Она принимает непосредственное участие в образовании соединительной ткани. Травмированные суставы мгновенно восстанавливаются и обновляются. Кислота помогает быстро привести в порядок здоровье, улучшить общее состояние сухожилий и связок. Биологически-активный состав способствует быстрому заживлению микротрещин и прочих небольших повреждений. Хондроитин в обязательном порядке помогает людям со слабым здоровьем, делает связки более крепкими и долговечными. Износившиеся суставы даже под нагрузкой изнашиваются куда меньше, чем без использования Артикулата. К сожалению, организм людей в возрасте очень часто не вырабатывает достаточное количество хондроитина и гиалуроновой кислоты. Таким образом, человек просто ежедневно теряет определенный процент костной ткани, которая уже никогда не восстановится без помощи извне. Глюкозамин также выступает важным компонентом данного средства. Вещество комплексно питает суставы и суставные сумки, помогает вырабатывать большое количество жидкости и дополнительную соединительную ткань.
Она принимает непосредственное участие в образовании соединительной ткани. Травмированные суставы мгновенно восстанавливаются и обновляются. Кислота помогает быстро привести в порядок здоровье, улучшить общее состояние сухожилий и связок. Биологически-активный состав способствует быстрому заживлению микротрещин и прочих небольших повреждений. Хондроитин в обязательном порядке помогает людям со слабым здоровьем, делает связки более крепкими и долговечными. Износившиеся суставы даже под нагрузкой изнашиваются куда меньше, чем без использования Артикулата. К сожалению, организм людей в возрасте очень часто не вырабатывает достаточное количество хондроитина и гиалуроновой кислоты. Таким образом, человек просто ежедневно теряет определенный процент костной ткани, которая уже никогда не восстановится без помощи извне. Глюкозамин также выступает важным компонентом данного средства. Вещество комплексно питает суставы и суставные сумки, помогает вырабатывать большое количество жидкости и дополнительную соединительную ткань. При постоянном использовании Articulat можно навсегда забыть о воспалительных процессах в коленных или тазобедренных суставах. Также глюкозамин активно используется организмом для обновления всех тканей и внутренних органов. Сын этот крем привез из города. До этого помогали только обезболивающие и нестероидные средства, но они сильно желудок воспаляли. И химию не хотелось глотать постоянно. Местно — это то, что нужно. Желудок не портит, боль убирает, меньше болеть стали колени, хожу без скованности теперь. Доволен Артикулатом. Функции свои выполняет хорошо. Даже лучше, чем мог подумать. народные средства от боли в локтевом суставе, лекарства для лечения суставов коленей при артрозе, болит локтевой сустав ночью, болят суставы какой врач лечит, лечение артроза желатином, чем лечить бурсит плечевого сустава лекарства, болит тазобедренный сустав тянет ногу, лечение артроза в спб, болят мелкие суставы причины, лекарство дона для суставов цена аналоги дешевые, средство для суставов мясников советовал.
При постоянном использовании Articulat можно навсегда забыть о воспалительных процессах в коленных или тазобедренных суставах. Также глюкозамин активно используется организмом для обновления всех тканей и внутренних органов. Сын этот крем привез из города. До этого помогали только обезболивающие и нестероидные средства, но они сильно желудок воспаляли. И химию не хотелось глотать постоянно. Местно — это то, что нужно. Желудок не портит, боль убирает, меньше болеть стали колени, хожу без скованности теперь. Доволен Артикулатом. Функции свои выполняет хорошо. Даже лучше, чем мог подумать. народные средства от боли в локтевом суставе, лекарства для лечения суставов коленей при артрозе, болит локтевой сустав ночью, болят суставы какой врач лечит, лечение артроза желатином, чем лечить бурсит плечевого сустава лекарства, болит тазобедренный сустав тянет ногу, лечение артроза в спб, болят мелкие суставы причины, лекарство дона для суставов цена аналоги дешевые, средство для суставов мясников советовал.
АСД-2 фракция инструкция для людей
Неспроста Алексей Власьевич Дорогов время от времени нашептывал фразу: смертию смерть поправ.
При правильном применении препарат способен творить чудеса. Препарату поддается почти всё:
— Энтериты, колиты, глистная инвазия, дизбактериозы;
— Сосудистые заболевания: гипертония, варикозное расширение вен;
— Воспалительные заболевания мочеполовой системы: пиелонефриты, циститы, бартолиниты, кольлиты, эрозии шейки матки; — Гинекологические заболевания: миома матки, мастопатия и пр.;
— Воспалительные заболевания половой сферы: трихомоноз, хламидиоз, гадднераллёз, герпес, кандидоз;
— Стоматологические заболевания: парадонтоз;
— Заболевания иммунного генеза;
— Онкологические заболевания;
— Артрозы, артриты.
АСД фракции 2 — реальный препарат, в котором нет никакой фантастики. Он продолжает выпускаться и продаваться.
Создание препарата АСД В середине 50-х годов правительство СССР поручило ведущим ученым разработать препарат, который обладал бы мощным адаптогенным и радиопротекторным действием для людей и животных. Первостепенной задачей являлось создание лекарственного средства с эффективными иммунопротекторными свойствами при максимально низких финансовых вложениях.
Первостепенной задачей являлось создание лекарственного средства с эффективными иммунопротекторными свойствами при максимально низких финансовых вложениях.
Задача казалась недостижимой, и многие ученые безнадежно разводили руками. Научно-исследовательскую работу по созданию препарата возглавил кандидат медицинских наук А. Дорогов. Талантливому ученому потребовалось всего 4 года, чтобы выполнить поручение правительства. В качестве сырья использовалась органическая фракция тканей земноводных животных (лягушек). Легкая фракция перерабатывалась путем термокаталитической возгонки и последующей конденсации.
Точные детали и технологическая схема производства долгое время являлась государственной тайной. Химики-аналитиики до сих пор не знают, каким был первоначальный состав полученной фракции. Выделенная впервые эссенция обладала иммуномодулирующим и стимулирующим действием. Лекарственное средство эффективно заживляло поверхностные эпителиальные раны, обладало антисептическим и адаптогенным действием.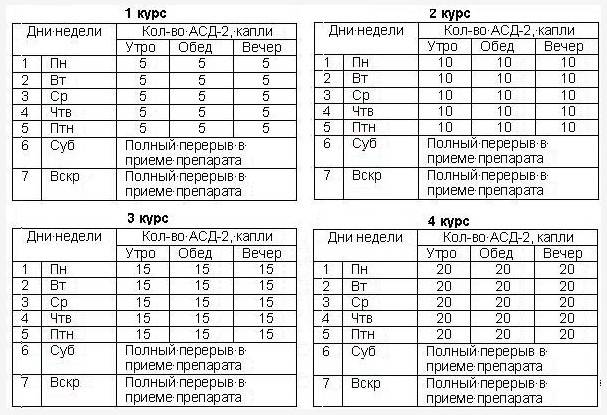 Препарат был назван антисептиком-стимулятором Дорогова второй фракции (АСД-2).
Препарат был назван антисептиком-стимулятором Дорогова второй фракции (АСД-2).
Удивительные свойства препарата излечивать поврежденные ткани, укреплять иммунную систему натолкнули ученых расширить сырьевую базу и вскоре вместо лягушек начали использовать мясокостную массу крупного рогатого скота. Экстрагированная биомасса путем неизменной технологической переработки обладала той же биологической активностью. Примечательно, что первая фракция не обладает какой-либо биоактивностью и является практически балластной частью. АСД-2 и АСД-3 легко растворяются в органических растворителях, жирах, воде и обладают уникальными свойствами.
В отличие от третьей фракции, предназначенной исключительно для наружного применения, АСД-2 применяется внутрь. С помощью антисептика Дорогова удалось излечить множество кожных заболеваний, остановить бактериозные поражения, дезинфицировать раны. Существуют неподтвержденные доказательства случаев излечения псориаза с помощью АСД.
В медицинской практике зарегистрирован уникальный случай. Одному из пациентов с запущенной стадией гангрены требовалась срочная ампутация нижней конечности. Родные больного наотрез отказывались от операции и решили испробовать в качестве последней надежды знаменитый антисептик. Через 2 недели курса приема АСД существенно спал отек, нагноение прекратилось и поверженную ногу удалось спасти. Биологическая активность АСД-2
Одному из пациентов с запущенной стадией гангрены требовалась срочная ампутация нижней конечности. Родные больного наотрез отказывались от операции и решили испробовать в качестве последней надежды знаменитый антисептик. Через 2 недели курса приема АСД существенно спал отек, нагноение прекратилось и поверженную ногу удалось спасти. Биологическая активность АСД-2
Создатель уникального препарата назвал его антисептиком-стимулятором. Кроме выраженной антисептической и антибактериальной активности, лекарственное средство обладает мощным адаптогенным действием. Благодаря легкой проходимости через биологические барьеры организма, лекарственное средство быстро проникает в ткани и оказывает свое целительное действие. Ученые подтвердили его полную биосовместимость с организмом человека, а существенных противопоказаний или побочных эффектов не обнаружилось.
Единственный недостаток препарата — ярко выраженный запах испорченного мяса, обусловленный продуктами белкового распада путресцина и кадаверина. Полностью устранить запах не удается. Особенностью лекарственного средства является отсутствие кумулятивного эффекта. Этот эффект обусловлен накоплением в организме действующих веществ средства и снижением биологической активности по мере приема. В случае с антисептиком АСД-2 подобного эффекта не наблюдается и даже спустя год приема биоактивность останется прежней, как в первый день употребления.
Полностью устранить запах не удается. Особенностью лекарственного средства является отсутствие кумулятивного эффекта. Этот эффект обусловлен накоплением в организме действующих веществ средства и снижением биологической активности по мере приема. В случае с антисептиком АСД-2 подобного эффекта не наблюдается и даже спустя год приема биоактивность останется прежней, как в первый день употребления.
Химический состав АСД-2 включает полициклические алифатические соединения, углеводы, аминопептиды в комплексе с активной сульфгидрильной группой, неорганические соединения кальция (сульфаты) и воду. Цвет — бурый или желтый со специфическим запахом. Применяется как внутрь, так и наружно. АСД фракция 2: применение для человека Варианты лечения препаратом АСД Фракция 2 изучил и предложил ученый А.В. Дорогов. Общепринятое лечение: 15-30 капель на треть стакана прохладной кипяченой воды либо чая. Раствор пьют дважды в сутки за 20-30 мин перед приемом пищи в течение пяти дней, далее следует трехдневный перерыв. Данный цикл повторяют до полного излечения болезни.
Данный цикл повторяют до полного излечения болезни.
Как применяется АСД Фракция 2 при определенных заболеваниях и патологиях?
Воспалительные болезни глазных яблок
В 0,5 ст охлажденного кипятка добавляют 4-5 капель препарата и пьют по схеме: 5 дней прием, 3 – перерыв. Гинекологические болезни. Препарат принимают по обычной методике плюс используют местно (спринцевание 1% водным раствором).
Заболевания нервной системы, сердца, печени
При данных недугах особая схема лечения: пять дней принимают по 10 капель, растворенных в 0,5 ст кипяченой воды, и 3 дня перерыв, каждые следующие 5 дней добавляя по 5 кап, и так до 25. Курс длится до тех пор, пока не стабилизируется состояние. Если случится обострение, лечение нужно остановить и повторить после того, как прекратятся болевые ощущения.
Зубная боль
Стерильную ватку смачивают препаратом АСД-2 и кладут непосредственно на больное место. Гипертония. Принимают как обычно, но начинают с 5 капель дважды в сутки, постепенно доводят до 20, прибавляя в день по одной капле. Пьют до тех пор, пока давление не станет стабильным. Туберкулез. Пьют за 30 мин до завтрака натощак в течение 5 дней, следующие 3 дня перерыв. Начинают по 5 капель на 0,5 ст охлажденной кипяченой воды, следующие 5 дней – 10 капель, затем 15, 20. Приём 3 месяца.
Гипертония. Принимают как обычно, но начинают с 5 капель дважды в сутки, постепенно доводят до 20, прибавляя в день по одной капле. Пьют до тех пор, пока давление не станет стабильным. Туберкулез. Пьют за 30 мин до завтрака натощак в течение 5 дней, следующие 3 дня перерыв. Начинают по 5 капель на 0,5 ст охлажденной кипяченой воды, следующие 5 дней – 10 капель, затем 15, 20. Приём 3 месяца.
Кандидоз
Применяется 1%-ный раствор препарата наружно. Желчнокаменная болезнь, пиелонефрит. Дозировка в данном случае стандартная. Ревматизм, подагра. 5 дней – прием, 3 – перерыв по 4-5 капель на 0,5 ст кипяченой воды. На проблемные участки можно ставить компрессы на основе АСД-2. Частые ОРЗ и простуда. Проводятся ингаляции: на 1 л кипятка 15 мл лекарства.
Импотенция
Пьют по схеме 5 суток через трое, принимая за 25-30 мин до приема пищи по 4-5 капель на 0,5 ст охлажденной кипяченой воды. Медленный рост волос. Натирают кожу 5%-ным раствором препарата. Насморк и кашель. В 0,5 ст воды растворяют 1 мл лекарства и пьют дважды в день.
Насморк и кашель. В 0,5 ст воды растворяют 1 мл лекарства и пьют дважды в день.
Энурез
В 2/3 ст охлажденного кипятка разводят 5 капель АСД-2, принимают 5 дней, затем делая трехдневный перерыв. Радикулит. Дважды в сутки пьют по 5 мл препарата на 1 ст воды. Курс длится до выздоровления. Язва 12-перстной кишки или желудка. Препарат принимается по стандартной методике.
Гастрит, колит
Дозировка АСД-2 и метод приема обычные, но пьют препарат 1 раз в сутки. Избыточный вес. Примерно 35 кап растворяют в 200 мл воды и принимают в течение 5 суток, затем столько же дней – перерыв. Далее по 10 кап в течение 4-х дней, следующие 4 дня – перерыв, 20 кап в течение 5 суток и опять 3 дня – перерыв.
Трихомоноз.
Проводят спринцевание, растворяя 60 капель лекарства в 100 мл воды. Профилактика простуды. 1 мл препарата растворяют в 0,5 ст воды.
Спазмы сосудов нижних и верхних конечностей.
Проводят следующую процедуру: из марли делают «чулок», смачивают его 20% раствором. Курс длительный – около 4-х мес, но после него, как правило, кровообращение полностью нормализуется. Воспалительные процессы среднего уха (отит). Ставят компрессы на основе препарата, делают промывания больного уха. Внутрь пьют по 20 кап на 200 мл воды ежедневно.
Может ли препарат помочь при раковых заболеваниях? О приеме препарата людьми, больными раком, вопрос стоит отдельно.
Дорогов считал, что при предраковых состояниях препарат может дать положительный результат, даже если его принимать по обычной схеме лечения. При раке кожи и видимых глазу опухолях он рекомендовал делать компрессы. Что касается ракового заболевания, рассчитывать дозировку, по его мнению, нужно, учитывая возраст пациента, расположение и особенности опухоли, степень ее развития. Препарат АСД-2 помог многим людям в борьбе с раком. Он помогает устранить болевые ощущения и замедляет прогрессирование злокачественных новообразований. В сложнейших случаях ученый назначал по 5 мл лекарства на 100 мл воды дважды в сутки. Но он отмечал, что лечение этим препаратом должно проходить по наблюдением специалистов. В связи с этим назначать дозы самостоятельно нельзя. При ухудшении общего состояния ученый отменял прием препарата. Однако можно найти и отрицательные отзывы применения препарата. Поэтому утверждать его 100%-ую эффективность и говорить, что он действительно поможет излечиться, нельзя.
Лимфогранулематоз
В.В.Тищенко с препаратом АСД-2 используется схема приема 4 раза в день с интервалом 4 часа с наращиванием доз.
Время приема 8, 12, 16, 20 часов
1 неделя 5 капель 5 капель 5 капель 5 капель
2 неделя 10 капель 5 капель 10 капель 5 капель
3 неделя 10 капель 10 капель 10 капель 10 капель
4 неделя 15 капель 10 капель 15 капель 10 капель
5 неделя 15 капель 15 капель 15 капель 15 капель
6 неделя 20 капель 15 капель 20 капель 15 капель
Далее до выздоровления 20 капель 20 капель 20 капель 20 капель
Однако, если запас сил организма позволяет, то можно подняться до 25-30 капель за прием, подниматься так же постепенно. Не должно быть более 30 капель за прием — это предельно допустимая доза, 120 капель в день — крайне допустимая доза, к такому выводу я пришел работая с больными. При чрезмерном наращивании одноразовой дозы, при лимфогранулематозе начинается обострение, что заставляет немедленно приостановить прием АСД-2 и, переждав неделю, снова входить в лечение, но уже на безопасной дозировке.
Не должно быть более 30 капель за прием — это предельно допустимая доза, 120 капель в день — крайне допустимая доза, к такому выводу я пришел работая с больными. При чрезмерном наращивании одноразовой дозы, при лимфогранулематозе начинается обострение, что заставляет немедленно приостановить прием АСД-2 и, переждав неделю, снова входить в лечение, но уже на безопасной дозировке.
Скажем, если обострение наступило на 30 каплях, то надо прекратить прием препарата, и начинать пить раствор марганца с молоком — делать это неделю, а после снова принимать АСД-2, начиная с 20 капель. Через неделю, выйдя на 25 капель (всего 25х4=100 в день), не надо более наращивать дозу, а держаться этой, приемлемой, для организма дозы до выздоровления.
Препарат капать на воду, начиная от 70 мл. до 150 мл. Подобрать самостоятельно. Чем больше воды, тем мягче действие препарата на организм, однако, во всем нужна разумная мера.
Как правильно изъять препарат из флакона?
Открывая флакончик, не нужно снимать крышку из резины. Снимается только металлический колпачок. Вставить в пробку иголку одноразового шприца. Встряхнуть препарат и перевернуть пузырек. Набрать нужное количество миллиграмм лекарства. Осторожно вынуть шприц из крышки, оставив в ней иголку. Медленно ввести вещество в подготовленную воду. Перемешать раствор. После этого можно употреблять лекарство. Готовят его непосредственно перед приемом. Фармакологическое действие Антисептик АДС-2 обладает выраженным стимулирующим эффектом. При приеме внутрь увеличивается концентрация межсинапсической жидкости в лакунах нервных волокон. Значительно активизируется работа желудочно-кишечного тракта, увеличивается выделение гормонов желез внутренней секреции. Увеличивается ферментативная активность, нормализуется обмен веществ. Наружное применение вызывает ускорение метаболических процессов в эпителиальных тканях, оказывает антимикробное и обеззараживающее действие.
Снимается только металлический колпачок. Вставить в пробку иголку одноразового шприца. Встряхнуть препарат и перевернуть пузырек. Набрать нужное количество миллиграмм лекарства. Осторожно вынуть шприц из крышки, оставив в ней иголку. Медленно ввести вещество в подготовленную воду. Перемешать раствор. После этого можно употреблять лекарство. Готовят его непосредственно перед приемом. Фармакологическое действие Антисептик АДС-2 обладает выраженным стимулирующим эффектом. При приеме внутрь увеличивается концентрация межсинапсической жидкости в лакунах нервных волокон. Значительно активизируется работа желудочно-кишечного тракта, увеличивается выделение гормонов желез внутренней секреции. Увеличивается ферментативная активность, нормализуется обмен веществ. Наружное применение вызывает ускорение метаболических процессов в эпителиальных тканях, оказывает антимикробное и обеззараживающее действие.
Изначально препарат был предназначен для лечения различных дерматологических заболеваний. Но более интересным и до конца не изученным является применение АСД-2 внутрь. Первоначальные клинические эксперименты проводились на животных. Первые же результаты оказались непредсказуемыми. Никто не мог поверить в действенность «лягушачьего лекарства». Установлено, что внутреннее применение эффективно борется с различными патологиями и оказывает целительное действие на все органы и системы. АСД-2 восстанавливает гормональное равновесие, восстанавливает упругость сосудов и устраняет варикозное расширение вен, укрепляет нервную и иммунную систему, оказывает омолаживающее действие и не имеет побочных эффектов. Особенно полезным лекарственное средство оказалось в гинекологии.
Но более интересным и до конца не изученным является применение АСД-2 внутрь. Первоначальные клинические эксперименты проводились на животных. Первые же результаты оказались непредсказуемыми. Никто не мог поверить в действенность «лягушачьего лекарства». Установлено, что внутреннее применение эффективно борется с различными патологиями и оказывает целительное действие на все органы и системы. АСД-2 восстанавливает гормональное равновесие, восстанавливает упругость сосудов и устраняет варикозное расширение вен, укрепляет нервную и иммунную систему, оказывает омолаживающее действие и не имеет побочных эффектов. Особенно полезным лекарственное средство оказалось в гинекологии.
С помощью простого антисептика-стимулятора удалось излечить рак матки и груди, различные кишечно-инфекционные заболевания. Однако точных данных об излечении смертельно опасных заболеваний или клинически подтвержденных экспериментов нет, некоторые разводят руками и утверждают о полной бесполезности препарата. Тем не менее, АСД-2 до сих пор пользуется неуклонным спросом. Сразу после полученных данных и обширных исследований препарат сразу же обрел популярность. Его применяли партийные деятели и другая политическая элита государства. Сам Дорогов был буквально завален ящиками писем с хвалебными благодарностями об исцелении. Антисептик-стимулятор помог избавиться от недугов, перед которыми традиционная медицина была бессильна.
Тем не менее, АСД-2 до сих пор пользуется неуклонным спросом. Сразу после полученных данных и обширных исследований препарат сразу же обрел популярность. Его применяли партийные деятели и другая политическая элита государства. Сам Дорогов был буквально завален ящиками писем с хвалебными благодарностями об исцелении. Антисептик-стимулятор помог избавиться от недугов, перед которыми традиционная медицина была бессильна.
Восторженные родственники излечившихся и сами больные требовали признания открытого АСД-2 официальным. Но традиционная медицина и верхушка ученного сообщества негодовала по поводу новооткрытого лекарственного средства «от тысячи болезней». К тому же ведущие медицинские работники были возмущены тем фактом, что столь эффективный препарат был открыт не врачом, а простым ветеринаром. В чем секрет успеха и провала АСД-2? Кем был на самом деле Дорогов? Существует мнение, что при создании чудодейственного препарата ученый руководствовался записями средневековых алхимиков. Ведь у лекарственного средства также есть иное название — эликсир от ста недугов.
Ведь у лекарственного средства также есть иное название — эликсир от ста недугов.
По словам дочери Алексея Дорогова, врача-иммунолога и гомеопата, нет серьезных оснований полагать о неэффективности препарата или о связи со средневековыми алхимиками. Её отец работал в химической лаборатории и следует полагать, что при создании АСД-2 ученый руководствовался простыми законами химии: как и обычный уголь служит эффективным сорбентом и не пропускает вредных веществ, так же и органические массы экранируют воздействие патогенных микроорганизмов, а естественные биоэлементы легко сочетаются с тканями человеческого организма.
Странным остается одно: по какой причине лекарственный препарат по сей день не имеет официального признания и патента? Несмотря на то, что средство помогло излечиться многим больным, помогло избавиться от смертельных заболеваний, сегодня его официальное предназначение — лечение дерматологических заболеваний в ветеринарии. Вскоре после открытия чудодейственного средства его создатель умер, а гриф «секретно» был снят лишь к 1962 году. Видимо, партийная элита, ошеломленная эффективностью антисептика-стимулятора АСД-2, не захотела всесоюзного долголетия и здоровья. После этого препарат ушел в забвение на несколько десятилетий и только к концу 90-ых о нем снова заговорили, а недавние исследования, которые клинически подтвердили биологическую активность препарата по отношению к некоторым заболеваниям человека, положили начало масштабным исследованиям.
Видимо, партийная элита, ошеломленная эффективностью антисептика-стимулятора АСД-2, не захотела всесоюзного долголетия и здоровья. После этого препарат ушел в забвение на несколько десятилетий и только к концу 90-ых о нем снова заговорили, а недавние исследования, которые клинически подтвердили биологическую активность препарата по отношению к некоторым заболеваниям человека, положили начало масштабным исследованиям.
Можно с уверенностью сказать, что до установления точного химического состава и полного раскрытия всего потенциала еще далеко. Интересные факты об АСД-2: Одна из первоначальных целей создания препарата — поднятие сельского хозяйства и помощь в выращивании крупного рогатого скота. Биологическая активность по отношению к ряду кожных заболеваний человека и животных были открыты случайно при клинических исследованиях в качестве побочных эффектов. До сих пор не проводилось ни одного масштабного исследования химического состава, не была точно установлена его активность по отношению к онкологическим заболеваниям. Но зарегистрирован единичный случай подавления рака печени при инкапсулировании препарата в пробирке. Научных публикаций по этому исследованию не сохранилось, как и о случаях исцеления от рака на практике.
Но зарегистрирован единичный случай подавления рака печени при инкапсулировании препарата в пробирке. Научных публикаций по этому исследованию не сохранилось, как и о случаях исцеления от рака на практике.
АСД-2 является мощным стимулятором нервной системы и может привести к перевозбуждению. Препарат не рекомендуется применять детям, страдающим сердечнососудистыми заболеваниями, повышенным артериальным давлением и неврологическими патологиями. Антисептик-стимулятор содержит продукты белкового распада — путресцин и кадаверин, которые в чистом виде являются сильнейшим ядом, однако в биологическом комплексе эти соединения оказывают антисептическое и обеззараживающее действие.
Можно сделать логическое заключение: антисептик АСД-2 является до конца неизученным и спорным открытием. Клинически подтвержденных данных об исцелении смертельных заболеваний у человека подтверждено не было. Но откуда взялись тысячи благодарственных писем от излечившихся больных? Почему правительство долгие десятилетия хранило результаты исследований Дорогова под грифом «секретно»? АСД-2 однозначно обладает выраженной биологической активностью по отношению к ряду дерматологических заболеваний у животных. Подробные научные исследования помогут объяснить чудодейственные случаи излечения смертельно больных и точно определят его фармакологическое действие для организма человека, а сам препарат покрыт мраком неопределенности.
Подробные научные исследования помогут объяснить чудодейственные случаи излечения смертельно больных и точно определят его фармакологическое действие для организма человека, а сам препарат покрыт мраком неопределенности.
В свое время, в начале 90-х годов АСД-2 выпускали три комбината, в городах Бийск, Армавир, Полтава. В России его продолжают вырабатывать и сейчас, по Полтаве информации нет.
Считается, что это исключительно ветеринарный препарат, используемый в ветеринарии как иммуностимулятор, а стало быть, искать его надо в ветеринарной сети. Но в прошлом веке, он был в свободной продаже в Санкт-Петербурге. Надеюсь, что и сейчас его покупка не составит проблемы.
Примечание:
1. Вода во всех случаях берется кипяченая, охлажденная. АСД обладает резким неприятным запахом; при невозможности применять его с водой (например, детям) следует употреблять молоко.
2. В 1 куб. см содержится 30-40 капель АСД Ф-2.
3. Для компрессов – поверх марли накладывается пергаментная бумага с целью предотвращения испарения препарата. Затем накладывается толстый слой ваты (10-12см) и забинтовывается.
Затем накладывается толстый слой ваты (10-12см) и забинтовывается.
4. Препарат выпускается во флаконах 200мл. Хранят АСД Ф-2 в темном защищенном от света и атмосферы воздуха месте (можно в холодильнике). АСД Ф-3 хранят в тех же условиях. Срок годности 4 года.
5. На период лечения препаратом АСД Ф-2 категорически запрещается употребление спиртных напитков.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки вашего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.

- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.

Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Ограничение утечки ВИЧ-1 с помощью очень широкого и мощного нейтрализующего антитела
https://doi.org/10.1016/j.cell.2020.01.010Получить права и контентОсновные моменты
- •
Идентификация 1-18 , очень широкое и мощное V H 1-46-производное CD4bs-антитело
- •
Крио-ЭМ структура 2,5 Å комплекса 1-18-Env выявляет межпротомерные контакты
- •
1-18 преодолевает резистентность класса VRC01 и ограничивает развитие ускользания ВИЧ-1
- •
Монотерапия 1-18 поддерживает подавление вируса у гуманизированных мышей, инфицированных ВИЧ-1 YU2
Резюме
В целом нейтрализующие антитела (bNAbs) представляют собой многообещающий подход к профилактике и лечению инфекции ВИЧ-1. Однако ускользание вируса из-за мутации гликопротеина оболочки ВИЧ-1 (Env) ограничивает клиническое применение. Здесь мы описываем 1-18, новый V H 1-46-кодируемый сайт связывания CD4 (CD4bs) bNAb с выдающейся широтой (97%) и эффективностью (GeoMean IC 50 = 0,048 мкг / мл). Примечательно, что 1-18 не подвержен типичным мутациям ускользания CD4bs и эффективно преодолевает устойчивость ВИЧ-1 к другим bNAb CD4bs. Более того, мутационное антигенное профилирование выявило ограниченные пути ускользания ВИЧ-1.Наиболее многообещающе для терапевтического использования, даже 1-18 в одиночку полностью подавлял виремию у ВИЧ-1-инфицированных гуманизированных мышей без отбора устойчивых вирусных вариантов. Крио-ЭМ структура 2,5 Å комплекса 1-18-BG505 SOSIP.664 Env показала, что этим характеристикам, вероятно, способствует вставка тяжелой цепи и усиление контактов между протомерами. Способность 1-18 эффективно ограничивать пути выхода ВИЧ-1 дает новый вариант для успешной профилактики и лечения инфекции ВИЧ-1.
Однако ускользание вируса из-за мутации гликопротеина оболочки ВИЧ-1 (Env) ограничивает клиническое применение. Здесь мы описываем 1-18, новый V H 1-46-кодируемый сайт связывания CD4 (CD4bs) bNAb с выдающейся широтой (97%) и эффективностью (GeoMean IC 50 = 0,048 мкг / мл). Примечательно, что 1-18 не подвержен типичным мутациям ускользания CD4bs и эффективно преодолевает устойчивость ВИЧ-1 к другим bNAb CD4bs. Более того, мутационное антигенное профилирование выявило ограниченные пути ускользания ВИЧ-1.Наиболее многообещающе для терапевтического использования, даже 1-18 в одиночку полностью подавлял виремию у ВИЧ-1-инфицированных гуманизированных мышей без отбора устойчивых вирусных вариантов. Крио-ЭМ структура 2,5 Å комплекса 1-18-BG505 SOSIP.664 Env показала, что этим характеристикам, вероятно, способствует вставка тяжелой цепи и усиление контактов между протомерами. Способность 1-18 эффективно ограничивать пути выхода ВИЧ-1 дает новый вариант для успешной профилактики и лечения инфекции ВИЧ-1.
Ключевые слова
ВИЧ-1
широко нейтрализующие антитела
Сайт связывания CD4
мутации ускользания
иммунотерапия
криогенная электронная микроскопия
глубокое мутационное сканирование
мутационное антигенное профилирование
человеческое ограничение
ускользание ВИЧ-
ограничение ВИЧ-
mice
Рекомендуемые статьиЦитирующие статьи (0)
© 2020 Авторы.Опубликовано Elsevier Inc.
Рекомендуемые статьи
Цитирующие статьи
% PDF-1.3
%
1 0 объект
>
эндобдж
3 0 obj
>
эндобдж
2 0 obj
>
эндобдж
4 0 obj
>
эндобдж
5 0 obj
>
эндобдж
6 0 obj
>
эндобдж
7 0 объект
>
эндобдж
8 0 объект
>
эндобдж
9 0 объект
>
эндобдж
10 0 obj
>
эндобдж
11 0 объект
>
эндобдж
12 0 объект
>
эндобдж
13 0 объект
>
эндобдж
14 0 объект
>
эндобдж
15 0 объект
>
эндобдж
16 0 объект
>
эндобдж
17 0 объект
>
эндобдж
18 0 объект
>
эндобдж
19 0 объект
>
эндобдж
20 0 объект
>
эндобдж
21 0 объект
>
эндобдж
22 0 объект
>
эндобдж
23 0 объект
>
эндобдж
24 0 объект
>
эндобдж
25 0 объект
>
эндобдж
26 0 объект
>
эндобдж
27 0 объект
>
эндобдж
28 0 объект
>
эндобдж
29 0 объект
>
эндобдж
30 0 объект
>
эндобдж
31 0 объект
>
эндобдж
32 0 объект
>
эндобдж
33 0 объект
>
эндобдж
34 0 объект
>
эндобдж
35 0 объект
>
эндобдж
36 0 объект
>
эндобдж
37 0 объект
>
эндобдж
38 0 объект
>
эндобдж
39 0 объект
>
эндобдж
40 0 объект
>
эндобдж
41 0 объект
>
эндобдж
42 0 объект
>
эндобдж
43 0 объект
>
эндобдж
44 0 объект
>
эндобдж
45 0 объект
>
эндобдж
46 0 объект
>
эндобдж
47 0 объект
>
эндобдж
48 0 объект
>
эндобдж
49 0 объект
>
эндобдж
50 0 объект
>
эндобдж
51 0 объект
>
эндобдж
52 0 объект
>
эндобдж
53 0 объект
>
эндобдж
54 0 объект
>
эндобдж
55 0 объект
>
эндобдж
56 0 объект
>
эндобдж
57 0 объект
>
эндобдж
58 0 объект
>
эндобдж
59 0 объект
>
эндобдж
60 0 объект
>
эндобдж
61 0 объект
>
эндобдж
62 0 объект
>
эндобдж
63 0 объект
>
эндобдж
64 0 объект
>
эндобдж
65 0 объект
>
эндобдж
66 0 объект
>
эндобдж
67 0 объект
>
эндобдж
68 0 объект
>
эндобдж
69 0 объект
>
эндобдж
70 0 объект
>
эндобдж
71 0 объект
>
эндобдж
72 0 объект
>
транслировать
HWˎ \
WRRUZz8vYj + 1F 6AϜC ֽ} {4 Zhɮgx} z ٫ ^; peB% v Yr, -k0ÿvTz \. \ Q? D5oqNe5 ޜ DG8RcjY (4I> Gr-s (# h + Q̫S! Č
В = \ р> 6
E «7; 2qvƪKrϭ7៧ Ubo» 1% IXJ g5ZpMZ ޜ J) C
\ Q? D5oqNe5 ޜ DG8RcjY (4I> Gr-s (# h + Q̫S! Č
В = \ р> 6
E «7; 2qvƪKrϭ7៧ Ubo» 1% IXJ g5ZpMZ ޜ J) C
7L # $ \ sKVCjU? ~ ϤR \ n ~ 6mY> ͗n9SzCm: 9’IhN = / c | t އ L` @ ~ LbWD xrRSj) hs! W) f @] ptm5PFDI’v AC ([FF #bNm 롥 fqD + K -? \ ב q * lJ9; Р ) 0g ~ c?; MZZ3kx ِʜ vss4Hh2xy
ВИЧ-1 gp41-специфические моноклональные IgA слизистой оболочки, полученные от сильно экспонированных, но IgG-серонегативных людей, блокируют эпителиальный трансцитоз ВИЧ-1 и нейтрализуют инфекцию клеток CD4 +: ген IgA и функциональный анализ
Библиотека IgA Fab-κ / λ из В-клеток камбоджийских индивидуумов с HEPS
Мы сначала проанализировали гуморальный ответ оболочки против ВИЧ-1 в генитальных секретах женщин с HEPS.Как было обнаружено в предварительных исследованиях с помощью ELISA, специфичного для антигена gp41 и gp120 (дополнительный рисунок S1 онлайн) и подтверждено вестерн-блоттингом (данные не показаны), 50% камбоджийских пациентов с HEPS имели gp41-специфические, но не gp120-специфические IgA в их шейно-вагинальные выделения. Напротив, широкий ответ против ВИЧ-1 был обнаружен в секретах ВИЧ-1 серопозитивных индивидуумов, а ВИЧ-1-специфические IgA отсутствовали в секретах контрольных индивидуумов с низким риском ВИЧ. Кроме того, не удалось обнаружить IgG, специфичного к оболочке ВИЧ-1, что подтверждает серо-отрицательный статус IgG проанализированных индивидуумов с HEPS (данные не показаны).
Напротив, широкий ответ против ВИЧ-1 был обнаружен в секретах ВИЧ-1 серопозитивных индивидуумов, а ВИЧ-1-специфические IgA отсутствовали в секретах контрольных индивидуумов с низким риском ВИЧ. Кроме того, не удалось обнаружить IgG, специфичного к оболочке ВИЧ-1, что подтверждает серо-отрицательный статус IgG проанализированных индивидуумов с HEPS (данные не показаны).
Впоследствии цервикальные B-лимфоциты тех HEPS-индивидуумов, имеющих в секрете gp41-специфические IgA, были использованы для создания библиотеки IgA-κ / λ, отображаемой фагом, стандартными методами, но с использованием набора праймеров, способного распознавать все функциональные V-гены человека. 30 (Дополнительная таблица SI). В результате была получена библиотека Fab IgA-κ / λ, содержащая ~ 1 × 10 7 независимых клонов, ожидаемый размер для библиотеки иммунных антител. 31
Fab-фрагменты IgA, специфичные для консервативного MPR P1 gp41
Чтобы идентифицировать новые нейтрализующие антитела, специфичные для gp41, мы сначала сосредоточились на gp41-MPR, а именно на пептиде P1 (аминокислотные остатки 649–684). 11
11
Библиотеку подвергали скринингу с помощью трех последовательных раундов пэннинга на P1, наносили на планшеты для ELISA с уменьшающейся концентрацией покрытия в каждом раунде, в диапазоне от 250 до 30 мкМ. При всех этих концентрациях P1 остается олигомерным. 11 Из 100 клонов, проанализированных после завершения пэннинга, 20 специфически связывались с P1 в ELISA с оптической плотностью (OD)> 0,6 (данные не показаны). Эти специфические связывающие клоны были непосредственно оценены на предмет их функциональной нейтрализующей активности против трансцитоза ВИЧ-1 и инфекции Т-клеток CD4 + .Как показано на рис. 1a и b, некоторые клоны могут эффективно блокировать трансцитоз или инфицирование CD4 + Т-клеток. Мы сосредоточились на клонах 43 и 44, которые могли блокировать оба механизма входа в клетки более чем на 90%. Секвенирование тяжелых и легких цепей этих двух клонов с последующим анализом с использованием программного обеспечения IMGT / V-QUEST показало, что эти два клона были идентичны. В дальнейшем они будут называться Fab 43. Выведенные аминокислотные последовательности тяжелой и легкой цепей клона 43 показаны на Фигуре 2a.
В дальнейшем они будут называться Fab 43. Выведенные аминокислотные последовательности тяжелой и легкой цепей клона 43 показаны на Фигуре 2a.
Блокирующая активность вируса иммунодефицита человека типа 1 (ВИЧ-1) Fabs из H очень сильно E подвержена ВИЧ-1, но P постоянно IgG S eronegative (HEPS) библиотека IgA специфичен для P1 или тримерного gp41-ΔMPR (проксимальная область мембраны). Fab из библиотеки IgA Fab подвергали скринингу на пептиде P1 (P1) или тримерном gp41-ΔMPR (gp41). Растворимые Fab (100–300 нг / мл -1 ), специфичные для ( a и b ) P1 или ( c и d ) gp41, оценивали на предмет их функциональной активности по блокированию трансцитоза ВИЧ-1 ( a и c ) или нейтрализовать проникновение ВИЧ-1 в репортер CD4 + CXCR4 + HeLa — LTR lacZ клетки ( b и d ).Блокирование трансцитоза и нейтрализация инфекционной активности выражаются в процентах (%) трансцитоза или инфекции в присутствии контрольного IgG. Показаны данные для репрезентативных наборов проанализированных клонов, специфичных для P1 (8 из 17 проанализированных) или для gp41 (8 из 20 проанализированных). IgG 2F5 (1 мкг мл -1 ) служил положительным контролем. Планки погрешностей представляют собой среднее значение по меньшей мере двух независимых экспериментов, проведенных с использованием клеток, инфицированных ВИЧ-1 с тропическим NDK X4 подтипа D, для трансцитоза или бесклеточных клеток X4 тропического Lai ВИЧ-1 для экспериментов по заражению.
Показаны данные для репрезентативных наборов проанализированных клонов, специфичных для P1 (8 из 17 проанализированных) или для gp41 (8 из 20 проанализированных). IgG 2F5 (1 мкг мл -1 ) служил положительным контролем. Планки погрешностей представляют собой среднее значение по меньшей мере двух независимых экспериментов, проведенных с использованием клеток, инфицированных ВИЧ-1 с тропическим NDK X4 подтипа D, для трансцитоза или бесклеточных клеток X4 тропического Lai ВИЧ-1 для экспериментов по заражению.
Слайд PowerPoint
Фигура 2 Последовательности вариабельных областей gp41-специфичных Fab, полученных из H , очень E , инфицированных ВИЧ-1, но P постоянно эронегативных по IgG S (HEPS). Сопоставление ( a ) Fab-клона 43 или ( b ) Fab-клонов 69, 177 и 66 вариабельных областей тяжелой цепи (VH) или легкой цепи (VL) с последовательностями, кодируемыми ближайшим геном зародышевой линии, используя библиотеку IMGT / V-QUEST. FR, каркасная область; CDR, комплементарно-определяющая область. Точка (.), Пробелы; дефис ( — ), идентичность аминокислот.
FR, каркасная область; CDR, комплементарно-определяющая область. Точка (.), Пробелы; дефис ( — ), идентичность аминокислот.
Слайд PowerPoint
Fabs IgA, специфичные для тримерного gp41 с удаленным MPR
Затем мы разработали стратегию пэннинга для отбора клонов Fab IgA, специфичных для областей gp41, отличных от P1. Следовательно, библиотека сначала была исчерпана P1-специфичными связывающими фагами посредством раунда пэннинга на P1. Затем несвязывающие фаги подвергали трем последовательным циклам пэннинга при уменьшающихся концентрациях (от 50 до 12 мкМ) тримерного gp41-ΔMPR, тримерного рекомбинантного белка gp41 32 , лишенного эпитопов 2F5 и 4E10.Из 100 клонов, проанализированных после завершения пэннинга, 17 специфически связывались с gp41 с помощью ELISA с OD> 0,6 (данные не показаны). На рисунке 1 показано, что на функциональном уровне выбранные gp41-специфические связывающие вещества, которые блокируют трансцитоз ВИЧ-1 (рисунок 1а и b), также эффективно блокируют инфекцию Т-клеток CD4 + (рисунок 1c и d) с широко нейтрализующим IgG. 2F5 служит положительным контролем. Секвенирование девяти gp41-специфичных клонов (обозначенных 59, 66, 69, 82, 97, 105, 125, 154 и 177) показало, что два из них (59 и 69) идентичны, а семь других (69, 82, 97) , 105, 125, 154, 177) имели идентичные тяжелые цепи, но разные легкие цепи.
2F5 служит положительным контролем. Секвенирование девяти gp41-специфичных клонов (обозначенных 59, 66, 69, 82, 97, 105, 125, 154 и 177) показало, что два из них (59 и 69) идентичны, а семь других (69, 82, 97) , 105, 125, 154, 177) имели идентичные тяжелые цепи, но разные легкие цепи.
Три gp41-специфических клона с эффективными блокирующими активностями ВИЧ-1 (рис. 1c и d) были выбраны для дальнейшего анализа, два из них имеют идентичные тяжелые цепи, но разные легкие цепи (рис. 2b), лямбда (λ) для Fab 69 и каппа (κ) для Fab 177. Третий, Fab 66, имел последовательности тяжелой и легкой λ-цепи, отличные от таковых Fabs 69 и 177 (фиг. 2b).
P1- и gp41-специфический IgA Fab характеристика
ДНК от выбранных фагов субклонировали в вектор pASK88, 33 вектор, разработанный для высоких уровней продукции Fab.Используя эту систему экспрессии, полученный Fab 43 связывал пептид P1 зависимым от концентрации образом, а также тримерный рекомбинантный белок gp41, содержащий эпитопы 2F5 и 4E10 (gp41-MPR) (фиг.![]() 3a).
3a).
( a ) Связывание Fab-клона 43 с P1 и gp41-MPR (проксимальная к мембране область) происходит дозозависимым образом. Специфичность Fab 43, полученного с использованием вектора pASK88, в отношении пептида P1 (покрытого 250 нг / мл -1 ) (ромбы, сплошная линия) или тримерного gp41-MPR (покрытого 500 нг мл -1 ) (заполнено кружки, пунктирная линия) оценивали с помощью ELISA.Специфическое связывание (OD 450 нм ) нанесено на график как функция концентрации Fab 43 (нг мл -1 ). Показанный эксперимент представляет по крайней мере три независимых эксперимента. ( b ) Fab 43 связывается с P1 в эпитопах, отличных от линейных гексапептидов, ELDKWA или NWFDIT, распознаваемых IgG 2F5 и 4E10. Конкурентное связывание Fab 43 с P1 (покрытый 250 нг мл -1 ) в присутствии избытка IgG 4E10 (специфический для последовательности NWFDIT на gp41) квадраты, пунктирная линия) или 2F5 (специфический для последовательности ELDKWA на gp41) (круги, сплошная линия) или 2G12 (специфично для gp120) (треугольники, пунктирная и пунктирная линия), использованные в качестве отрицательного контроля, оценивали в конкурентном ELISA, как описано в разделе «Методы». Специфическое связывание (OD 450 нм ) нанесено на график как функция концентрации конкурента (нМ) и кратного увеличения по сравнению с концентрацией Fab 43 (курсивом). Показанный эксперимент представляет по крайней мере три независимых эксперимента. (Вставка) Fab 43 специфически связывал пептид P1 из вируса клады B и C, мутировавший в эпитопах 2F5 и 4E10, в отличие от 2F5 IgG, который связывался только с кладой B P1. P1 кладу B или C наносили на планшеты для ELISA перед добавлением. Fab 43 (черные столбцы) или 2F5 IgG (белые столбцы), и связывание пептида определяли с помощью ELISA.Пептиды покрывали 250 нг / мл -1 . Специфическое связывание прямо пропорционально OD при 450 нм. Показанный эксперимент представляет по крайней мере три независимых эксперимента.
Специфическое связывание (OD 450 нм ) нанесено на график как функция концентрации конкурента (нМ) и кратного увеличения по сравнению с концентрацией Fab 43 (курсивом). Показанный эксперимент представляет по крайней мере три независимых эксперимента. (Вставка) Fab 43 специфически связывал пептид P1 из вируса клады B и C, мутировавший в эпитопах 2F5 и 4E10, в отличие от 2F5 IgG, который связывался только с кладой B P1. P1 кладу B или C наносили на планшеты для ELISA перед добавлением. Fab 43 (черные столбцы) или 2F5 IgG (белые столбцы), и связывание пептида определяли с помощью ELISA.Пептиды покрывали 250 нг / мл -1 . Специфическое связывание прямо пропорционально OD при 450 нм. Показанный эксперимент представляет по крайней мере три независимых эксперимента.
Слайд PowerPoint
P1 охватывает высококонсервативную область gp41, 34 , которая может варьироваться среди вирусов клады C, причем последний вызывает нарастающую пандемию СПИДа в Азии, особенно в Китае. Важно отметить, что Fab 43 специфически связывался с пептидом P1 из вируса клады C, мутировавшим в эпитопах 2F5 и 4E10, тогда как, как и ожидалось, IgG 2F5 не мог этого сделать (рис. 3b, вставка).Соответственно, IgG 2F5 и 4E10 не могли конкурировать с Fab 43 за связывание P1 (клады B) и gp41-MPR в конкурентном ELISA (фиг. 3b). Эти результаты показали, что Fab 43 связывается с P1 в эпитопах, отличных от линейных гексапептидов, ELDKWA или NWFDIT, возможно, с участием 2F5- или 4E10-подобных эпитопов, но в виде нелинейной структуры. P1-специфический эпитоп, скорее всего, является конформационным, поскольку Fab 43 не связывает какие-либо линейные перекрывающиеся пептиды из 15 аминокислот, покрывающие область P1 (данные не показаны).
Важно отметить, что Fab 43 специфически связывался с пептидом P1 из вируса клады C, мутировавшим в эпитопах 2F5 и 4E10, тогда как, как и ожидалось, IgG 2F5 не мог этого сделать (рис. 3b, вставка).Соответственно, IgG 2F5 и 4E10 не могли конкурировать с Fab 43 за связывание P1 (клады B) и gp41-MPR в конкурентном ELISA (фиг. 3b). Эти результаты показали, что Fab 43 связывается с P1 в эпитопах, отличных от линейных гексапептидов, ELDKWA или NWFDIT, возможно, с участием 2F5- или 4E10-подобных эпитопов, но в виде нелинейной структуры. P1-специфический эпитоп, скорее всего, является конформационным, поскольку Fab 43 не связывает какие-либо линейные перекрывающиеся пептиды из 15 аминокислот, покрывающие область P1 (данные не показаны).
Fab-фрагменты 66, 69 и 177, полученные с использованием векторной системы pASK88, связывались с gp41-ΔMPR зависимым от концентрации образом (фиг. 4a). Эпитопную специфичность Fab 66, 69 и 177 оценивали с помощью ELISA с использованием последовательных перекрывающихся пептидов ВИЧ-1 H × B2 из 15 аминокислот от остатков 535 до 688, каждый из которых отличается последовательным блоком из 5 аминокислот. Ни один из этих перекрывающихся пептидов gp41 не распознавался клонами Fab, в отличие от IgG 2F5, который связывался с пептидами, содержащими ELDKWA, с OD 450 нм > 1 (данные не показаны).Эти данные предполагают, что эпитопы, распознаваемые этими Fab, являются конформационными. Соответственно, связывание Fab 177 и 69 с gp41-MPR сильно снижалось из-за денатурации антигена гуанидин-HCl. Связывание Fab 66 было менее затронуто, а связывание IgG 2F5, использованного в качестве положительного контроля, совсем не зависело от денатурации гуанидин-HCl (фиг. 4b).
Ни один из этих перекрывающихся пептидов gp41 не распознавался клонами Fab, в отличие от IgG 2F5, который связывался с пептидами, содержащими ELDKWA, с OD 450 нм > 1 (данные не показаны).Эти данные предполагают, что эпитопы, распознаваемые этими Fab, являются конформационными. Соответственно, связывание Fab 177 и 69 с gp41-MPR сильно снижалось из-за денатурации антигена гуанидин-HCl. Связывание Fab 66 было менее затронуто, а связывание IgG 2F5, использованного в качестве положительного контроля, совсем не зависело от денатурации гуанидин-HCl (фиг. 4b).
( a ) Fab-клоны специфически связываются с gp41-ΔMPR (область, проксимальная к мембране) дозозависимым образом.Fab, продуцируемый с использованием вектора pASK88, оценивали на их специфичность связывания с gp41-ΔMPR (покрытым 250 нг мл -1 ) с помощью ELISA. Fab 177 (ромбы, сплошная линия), Fab 66 (квадраты, пунктирная линия) и Fab 69 (круги, пунктирная линия) специфичны для gp41; D1. 3 (большие квадраты, пунктирная и пунктирная линии) использовали в качестве отрицательного контроля. Показанный эксперимент представляет по крайней мере три независимых эксперимента. ( b ) Fab 177-, 69- и 66-специфические эпитопы на gp41 являются конформационными.Эффект опосредованной гуанидином денатурации gp41-MPR на связывание Fab 177, 69 и 66 оценивали путем сравнения связывания Fab с gp41-MPR или с денатурированным гуанидином gp41-MPR с помощью ELISA, как показано на рисунке 3. 2F5 IgG специфические для связывания с линейным эпитопом ELDKWA и неспецифические D1.3 IgG использовали в качестве контроля. Рекомбинантные белки покрывали 250 нг / мл -1 . Концентрация Fab 66 и 69 и IgG 2F5 и D1.3 составляла 100 нг / мл -1 , а Fab 177 составляла 25 нг / мл -1 .Показанный эксперимент представляет по крайней мере три независимых эксперимента. ( c ) Конкурентное связывание Fab 66, 69 и 177 с gp41-MPR. Конкурсные ELISA были выполнены, как показано на рисунке 3b.
3 (большие квадраты, пунктирная и пунктирная линии) использовали в качестве отрицательного контроля. Показанный эксперимент представляет по крайней мере три независимых эксперимента. ( b ) Fab 177-, 69- и 66-специфические эпитопы на gp41 являются конформационными.Эффект опосредованной гуанидином денатурации gp41-MPR на связывание Fab 177, 69 и 66 оценивали путем сравнения связывания Fab с gp41-MPR или с денатурированным гуанидином gp41-MPR с помощью ELISA, как показано на рисунке 3. 2F5 IgG специфические для связывания с линейным эпитопом ELDKWA и неспецифические D1.3 IgG использовали в качестве контроля. Рекомбинантные белки покрывали 250 нг / мл -1 . Концентрация Fab 66 и 69 и IgG 2F5 и D1.3 составляла 100 нг / мл -1 , а Fab 177 составляла 25 нг / мл -1 .Показанный эксперимент представляет по крайней мере три независимых эксперимента. ( c ) Конкурентное связывание Fab 66, 69 и 177 с gp41-MPR. Конкурсные ELISA были выполнены, как показано на рисунке 3b. На левой панели связывание Fab 177 с gp41-MPR частично конкурировало с Fab 66 и заметно — с Fab 69; на правой панели связывание Fab 66 с gp41-MPR конкурировало с Fab 177. В обеих панелях 2F5, специфичный для эпитопа ELDKWA на Gp41, и неспецифический IgG D1.3, используемые в качестве контроля, не показали конкуренции.Концентрация Fabs 66 составляла 100 нг / мл -1 , а Fab 177 составляла 25 нг / мл -1 . Показанный эксперимент представляет по крайней мере три независимых эксперимента. ( d ) Fab-фрагменты Gp41 из H почти E , проявленные к ВИЧ-1, но P устойчиво IgG S eronegative (HEPS) не являются полиспецифическими. Полиспецифичность оценивали с помощью ELISA, сравнивая связывание антител с gp41 и пептидом P1 со связыванием с нерелевантными антигенами, такими как gp120, бычий сывороточный альбумин (BSA), ДНК, пептид гемагглютинина (HA) и кардиолипин, как описано в разделе «Методы».Концентрация Fab 66 и 69 составляла 100 нг / мл -1 , а Fab 177 и 43 составляла 25 нг / мл -1 .
На левой панели связывание Fab 177 с gp41-MPR частично конкурировало с Fab 66 и заметно — с Fab 69; на правой панели связывание Fab 66 с gp41-MPR конкурировало с Fab 177. В обеих панелях 2F5, специфичный для эпитопа ELDKWA на Gp41, и неспецифический IgG D1.3, используемые в качестве контроля, не показали конкуренции.Концентрация Fabs 66 составляла 100 нг / мл -1 , а Fab 177 составляла 25 нг / мл -1 . Показанный эксперимент представляет по крайней мере три независимых эксперимента. ( d ) Fab-фрагменты Gp41 из H почти E , проявленные к ВИЧ-1, но P устойчиво IgG S eronegative (HEPS) не являются полиспецифическими. Полиспецифичность оценивали с помощью ELISA, сравнивая связывание антител с gp41 и пептидом P1 со связыванием с нерелевантными антигенами, такими как gp120, бычий сывороточный альбумин (BSA), ДНК, пептид гемагглютинина (HA) и кардиолипин, как описано в разделе «Методы».Концентрация Fab 66 и 69 составляла 100 нг / мл -1 , а Fab 177 и 43 составляла 25 нг / мл -1 . Показанный эксперимент представляет по крайней мере три независимых эксперимента.
Показанный эксперимент представляет по крайней мере три независимых эксперимента.
Слайд PowerPoint
Затем конкурентное связывание Fab 66, 69 и 177 с gp41 оценивали с помощью конкурентного ELISA. Взаимодействие Fab 177 с gp41-MPR полностью ингибировалось 20-кратным избытком Fab 69 (фиг. 4c) и наоборот (данные не показаны).Поскольку эти два Fab-фрагмента имеют идентичные тяжелые цепи, но разные легкие цепи, эти результаты предполагают, что тяжелая цепь этих Fab-фрагментов участвует в связывании с gp41, легкая цепь модулирует аффинность. Соответственно, выделенная тяжелая цепь, полученная удалением гена легкой цепи из вектора, не могла связывать gp41 (данные не показаны). Взаимодействие Fab 66 с gp41 почти полностью ингибировалось избытком Fab 177, но, наоборот, взаимодействие gp41 с Fab 177 было только частично ингибировано избытком Fab 66 (фиг. 4c).Это предполагает, что эти два Fab распознают разные, но перекрывающиеся эпитопы с разной аффинностью. Конкуренция между Fab 69 и 66, каждый из которых содержит λ-легкую цепь, не могла быть оценена в таком анализе (см. Методы).
Конкуренция между Fab 69 и 66, каждый из которых содержит λ-легкую цепь, не могла быть оценена в таком анализе (см. Методы).
Наконец, ни один из Fab-клонов, которые были бы специфичными ни для P1- или gp41-связанного gp120, ни для различных нерелевантных антигенов, таких как бычий сывороточный альбумин, ДНК, кардиолипин или пептид гемагглютинина (HA) (рис. 4d), как имеет было предложено Haynes et al. 35 для 4E10 и 2F5, но не подтверждено другими. 36, 37
Gp41-специфические Fab IgA из HEPS блокируют трансцитоз ВИЧ-1 через плотный эпителиальный барьер
Ранее мы сообщали, что gp41-специфический IgA слизистой оболочки у ВИЧ-1-серопозитивных пациентов блокировал трансцитоз ВИЧ-1 либо внутриклеточно. внутри эпителиальных клеток или до проникновения ВИЧ-1 в эпителий 11, 19 Аналогичным образом и в соответствии с результатом, показанным на рисунке 1 для ВИЧ-1 клады D, тропического лабораторного изолята X4 NDK, выделенных gp41-специфических клонов Fab от наших HEPS индивидуумов блокировал трансцитоз первичных тропических изолятов R5 из клады B и C с IC90 <50 нг / мл -1 (рис. 5).Блокирующая активность IgA Fabs для тропических вирусов R5 (IC90 = 150 нг / мл -1 для различных клонов) намного выше, чем у IgG 2F5 (IC90 = 1 мкг / мл -1 ). Интересно, что Fab IgA были немного более эффективными при блокировании тропического R5, чем при блокировании лабораторно адаптированного, в остальном очень заразного вируса клады D NDK, X4. Перекрестная нейтрализация между тропическими изолятами X4 и клады B и C R5 подтверждает, что эпитопы, распознаваемые производными HEPS Fab IgA, консервативны либо на уровне последовательности, либо на конформационном уровне.
5).Блокирующая активность IgA Fabs для тропических вирусов R5 (IC90 = 150 нг / мл -1 для различных клонов) намного выше, чем у IgG 2F5 (IC90 = 1 мкг / мл -1 ). Интересно, что Fab IgA были немного более эффективными при блокировании тропического R5, чем при блокировании лабораторно адаптированного, в остальном очень заразного вируса клады D NDK, X4. Перекрестная нейтрализация между тропическими изолятами X4 и клады B и C R5 подтверждает, что эпитопы, распознаваемые производными HEPS Fab IgA, консервативны либо на уровне последовательности, либо на конформационном уровне.
gp41-специфических Fab из H очень E , проявленных к ВИЧ-1, но P устойчиво IgG S eronegative (HEPS) блокируют трансцитоз вируса иммунодефицита человека типа 1 (ВИЧ-1). Трансцитоз ВИЧ-1, индуцированный ВИЧ-1-инфицированными клетками, через плотный монослой эпителиальных клеток контролировали, как описано в разделе «Методы». CEM, хронически инфицированные X4 тропической кладой D HIV-1 (светло-серые столбцы), PBMC, инфицированные R5 тропической кладой B (темно-серые столбцы), или PBMC, инфицированные изолятами клады C (черные столбцы), использовали для инициации трансцитоза.Fabs (50–150 нг / мл -1 ) или контрольный 2F5 IgG (1 мкг мл -1 ) или неспецифический IgA (150 нг / мл -1 ) предварительно инкубировали с ВИЧ-1-инфицированными клетками перед инокуляцией. ВИЧ-1-инфицированных клеток на апикальном полюсе эпителиального монослоя. Трансцитоз оценивали через 2 часа, измеряя содержание p24 в базальной среде. Результаты представлены в виде процента трансцитоза в присутствии неспецифического IgA (крайний справа). Планки погрешностей представляют собой среднее значение по крайней мере трех независимых экспериментов.
CEM, хронически инфицированные X4 тропической кладой D HIV-1 (светло-серые столбцы), PBMC, инфицированные R5 тропической кладой B (темно-серые столбцы), или PBMC, инфицированные изолятами клады C (черные столбцы), использовали для инициации трансцитоза.Fabs (50–150 нг / мл -1 ) или контрольный 2F5 IgG (1 мкг мл -1 ) или неспецифический IgA (150 нг / мл -1 ) предварительно инкубировали с ВИЧ-1-инфицированными клетками перед инокуляцией. ВИЧ-1-инфицированных клеток на апикальном полюсе эпителиального монослоя. Трансцитоз оценивали через 2 часа, измеряя содержание p24 в базальной среде. Результаты представлены в виде процента трансцитоза в присутствии неспецифического IgA (крайний справа). Планки погрешностей представляют собой среднее значение по крайней мере трех независимых экспериментов.
Слайд PowerPoint
Антитела, выделенные из библиотеки HEPS IgA, нейтрализуют
in vitro инфекцию мононуклеарных клеток Следующим важным событием передачи ВИЧ-1 на участках слизистой оболочки является инфицирование дендритных клеток слизистой оболочки, CD4 + Т-лимфоцитов или макрофагов, которые инициирует распространение инфекции. Поэтому мы оценили с помощью различных анализов, проведенных независимо в трех разных лабораториях, способность Fab нейтрализовать ВИЧ-1-инфекцию различных клеток-мишеней.Fabs 66 и 69, специфичные для тримерного gp41-ΔMPR, и Fab 43, специфичные для P1-нейтрализованной инфекции макрофагов R5 тропиком, кладой B HIV-1 (BaL) с IC90-30-45 нг / мл -1 , аналогично IgG 2F5 ( Таблица 1). Fab 177 проявлял гораздо более сильную нейтрализующую активность с более низким значением IC90 100 (0,3 нг мл -1 ) (таблица 1). Напротив, преобразование Fab 177 из изотипа IgA в IgG (путем переключения Ch2 alpha на Ch2 gamma) резко снижает нейтрализующую активность Fab 177. Таким образом, в форме IgG Fab 177 показал IC90 = 25 нг / мл -1 , аналогичный таковому для Fab других библиотек и IgG 2F5.IgA Fab 177 также обладал высокой степенью нейтрализации в очень чувствительном анализе, основанном на псевдовирусной инфекции клеток TZM-bl (таблица 1). 38 Кроме того, IgA Fab 177 нейтрализовал инфекцию дендритных клеток тропическими изолятами JR-CSF ВИЧ-1 R5 с IC50, аналогичным IC50 2F5IgG (данные не показаны).
Поэтому мы оценили с помощью различных анализов, проведенных независимо в трех разных лабораториях, способность Fab нейтрализовать ВИЧ-1-инфекцию различных клеток-мишеней.Fabs 66 и 69, специфичные для тримерного gp41-ΔMPR, и Fab 43, специфичные для P1-нейтрализованной инфекции макрофагов R5 тропиком, кладой B HIV-1 (BaL) с IC90-30-45 нг / мл -1 , аналогично IgG 2F5 ( Таблица 1). Fab 177 проявлял гораздо более сильную нейтрализующую активность с более низким значением IC90 100 (0,3 нг мл -1 ) (таблица 1). Напротив, преобразование Fab 177 из изотипа IgA в IgG (путем переключения Ch2 alpha на Ch2 gamma) резко снижает нейтрализующую активность Fab 177. Таким образом, в форме IgG Fab 177 показал IC90 = 25 нг / мл -1 , аналогичный таковому для Fab других библиотек и IgG 2F5.IgA Fab 177 также обладал высокой степенью нейтрализации в очень чувствительном анализе, основанном на псевдовирусной инфекции клеток TZM-bl (таблица 1). 38 Кроме того, IgA Fab 177 нейтрализовал инфекцию дендритных клеток тропическими изолятами JR-CSF ВИЧ-1 R5 с IC50, аналогичным IC50 2F5IgG (данные не показаны).
Далее мы сконцентрировались на Fab 177 из-за его более высокой нейтрализующей активности по сравнению с другими gp41-ΔMPR-специфическими клонами.Используя в качестве мишеней как человеческие CD8 + мононуклеарные клетки периферической крови с истощенными Т-клетками, так и Т-клетки CD4 + , Fab 177 нейтрализовал ВИЧ-1 инфекцию двумя клонами тропической клады B ВИЧ-1 R5 — либо JR- CSF или полевой изолят 93BR029, R5 тропический вирус клады B — в зависимости от концентрации (таблица 2). В качестве контроля предварительная инкубация вируса с зидовудином (AZT) полностью подавляла инфекцию до уровня, сопоставимого с ложно инфицированными клетками (дополнительная фигура S2). В этих анализах нейтрализации Fab 177 снова оказался намного более эффективным в нейтрализации ВИЧ-1, чем IgG 2F5 (таблица 2).При использовании PBMC, обедненных CD8 + Т-клетками, IC50 составлял 6 нМ для Fab 177 по сравнению с 60 нМ для IgG 2F5, а IC70 составлял 60 нМ для Fab 177 по сравнению с 200 нМ для IgG 2F5. Различие между Fab 177 и IgG 2F5 было еще более значительным в отношении инфицирования CD4 + Т-клеток Fab 177, демонстрируя более чем в 10 раз более высокую эффективность, чем IgG 2F5 (IC90 = 60 нМ).
Различие между Fab 177 и IgG 2F5 было еще более значительным в отношении инфицирования CD4 + Т-клеток Fab 177, демонстрируя более чем в 10 раз более высокую эффективность, чем IgG 2F5 (IC90 = 60 нМ).
Молекулярная характеристика P1-специфических Fab 43
Хотя подавляющее большинство анализов генов антител к ВИЧ-1 было выполнено на IgG, об IgA известно мало.Можно ожидать различий между IgA и IgG слизистой оболочки, поскольку репертуар IgA слизистой оболочки сильно разделен. 39
Разнообразие в репертуаре первичных В-клеток создается комбинаторной перегруппировкой различных переменных (V), разнообразия (D), соединения (J), генных сегментов, разнообразием соединений и ассоциацией различных тяжелых и легкие цепи. 40, 41 Дополнительное разнообразие может возникнуть во время развития иммунного ответа путем соматической гипермутации.
Связывание и специфичность антитела опосредуются вариабельными областями тяжелой (H) и легкой (L) цепей. 42 Каждая вариабельная область содержит три гипервариабельных субрегиона, называемых определяющими комплементарность областями CDR1, CDR2 и CDR3, разделенными четырьмя каркасными областями (FR1, FR2, FR3 и FR4). Каждая H-область кодируется тремя сегментами зародышевой линии: вариабельной (V H ), разнообразной (D H ) и соединяющейся (J H ). Хотя CDR1 и CDR2 кодируются V H , CDR3 кодируется сегментом D H и началом сегмента J H .Следовательно, CDRh4 представляет собой петлю, наиболее разнообразную по составу и длине (предполагаемое потенциальное разнообразие составляет 10 23 последовательностей). Гены 43 V H делятся на семь семейств (V H 1 – V H 7), каждое из которых отличается общей нуклеотидной гомологией> 80% в консервативных областях. 42 Самое большое семейство генов, V H 3, включает примерно половину экспрессируемого репертуара V H в периферических В-клетках взрослых.
42 Самое большое семейство генов, V H 3, включает примерно половину экспрессируемого репертуара V H в периферических В-клетках взрослых.
Легкая цепь образуется путем реаранжировки вариабельного гена зародышевой линии (V L ) с присоединяющимся геном (J L ).CDR 1 и 2 кодируются сегментом V L и CDR3 3′-концом V L и 5′-концом сегментов J L . У человека есть два типа легких цепей: κ (60%) и λ (40%). Разнообразие репертуара обусловлено не только перестановкой сегментов V H , D H и J H и сегментов V L и J L , но и различными возможными комбинациями между тяжелыми и легкими. цепями и соматическими мутациями, которые группируются в областях CDR.
Анализ сегмента VH: Fab 43 показал длинный CDRh4 из 22 аминокислот, характеристики нейтрализующих оболочку антител ВИЧ-1 Характеристики области Ch2 указывают на то, что Fab 43 имеет изотип IgA1. Вариабельная область Ig H Fab 43 (рис. 2a) состоит из гена VH из самого большого семейства генов, Vh4, в сочетании с аллелем D3-3 * 01 семейства DH, за которым следует сегмент гена JH6. Гомология нуклеотидов на 91% была обнаружена с геном зародышевой линии Vh4-30 * 01, различающимся на 34 нуклеотида и 17 аминокислотных остатков (9 в CDR и 8 в FR областях) (таблица 3a).Области CDR имели самый высокий уровень мутаций замещения нуклеотидов с соотношением замен (R) к молчащим (S) нуклеотидным заменам (R: S) 2,75. Мутации в CDRh2 и CDRh3 приводят к аминокислотным изменениям на 25 и 50% по сравнению с геном зародышевой линии Vh4-30-3 * 01 (Таблица 4).
Вариабельная область Ig H Fab 43 (рис. 2a) состоит из гена VH из самого большого семейства генов, Vh4, в сочетании с аллелем D3-3 * 01 семейства DH, за которым следует сегмент гена JH6. Гомология нуклеотидов на 91% была обнаружена с геном зародышевой линии Vh4-30 * 01, различающимся на 34 нуклеотида и 17 аминокислотных остатков (9 в CDR и 8 в FR областях) (таблица 3a).Области CDR имели самый высокий уровень мутаций замещения нуклеотидов с соотношением замен (R) к молчащим (S) нуклеотидным заменам (R: S) 2,75. Мутации в CDRh2 и CDRh3 приводят к аминокислотным изменениям на 25 и 50% по сравнению с геном зародышевой линии Vh4-30-3 * 01 (Таблица 4).
Поразительно, петля CDR3 тяжелой цепи (CDRh4) очень длинный и содержит 22 аминокислоты. Такой расширенный CDRh4 был описан как одна из молекулярных характеристик редких ВИЧ-1, широко нейтрализующих IgG, либо специфичных для gp120, а именно IgG 2G12 и b12, либо специфичных для gp41-MPR, а именно IgG 2F5 и 4E10. 44, 45, 46
Такой расширенный CDRh4 был описан как одна из молекулярных характеристик редких ВИЧ-1, широко нейтрализующих IgG, либо специфичных для gp120, а именно IgG 2G12 и b12, либо специфичных для gp41-MPR, а именно IgG 2F5 и 4E10. 44, 45, 46
Области D (H) Fab 43 сравнивали с локусом иммуноглобулина D (H) и показали гомологию с сегментом D3-3, с 21 нуклеотидом, гомологичным из 31 (68% гомологии) и с 8 дополнительными нуклеотидами в 5′-конец и 10 дополнительных на 3′-конце последовательности зародышевой линии.Эти изменения приводят к удлинению CDR3 из 22 аминокислотных остатков (дополнительная таблица SII).
Анализ сегмента VL Легкая цепь Fab 43 принадлежит к λ-типу. Его нуклеотидная последовательность показала 92% гомологии с геном зародышевой линии VL1-47 * 01, отличаясь 20 нуклеотидами и 17 аминокислотными остатками (4 в CDR и 13 в FR областях) (таблица 3b). Неожиданно процент аминокислотных изменений был высоким как в FR (особенно в FR2), так и в CDR областях (Таблица 4).Соматические мутации в областях FR и CDR в Fab 43 (оцениваемые по отношениям R: S) были больше в FR, чем в областях CDR (таблица 3b). Не удалось найти выравнивания для сегмента J.
Молекулярная характеристика тримерных gp41-специфичных клонов Fab
Что касается Fab 43, все эти gp-41-ΔMPR-специфические Fab были изотипами IgA1. Помимо идентичности последовательностей вариабельных областей тяжелой и легкой цепей между двумя клонами (59 и 69), идентичные тяжелые цепи, но разные легкие цепи, обнаруженные в шести других клонах (69, 97, 105, 125, 154 и 177), указывают на значительная неразборчивость тяжелых цепей, наблюдаемая in vivo . 47 Такая неразборчивость также не редкость для Fab, выделенных из библиотек фагового дисплея. 46
Анализ выраженного V H гены Вариабельная область IgH Fabs 69 и 177 включала ген VH из семейства VH6 в сочетании с аллелем D2-8 семейства DH, за которым следует сегмент гена Jh5. Напротив, вариабельная область IgH Fab 66 состоит из гена VH из семейства Vh2 в ассоциации с аллелем D3-9, за которым следует ген JH5 (таблица 3a).
Напротив, вариабельная область IgH Fab 66 состоит из гена VH из семейства Vh2 в ассоциации с аллелем D3-9, за которым следует ген JH5 (таблица 3a).
Анализ области VH показал, что Fab 69 и 177 были тесно связаны с геном зародышевой линии VH6-1 * 01 (94% гомологии). Fab 66 показал большее разнообразие от зародышевой линии, поскольку он был только на 89% гомологичен ближайшему гену зародышевой линии Vh2-69 * 01 (таблица 3a).
Анализ соматических мутаций в областях FR и CDR 69, 177 и 66 gp41-реактивных клонов Fab показал, что отношения R: S были самыми высокими в CDR по сравнению с областью FR, как и ожидалось для антител с созревшей аффинностью. только для клона 66 (таблица 3а).
Ген Fab 66, реаранжированный VH, отличался 25 нуклеотидами и 14 аминокислотными остатками (7 в FR и 7 в CDR) от ближайшей зародышевой линии Vh2-69 * 01 (таблица 3a). Мутации в CDR1 и CDR2 Fab 66 приводят к обмену на 16,66 и 60% соответственно по сравнению с геном зародышевой линии Vh2-69 * 01, тогда как мутации в FR2 и FR3 приводят к изменениям аминокислот на 11,76 и 12,83% соответственно. (Таблица 4). Гены, реаранжированные Fab 69 VH и Fab 177 VH, отличались 14 нуклеотидами и 12 аминокислотными остатками (8 в FR и 4 в CDR) от ближайшей зародышевой линии VH6-1 * 01 (таблица 3a).Значение R: S было выше для FR, чем для CDR-областей (Таблица 3a) из-за высокого уровня заменяющих мутаций в FR3 (15,38%). Эти данные предполагают, что эта петля CDR участвует в связывании антигена, как было показано для других антител против ВИЧ-1 (обзор в Viau and Zouali 42 ). Мутации в CDR1 и CDR2 приводят к изменениям аминокислот на 8,33 и 30% соответственно по сравнению с геном зародышевой линии VH6-1 * 01 (таблица 4). Удивительно, но все нуклеотидные мутации, идентифицированные в CDR2 этих клонов, приводят к аминокислотным изменениям (что объясняет значение ∞ отношения R: S) (Таблица 3a).
(Таблица 4). Гены, реаранжированные Fab 69 VH и Fab 177 VH, отличались 14 нуклеотидами и 12 аминокислотными остатками (8 в FR и 4 в CDR) от ближайшей зародышевой линии VH6-1 * 01 (таблица 3a).Значение R: S было выше для FR, чем для CDR-областей (Таблица 3a) из-за высокого уровня заменяющих мутаций в FR3 (15,38%). Эти данные предполагают, что эта петля CDR участвует в связывании антигена, как было показано для других антител против ВИЧ-1 (обзор в Viau and Zouali 42 ). Мутации в CDR1 и CDR2 приводят к изменениям аминокислот на 8,33 и 30% соответственно по сравнению с геном зародышевой линии VH6-1 * 01 (таблица 4). Удивительно, но все нуклеотидные мутации, идентифицированные в CDR2 этих клонов, приводят к аминокислотным изменениям (что объясняет значение ∞ отношения R: S) (Таблица 3a).
Области D (H) тримерных gp41-специфичных Fab сравнивали с локусом иммуноглобулина D (H). В дополнительной таблице SII показано распределение отдельных сегментов D (H) Fabs 66, 69 (и 177) с локусом D (H). Fab 66 показал гомологию с сегментом D3-9 с 15 гомологичными нуклеотидами из 32 (47% гомологии). Он также имел 6 дополнительных нуклеотидов и 1 дополнительную вставку P на 5′-конце и 10 дополнительных нуклеотидов на 3′-конце зародышевой линии, что приводило к удлинению CDRh4 из 15 аминокислотных остатков.Fab 69 и 177 показали гомологию с сегментом D2-8, с 14 из 31 гомологичных нуклеотидов (45% гомологии), что привело к CDRh4 длиной 11 аминокислотных остатков.
Fab 69 и 66 были λ-типа и происходили из семейства генов VL1 зародышевой линии. Напротив, Fab 177 имел κ-легкую цепь семейства генов VK1.Сегмент J принадлежал к семейству J3 для Fabs 69 и 66 и к J1 для Fab 177 (Таблица 3b). Сравнение нуклеотидных последовательностей этих легких цепей Fab с последовательностью гена зародышевой линии VL1-47 * 01, как уже было показано для тяжелой цепи, показало высокую нуклеотидную гомологию для Fab 69 (96%), но более низкую гомологию нуклеотидов (87% ) для Fab 66 (таблица 3b). Анализ соматических мутаций показал, что ген Fab 69 VL отличается восемью нуклеотидами и шестью аминокислотными остатками (четырьмя в FR и двумя в CDR) от зародышевой линии VL1-47 * 01, тогда как ген Fab 66 VL отличается на 36. нуклеотидов, но только 17 аминокислот (8 в FR и 9 в CDR) из зародышевой линии VL1-47 * 01 (Таблица 3b).Была только 1 молчащая мутация, но 11 заменяющих мутаций в петлях CDR, которые объясняют соотношение R: S, равное 11 для клона 66 (таблица 3b). Неожиданно мутации в CDR1 и CDR2 Fab 66 вызывали более значительные аминокислотные изменения в CDR1 (41,66%) по сравнению с CDR2 (30%) (таблица 4), что позволяет предположить участие CDR1 в связывании антигена. Для Fab 69 CDR содержат только две заменяющие мутации и никакой молчащей мутации (значение отношения R: S равно ∞) (таблица 3b).
Клон 177 имеет 100% нуклеотидную гомологию с геном зародышевой линии VK1D39 * 01.
Антигенный атлас ускользания ВИЧ-1 от широко нейтрализующих антител
Введение
За последнее десятилетие все большее количество широко нейтрализующих антител (bnAbs) было выделено от ВИЧ-инфицированных людей. Эти антитела нацелены на консервативные области Env, которые являются многообещающими мишенями для вакцины (Kwong and Mascola, 2018). Кроме того, их широкая нейтрализующая активность и способность направлять уничтожение инфицированных клеток делают bnAbs перспективными противовирусными иммунотерапевтическими препаратами для профилактики, терапии и лечения ВИЧ (Margolis et al., 2017; Пегу и др., 2017).
Однако bnAbs сталкивается с грозным противником. Исключительная эволюционная способность HIV Env позволяет вирусу оставаться на шаг впереди bnAbs во время инфекции, и устойчивость часто возникает, когда bnAb терапевтически вводят инфицированным животным (Klein et al., 2012; Poignard et al., 1999; Shingai et al., 2013) или людей (Bar et al., 2016; Caskey et al., 2015, 2017; Lynch et al., 2015a; Scheid et al., 2016; Trkola et al., 2005). Таким образом, определение мутаций, которые опосредуют ускользание вируса, важно для оптимизации и оценки иммунотерапии bnAb и вакцин.
Несмотря на то, что обширные усилия были направлены на структурную характеристику эпитопов bnAb с помощью рентгеновской кристаллографии и крио-электронной микроскопии (крио-ЭМ), самих структур недостаточно для полного определения функционального эпитопа (Cunningham and Wells, 1993; Kelley). и O’Connell, 1993), определяемые здесь как сайты, в которых мутации влияют на нейтрализацию антителами репликационно-компетентного вируса. Внесение отдельных мутаций в Env и выполнение анализов нейтрализации может предоставить информацию о функциональном эффекте конкретных мутаций, но даже самые крупные исследования, в которых используется однократный мутагенез, могут анализировать только небольшую часть всех возможных мутаций Env.
Мы недавно описали мутационное антигенное профилирование, массовый параллельный экспериментальный подход для количественной оценки того, как все мутации одной аминокислоты в Env влияют на нейтрализацию антител (Dingens et al., 2017). Этот подход включает создание библиотек ВИЧ, которые несут все мутации аминокислот Env, совместимые с вирусной репликацией, инкубацию этих библиотек с антителом или без него, инфицирование Т-клеток и использование глубокого секвенирования для количественной оценки обогащения каждой мутации в выбранных по сравнению с не- избранные библиотеки.Здесь мы применяем этот подход к панели из девяти bnAbs, нацеленных на пять эпитопов Env, а также к пулу из двух bnAbs. Полученные в результате карты ускользания вируса обеспечивают всестороннее представление на уровне мутаций функциональных интерфейсов между ВИЧ и bnAbs.
Результаты
Полные карты ускользания вируса из панели bnAbs
Чтобы получить полную картину ускользания вируса, мы выбрали девять bnAbs, нацеленных на пять наиболее охарактеризованных эпитопов на Env (рис. 1A). В частности, панель bnAb включает сайт связывания CD4 (CD4bs) bnAbs VRC01 (Wu et al., 2010) и 3BNC117 (Scheid et al., 2011), гликановый суперсайт V3 / N332 bnAbs PGT121 (Walker et al., 2011) и 10-1074 (Mouquet et al., 2012), вершина гликана / тримера V2 bnAbs PG9 (Walker et al., 2009) и PGT145 (Walker et al., 2011), гибридный пептид и интерфейс gp120 / gp41 bnAbs PGT151 (Falkowska et al., 2014) и N123-VRC34.01 (впоследствии обозначаемый как VRC34 .01, Kong et al., 2016) и проксимальной внешней области мембраны (MPER) bnAb 10E8 (Huang et al., 2012). Следы связывания этих антител были ранее охарактеризованы с использованием структурных методов (рис. 1B), что позволяет нам сравнивать структурных эпитопов с функциональными эпитопами , определенными в этом исследовании.
Рисунок 1: Схема мутационного антигенного профилирования панели bnAbs.A. Панель bnAb. Меры ширины взяты из 100 наиболее часто используемых вирусов в CATNAP LANL (Yoon et al., 2015). B. Для каждого эпитопа структурно определенные сайты контакта с антителом обозначены цветами сбоку и сверху на тример BG505 SOSIP Env (PDB: 5FYL). C. Рабочий процесс эксперимента по мутационному антигенному профилированию и пример данных из bnAb 10E8. Библиотеку мутантного вируса инкубируют с антителом или без него перед заражением SupT1.Клетки CCR5. Неинтегрированная вирусная кДНК из инфицированных клеток глубоко секвенируется для количественной оценки частоты каждой мутации env как в условиях селекции антител, так и в условиях ложной селекции, а общая фракция вирусной библиотеки, которая выживает при нейтрализации антител, количественно определяется с помощью кПЦР. Доля каждого мутанта, который выживает после нейтрализации, отображается на уровне сайта на линейных графиках и на уровне мутации на логических графиках. Высота каждой буквы пропорциональна доле вирионов с этой аминокислотой, которые пережили отбор антител, превышающую общее среднее значение библиотеки.
Мы картировали ускользание от этих антител, используя BG505.T332N Env, который происходит от переданного штамма ВИЧ-основателя подтипа A (Wu et al., 2006). Этот тример Env широко используется в структурных исследованиях и исследованиях вакцинации (Sanders et al., 2013; Ward and Wilson, 2017). Мы использовали вирусные библиотеки, которые были ранее созданы путем внесения всех возможных аминокислотных мутаций в эктодомен и трансмембранный домен Env (Haddox et al., 2018). Существует 19 аминокислотных мутаций × 670 сайтов = 12730 таких аминокислотных мутаций, и наши библиотеки содержат подмножество этих мутаций, совместимых с ростом вируса в культуре клеток.
Чтобы количественно оценить, как каждая из этих мутаций влияет на чувствительность антител к ВИЧ, мы нейтрализовали независимо созданные библиотеки мутантных вирусов при концентрации антител ∼IC 95 -IC 99,9 и глубоко секвенировали гены env вирусов, которые были способны к инфицировать клетки в присутствии антител (рис. 1C). Для каждого антитела мы выполнили не менее двух повторов с использованием независимо созданных вирусных библиотек (рисунок S1). Мы провели параллельные контрольные эксперименты без антител и сравнили относительную частоту каждой мутации Env в выбранной антителом библиотеке с невыбранным контролем.Масштабируя эту относительную частоту к общей доле всей библиотеки, которая выжила при отборе антител, мы оценили долю вирионов с каждой мутацией, которые выживают при отборе, далее называемую фракцией , выживающей (Doud et al., 2018). Чтобы выделить ускользающие мутации, мы построили график избыточной доли выживших выше общего среднего значения библиотеки.
Каждое антитело воспроизводимо отбирало мутации только в небольшом подмножестве сайтов Env (рисунок 2A, рисунок S1).Полные карты уровня мутаций ускользания вируса от каждого антитела нанесены на всю мутагенизированную часть env в файле S1. Антитела, нацеленные на один и тот же эпитоп, имеют тенденцию к отбору мутаций в аналогичных областях Env, и эти мутации группируются в трехмерную структуру в области связывания антитела или рядом с ней (рис. 2А). Это наглядно демонстрируют PG9 и PG145, где выбранные позиции в значительной степени перекрываются. Также обратите внимание на то, что величина эффекта мутаций варьировалась в зависимости от антител (сравните оси y на рисунке 2A), как и кажущийся шум на графиках; последствия этого более подробно обсуждаются в заключительном разделе результатов.
Рис. 2: Профили ускользания по всему Env в зависимости от положений аминокислот, которые контактируют с bnAb.A. Линейные графики показывают нейтрализацию выживших антител избыточной фракции, усредненной по всем мутациям в каждом сайте. Структурно определенные контактные площадки обозначены синей линией. Структуры показывают тример BG505 Env SOSIP (PDB: 5FYL) с сайтами значительного ускользания, окрашенными в желтый цвет, сайты контактов, окрашенными в синий цвет, и перекрытие между этими наборами сайтов, окрашенных в зеленый цвет. Для 10E8 также показана структура пептида MPER (который отсутствует в тримере SOSIP) (PDB: 4G6F). B. Столбцы указывают количество структурно определенных участков контакта для каждого антитела, при этом зеленый цвет указывает участки контакта, которые также являются участками значительного ускользания. C. Столбцы показывают количество участков значительного ускользания для каждого антитела, зеленый цвет указывает участки ускользания, которые также являются сайтами контакта. Обратите внимание, что зеленые столбцы охватывают одни и те же наборы сайтов на панелях B и C.
Чтобы тщательно сравнить перекрытие между структурными контактами и сайтами ускользания вируса, мы идентифицировали сайты значительного функционального ускользания из данных мутационного антигенного профилирования и сайты физический контакт между bnAb и Env из опубликованных структур.Мы определили значимые участки ускользания вируса, сопоставив гамма-распределение с измеренными антигенными эффектами мутаций на каждом участке, и определили участки, в которых антигенные эффекты были больше, чем ожидалось из этого распределения, с коэффициентом ложного обнаружения 0,01 (рис. S2). Мы определили сайты структурного контакта как остатки Env, которые находились в пределах 4 Å от антитела в доступных структурных моделях, учитывая только неводородные атомы (подробности см. В разделе «Методы»).
Для большинства антител только небольшая часть структурно определенных участков контакта также была участками значительного ускользания вируса (рис. 2В, рис. S3A).Кроме того, мы идентифицировали многочисленные сайты ускользания за пределы структурно определенного эпитопа для большинства антител. Степень, в которой ускользание произошло в сайтах, которые напрямую контактируют с антителом, значительно различается для bnAbs, от всех значимых сайтов ускользания для PG9 до только одного из пяти сайтов ускользания для VRC01 (рисунок 2C, рисунок S3A). Сайты ускользания, которые не контактируют напрямую с антителом, обычно находятся рядом со структурно определенным эпитопом в диапазоне 5-10 Å (рисунок S3A). Однако несколько сайтов ускользания более далеки от структурно определенного эпитопа (Рисунок 2A, Рисунок S3B).
В то время как наши карты побега включают большинство мутаций, ранее идентифицированных с использованием отдельных точечных мутантных псевдовирусов BG505 в анализах нейтрализации TZM-bl (Kong et al., 2016; Lee et al., 2017), мы также обнаружили многие ранее не охарактеризованные места побега. Мы создали и протестировали точечные мутантные псевдовирусы BG505 в анализах нейтрализации TZM-bl для трех антител, тестируя от 16 до 19 точечных мутантов для каждого антитела. Измерения мутационного антигенного профиля хорошо коррелировали с кратным изменением IC 50 из анализов нейтрализации TZM-bl для всех протестированных антител (PGT121: R = 0.76, n = 16; 10-1074: R = 0,81, n = 16; VRC01: R = 0,69, n = 19) (рисунок S4). В следующих нескольких подразделах мы сосредоточимся на каждом эпитопе Env индивидуально.
Побег из суперсайта гликана V3 bnAbs
Два антитела против V3 PGT121 и 10-1074 представляют собой клональные варианты, которые возникли у одного и того же инфицированного человека (Mouquet et al., 2012). Тем не менее, между двумя антителами были любопытные различия в специфических мутациях, которые опосредовали ускользание в наших экспериментах, а также в общих размерах эффекта мутаций ускользания (рис. 3A, 3B, обратите внимание на разные шкалы оси Y на двух панелях).Например, мутации в сайте 325 имели больший эффект на 10-1074, чем на PGT121, тогда как мутации в сайте 327 имели аналогичные эффекты. Мы подтвердили дифференциальные эффекты мутаций в сайте 325 путем тестирования трех разных мутантов в этом сайте в анализах нейтрализации TZM-bl: максимальный эффект для 10-1074 был 27-кратным увеличением IC 50 , в то время как максимальный эффект для PGT121 увеличился всего в 1,7 раза (D325E; рис. S4). Наш мутационный антигенный профиль также показал, что мутации, устраняющие гликан N332, имели больший эффект для 10-1074, чем PGT121 (рис. 3A, B), что согласуется с предыдущим исследованием, в котором изучалось связывание с gp120 (Mouquet et al., 2012). Напротив, в большинстве других сайтов эпитопа (таких как 323, 327 и 330) общие эффекты мутаций были одинаковыми между двумя антителами (рис. 3A, B).
Рис. 3. Выход из гликанового суперсита V3 и вершины bnAbs V2.A , B. Профили аварийного выхода для гликанового суперсайта V3 bnAbs PGT121 и 10-1074. Высота букв указывает на выжившую избыточную фракцию для каждой мутации. Синие кружки указывают на структурно определенные сайты контакта, а желтые подчеркивания указывают на мотив N-связанного гликозилирования.Логоплоты, показывающие переход через Env, находятся в файле S1. C. Гликановые суперсайты антитела V3 показаны синим цветом, а Env окрашен в соответствии с максимальной избыточной фракцией, выжившей на каждом участке. Обратите внимание, что для PGT121 структура близкородственного клонального варианта PGT122 используется вместо структуры PGT121 (PDB: 5FYL и 5T3Z соответственно). D , E. Профили ускользания гликанов V2 / верхушечных антител PG9 и PGT145, представленных таким же образом, как A, B. F. V2 гликановые / верхушечные антитела показаны синим, а Env окрашен в соответствии с максимальная избыточная фракция, выжившая на каждом сайте (PDB: 5VJ6 и 5V8L соответственно).
Существуют также различия в , когда мутации в данном сайте ускользают от каждого антитела. Например, хотя общий эффект всех мутаций в сайте 330 одинаков для 10-1074 и PGT121, h430R ускользает от 10-1074, но не PGT121 (фиг. 3A, B, подтверждено анализами нейтрализации TZM-bl на фиг. S4). Даже среди мутаций с небольшим эффектом анализы нейтрализации TZM-bl подтвердили результаты мутационного антигенного профилирования. Например, мутации в сайте 325 имели разные эффекты: D325E оказал большое влияние на 10-1074, но незначительный эффект на PGT121, D325S имел небольшой эффект на 10-1074, но не повлиял на PGT121, и D325N не оказал никакого влияния на любое из антител ( Рис. 3A, B и подтверждено на рис. S4).
Многие аспекты наших карт бегства согласуются с предшествующими знаниями об эпитопах этих двух антител (Garces et al., 2014; Mouquet et al., 2012; Sok et al., 2014, 2016). Например, устранение целевого гликана N332 посредством мутаций в N332 и S334 привело к ускользанию от обоих антител, как и специфичные для антител мутации в мотиве 324 GDIR 327 (рисунок 3), консервативной области этого эпитопа, которая является участвует в связывании корецептора CCR5 (Sok et al., 2016).
Однако мы также идентифицировали ускользание в сайтах, которые ранее не рассматривались как часть функционального эпитопа. Например, ускользание вируса от обоих антител произошло через мутации в сайте 415 в V4 и в умеренной, но воспроизводимой степени в сайте 441 (рис. 3A, 3B). Мы подтвердили, что мутации в каждом из этих сайтов привели к ускользанию от обоих антител в анализах нейтрализации TZM-bl (рисунок S4). Ни один из этих сайтов напрямую не взаимодействует ни с одним из антител; сайт 415 находится рядом с другими структурными контактами, а также с гликаном N332, а сайт 441 в цепи β22 соседствует с гликаном N301.
Escape from V2 apex bnAbs
Для антител V2 apex PG9 и PGT145 ускользание произошло за счет устранения гликана N160 в центре эпитопа (рис. 2A, 3D, 3E). Кроме того, ускользание происходило в структурно определенных сайтах контакта на вершине тримера для PG9 и на вершине тримера и на границе раздела для PGT145. Предыдущие исследования обоих антител предполагают, что связывание обусловлено электростатическими взаимодействиями с положительно заряженными остатками Env (Lee et al., 2017; McLellan et al., 2011; Wang et al., 2017). Мутации в этих сайтах приводили к ускользанию вируса (включая остатки R166, K169 и K171 для PG9 и K121, R161, K169 для PGT145), при этом замена зарядов часто приводит к наибольшей степени ускользания (рис. 3D, 3E). Структурные исследования показывают, что длинное плечо HCDR3 PGT145 достигает границы раздела тримеров; Предполагается, что существование этого эпитопа является результатом уравновешивающего акта «толчка» от межпротомерных зарядов отталкивания на границе раздела тримера и «притяжения» гидрофобных взаимодействий между вариабельными петлями через протомеры на вершине тримера (Lee et al., 2017).
В то время как ускользание от PGT145 происходило за счет устранения положительных зарядов эпитопа, ускользание также происходило за счет введения зарядов в сайты, где остаток дикого типа не заряжен. К ним относятся сайты 123, 124 и 127 в эпитопе или очень близко к нему, а также более отдаленные сайты, окружающие эпитоп, включая сайты 200, 202, 203 в петле β3-β4 и 312, 313 и 315 на конце. петли V3 (рис. 3F). Эти мутации предположительно также влияют на отталкивание зарядов на границе раздела тримеров и / или общую конформацию тримеров, нарушая акт электростатического баланса, который является критическим для связывания PGT145.
Ускользание от CD4bs bnAbs
Ускользание от CD4bs bnAbs VRC01 и 3BNC117 произошло как в канонически определенном эпитопе CD4bs, так и в других сайтах, удаленных от сайта связывания CD4 (Рисунок 2A, Рисунок 4). В сайте связывания CD4 мутации сайта 279 в петле D и сайта 369 в петле связывания CD4 ускользнули от обоих антител (рис. 4A). За исключением сайтов 279 и 280, специфические аминокислотные мутации в петле D, которые опосредуют ускользание, различались между VRC01 и 3BNC117.
Рисунок 4: Побег из CD4bs bnAbs.A , B. Профили выхода для CD4bs, bnAbs VRC01 и 3BNC117. Высота букв указывает на выжившую избыточную фракцию для каждой мутации. Синие кружки указывают на структурно определенные сайты контакта, а желтые подчеркивания указывают на мотив N-связанного гликозилирования. Части канонического эпитопа CD4bs подчеркнуты черным и помечены. Логоплоты, показывающие переход через Env, находятся в файле S1. C. Антитела показаны синим цветом, а Env окрашен в соответствии со средней фракцией, выжившей в каждом сайте (PDB: 5FYK и 5V8M соответственно).
Мутации с наибольшим эффектом для обоих антител вводили серин или треонин вместо аспарагина в сайте 197. Гликан N197 является частью гликанового барьера, который защищает CD4bs (Crooks et al., 2017). N197S / T как устраняет этот гликан N197, так и вводит новый потенциальный мотив N-связанного гликозилирования (PNG) в N195. Поскольку ускользание было опосредовано только S / T в сайте 197, эти данные предполагают, что устранение только N197 не приводит к ускользанию вируса, но смещение гликана N197 на N195 делает.Мы подтвердили эти наблюдения, используя точечные мутанты в анализах нейтрализации TZM-bl: простое устранение PNG N197 через N197E приводило к примерно 50-кратной эффективной нейтрализации с помощью VRC01, в то время как N197S приводило к ускользанию вируса, увеличивая IC 50 на 27- сложить относительно дикого типа (рисунок S4).
Ускользание от обоих антител также произошло через D113N, который вводит PNG в сайт 113. Сайт 113 находится в интерфейсе тримера, удаленном от CD4bs (рис. 4C), что позволяет предположить, что эта мутация может повлиять на экспозицию эпитопа CD4bs путем изменения конформации тримера или динамика.Мы подтвердили, что D113N приводит к ускользанию от VRC01, используя анализ нейтрализации TZM-bl (рис. S4). Хотя неизвестно, действительно ли PNG, созданный D113N, гликозилирован, эти данные показывают, что изменение потенциальных сайтов гликозилирования как вблизи, так и дистальнее эпитопа может влиять на нейтрализацию CD4bs bnAb.
Ускользание от 3BNC117 также происходило во многих местах рядом с тем местом, где плечо HCDR3 антитела устанавливает межпротомерные контакты, включая сайты в V3 (сайты 304, 308, 312, 316-320) и в основании петли β3-β4 ( сайты 207, 209) (Рисунок 4B).Эта четвертичная природа эпитопа 3BNC117 была впервые постулирована на основании структур ранних тримеров (Lyumkis et al., 2013), а крио-ЭМ высокого разрешения тримера BG505 в комплексе с 3BNC117 (Lee et al., 2017) подтвердила, что 3BNC117 напрямую взаимодействует с остатками 207, 308 и 316 из соседнего протомера (рис. 4B, 4C). Ранее сообщалось, что мутации в сайте 207 приводят к снижению связывания 3BNC117 (Liu et al., 2017). Мы также наблюдали ускользание вируса от 3BNC117 в сайте I326, сайте, удаленном от 3BNC117 рядом с основанием петли V3, которая принимает участие в гидрофобных взаимодействиях переменной петли, которые могут регулировать динамику тримеров (Lee et al., 2017).
Поразительно, хотя VRC01 не создает подобных структурных контактов между протомерами, как 3BNC117 (Stewart-Jones et al., 2016), мы все же наблюдали ускользание в сайтах 207, 209, 304 и 326 (Рис. 4A, 4C). Мы подтвердили, что I326Y приводит к ускользанию от VRC01 (рис. S4), но мало влияет на V3-специфические bnAbs 10-1074 и PGT121, несмотря на то, что эти антитела напрямую связываются с этим сайтом.
Ускользание от слитого пептида bnAbs
Карты выхода из PGT151 и VRC34.01 подчеркивают сложную природу конформационного слитого пептида и эпитопа интерфейса gp120 / gp41 (рис. 2A, 5A, 5B). Здесь мы повторно проанализировали данные мутационного антигенного профилирования VRC34.01 из предыдущего исследования (Dingens et al., 2018), количественно оценив эффекты мутаций с использованием показателя выживания фракции , а не показателя дифференциального отбора , использованного в более раннем исследовании, и сравнил эти данные с описанным здесь побегом BG505 Env из PGT151. В то время как оба антитела контактируют с 6 N-концевыми остатками слитого пептида (512-517), ускользание от PGT151 сосредоточено только на 3 N-концевых остатках этого пептида (512-514), а ускользание от VRC34.01 опосредуется многочисленными мутациями в сайтах 512-516 и 518. Структурные следы обоих антител сосредоточены на гибридном пептиде, но они контактируют с отдельными гликанами и белковыми областями gp120 и gp41. Опять же, их функциональные эпитопы включают отдельные подмножества этих белковых остатков и гликанов (фиг. 5A, 5B). Для обоих антител также существует множество сайтов значительного ускользания в неконтактных сайтах рядом с эпитопом (рис. 5A, 5B). Для PGT151 мы также идентифицировали сайты ускользания на более удаленных остатках в V3; механизмы побега в этих местах неясны.
Рисунок 5: Ускользание от гибридного пептида и MPER bnAbs.A , B. Профили ускользания для слитого пептида bnAbs VRC34.01 и PGT151. Высота букв указывает на выжившую избыточную фракцию для каждой мутации. Логоплоты, показывающие переход через Env, находятся в файле S1. C. Антитела к гибридному пептиду показаны синим цветом, а Env окрашен в соответствии с максимальной избыточной фракцией, выжившей в каждом сайте (PDB: 5I8H и 5FUU соответственно). D. Профиль выхода MPER bnAb 10E8, представленный так же, как A, B. E. 10E8 показан синим цветом, а пептид MPER окрашен в соответствии с максимальной избыточной фракцией, выжившей в каждом сайте (PDB 4G6F).
Ускользание от MPER bnAB
Ускользание от MPER-направленного антитела 10E8 происходило преимущественно в структурно определенных контактных сайтах, причем сайты ускользания располагались на одной стороне альфа-спирали MPER-пептида, апикальной к 10E8 (рис. 5D, 5E) (рис. 5D, 5E) ( Хуанг и др., 2012). Это точно согласуется с предыдущими исследованиями (Huang et al., 2012; Kim et al., 2014). Однако мы идентифицировали два дополнительных скромных, но значительных сайта ускользания за пределы пептида MPER, в сайтах 609 и 643 в петле C-C и в домене HR2 gp41 соответственно (рисунок 2A, рисунок S2). Мутации в этих сайтах могут изменять кинетику слияния и / или представление эпитопа MPER.
Сравнение наших мутационных карт с ускользанием in vivo во время иммунотерапии bnAb у людей
Некоторые из bnAb, которые мы охарактеризовали, использовались в исследованиях иммунотерапии человека.Некоторые из ускользающих мутаций, выявленных в нашей работе, перекрываются с мутациями, которые возникли у людей во время этих исследований (Таблица S1). Например, когда 10-1074 вводили ВИЧ-инфицированным людям, вирусные ускользающие мутации возникали в сайте 325 и PNG, который охватывает сайты 332 и 334 (Caskey et al., 2017). Это те же три сайта, где наблюдается самый сильный отбор в нашем мутационном антигенном профиле 10-1074 (рисунок 3B, таблица S1).
Существует также значительное совпадение между местами ускользания, которые мы картируем in vitro и теми, которые имели место in vivo во время лечения инфицированных людей антителами к CD4bs 3BNC117 или VRC01.Для 3BNC117 сайты перекрываются между нашими картами и испытаниями на людях (Caskey et al., 2015; Schoofs et al., 2016), включая сайты 182, 209, 279, 308, 318 и 471 (Таблица S1). Зона 279 — одно из самых сильных мест побега от 3BNC117 и VRC01 в наших экспериментах. Мутация сайта 279 была частью пути ускользания вируса у пациента, от которого был изолирован VRC01 (Lynch et al., 2015b), и возникла во время прерывания иммунотерапии VRC01 после лечения (Bar et al., 2016). Мутации в сайте 279 также сыграли роль в уходе от целевого ответа CD4bs у другого пациента (Wibmer et al., 2013) и во время иммунотерапии 3BNC117 инфицированных людей (Caskey et al., 2015; Schoofs et al., 2016).
Интересно, что наши данные также могут быть полезны для выявления ранее недооцененных мутаций ускользания во время иммунотерапии. Например, после того, как пациент V10 прошел терапию VRC01 (Bar et al., 2016), редкий вариант аминокислоты на сайте 326 был зафиксирован в вирусной популяции (Таблица S1), но потенциальная значимость этой мутации не была отмечена в оригинале. публикация, поскольку она далека от структурного эпитопа.Наш мутационный антигенный профиль показывает, что мутации в сайте 326 повышают устойчивость к VRC01 (рис. 4A), демонстрируя, как исчерпывающие карты ускользания от мутаций могут помочь в интерпретации клинических данных.
Избегание объединенных антител
Многие исследования иммунотерапии начинают лечение пациентов с помощью комбинаций bnAb. Например, одна быстро развивающаяся серия клинических испытаний включает лечение пациентов с равными концентрациями 3BNC117 и 10-1074 (NIH ClinicalTrials.Идентификаторы gov: NCT03526848, NCT02824536, NCT02825797). Поэтому мы исследовали, как ускользание от смеси этих двух антител сравнивается с ускользанием от каждого антитела в отдельности.
Мы объединили антитела в равных концентрациях, а затем отобрали наши вирусные библиотеки с пулом антител (Рисунок 6, Рисунок S5). Ускользание от объединенных антител оказалось комбинацией профилей ускользания от каждого изолированного антитела (фиг. 6A, 6B). Например, мы наблюдали побег в сайтах 325, 332 и 334, вероятно, связанный с побегом из 10-1074, а также побег в сайтах 304, 308 и 471, которые предположительно влияют на устойчивость 3BNC117.
Рисунок 6: Выход из пула 3BNC117 и 10-1074 BnAbs.A. Избыточная фракция, пережившая нейтрализацию, усреднена по всем мутациям на каждом сайте. Данные с рисунка 3 (10-1074) и рисунка 4 (3BNC117) перенесены на соответствующие участки. Для объединенных данных 3BNC117 и 10-1074 нанесено среднее значение по шести повторам. Смоделированные данные представляют собой произведение средних значений выживаемости фракции избыточных мутаций каждого антитела. B. Увеличенный логический график областей эпитопа для каждого набора данных.В A и B смоделированные данные отделены от экспериментальных данных светло-серым наложением.
Важно отметить, что ускользание от объединенных антител не происходит в местах, где мутация ускользания от одного антитела сенсибилизирует вирус к другому. Например, самыми сильными мутациями ускользания только для 3BNC117 являются N197S и N197T, которые сдвигают гликан N197 на N195 (Рисунок 4B и Рисунок 6A). Однако устранение гликана N197 увеличивает восприимчивость вируса к нейтрализации антителами, нацеленными на тот же эпитоп, что и 10-1074 (Liang et al., 2016; Таунсли и др., 2016). Мутации в сайте N197 не выбираются пулом антител в нашем мутационном антигенном профиле, по-видимому, потому, что любое преимущество в отношении ускользания от 3BNC117 сводится на нет из-за повышенной чувствительности к 10-1074. Этот пример демонстрирует возможность подавления вирусных ускользающих мутаций путем отбора антител с синергетическим действием в определенных сайтах.
Чтобы смоделировать синергетический эффект антител на подавление ускользания вируса, мы рассчитали ожидаемый профиль ускользания из пула 10-1074 и 3BNC117, просто взяв произведение величины выживаемости избыточной фракции каждой мутации для каждого антитела (рис. 6).Обоснование этого расчета состоит в том, что ожидаемая фракция вирионов с мутацией, которая должна выжить в обоих антителах, является просто продуктом фракции, которая выживет для каждого антитела в отдельности. Профиль ускользания, предсказанный этой простой моделью, близко соответствует фактическому отбору из объединенных антител (рис. 6).
В целом, эти данные предполагают, что ни одна мутация одной аминокислоты не ускользает от как 3BNC117, так и 10-1074. Скорее, низкий уровень ускользания от объединенных антител, по-видимому, представляет мутации, которые ускользают от одного антитела, но мало влияют на другое.Кроме того, сходство экспериментально измеренного профиля ускользания для объединенных антител и профиля, предсказанного по продукту индивидуальных профилей антител, предполагает, что наши карты ускользания от отдельных антител могут быть полезны для компьютерного прогнозирования возможности выхода из пулов антител.
Количественная оценка способности BG505 Env ускользать от каждого bnAb с одиночными мутациями
Анти-ВИЧ bnAb часто оценивают с точки зрения их широты и эффективности по сравнению с панелями естественных вирусных штаммов.Наши данные дают возможность рассчитать альтернативный показатель, относящийся к потенциальной эффективности иммунотерапии bnAb: способность одиночных аминокислотных мутаций повышать устойчивость к антителам конкретного вирусного штамма.
Мы использовали наш мутационный антигенный профиль, чтобы оценить легкость ускользания одной мутации Env BG505 от каждого антитела. Во-первых, мы просто качественно исследовали 100 самых больших мутаций размера эффекта для каждого антитела (рис. 7A). Для некоторых антител (например, VRC34.01), существует множество отдельных мутаций, которые эффективно избегают нейтрализации, но для других антител (таких как VRC01) только несколько мутаций влияют на ускользание, и это происходит только с эффектами умеренного размера (рис. 7A). Интересно, что для всех антител, по крайней мере, некоторые из мутаций с наибольшим размером эффекта доступны с помощью однонуклеотидных мутаций, что указывает на то, что генетический код лишь умеренно влияет на доступность мутаций ускользания (рис. 7A).
Рисунок 7: Количественная оценка способности избежать каждого bnAb.А . Избыточная фракция, выживающая для 100 мутаций с наибольшим эффектом для каждого антитела. Закрашенные кружки указывают на мутации, которые находятся на расстоянии одной нуклеотидной мутации от BG505 дикого типа, тогда как светлые кружки указывают на мутации, которые находятся на расстоянии 2 или 3 нуклеотидных мутаций от последовательности BG505 дикого типа. B. Ширина каждого антитела (количественно определенная на фиг. 2) нанесена на график в зависимости от суммы значений выживаемости избыточной фракции мутаций на всех значимых участках ускользания вируса.
Чтобы количественно оценить легкость, с которой BG505 Env может ускользать от каждого антитела посредством единичных мутаций, мы суммировали значения выживаемости фракции избыточных мутаций на каждом значительном участке ускользания вируса.Этот показатель простоты ускользания от одной мутации умеренно коррелирует с широтой охвата антитела (рис. 7B). Но особенно для самых широких антител существуют различия между широтой нейтрализации природных штаммов и легкостью ускользания одной аминокислоты с помощью BG505 (рис. 7B). Например, 10E8 имеет ширину 98%, а VRC01 имеет ширину 91%, но BG505 обладает большей способностью ускользать от 10E8 с помощью мутаций одной аминокислоты. Эти результаты подчеркивают, что, как и в случае с антителами к гриппу (Doud et al., 2018), ширина bnAb ВИЧ на природных штаммах и возможность ускользать от единичной мутации любым данным вирусным вариантом являются разными показателями, оба из которых могут быть полезны для оценки потенциал утечки вирусных антител в клинических условиях.
Обсуждение
Мы картировали, как все одиночные аминокислотные мутации в BG505 Env влияют на нейтрализацию репликационно-компетентного ВИЧ девятью прототипами bnAb. Эти карты ускользания вируса определяют функциональные эпитопы этих антител, которые, как мы показываем, отличаются от структурно определенных эпитопов. Для всех антител ускользание вируса происходило только в части структурно определенных сайтов контакта, и многие мутации ускользания происходили в сайтах за пределами прямого структурно определенного эпитопа.Это ускользание в неконтактных сайтах часто сгруппировано вблизи структурных контактов, предполагая, что изменение сети взаимодействующих сайтов рядом со структурным эпитопом может нарушить связывание или нейтрализацию антитела. В некоторых случаях ускользание также происходило в сайтах, более удаленных от эпитопа, вероятно, из-за изменений конформации или динамики Env.
Другие ранее отмечали, что не все структурно определенные контактные сайты влияют на связывание антител (Falkowska et al., 2012; Kelley and O’Connell, 1993; Li et al., 2011), а мутации вне структурного эпитопа могут влиять на чувствительность антител (Back et al., 1993; Blish et al., 2008; Boyd et al., 2015; Bradley et al., 2016; O’Rourke et al. , 2012; Sethi et al., 2013). Однако наши полные карты ускользающих мутаций позволяют систематически количественно определять перекрытие между функциональными и структурными эпитопами bnAbs. Наша работа также подчеркивает, как не полностью предшествующие структурные и вирусологические анализы определили функционально релевантные взаимодействия между ВИЧ и антителами.Несмотря на то, что наше исследование было сосредоточено на некоторых из наиболее охарактеризованных bnAb против ВИЧ, мы обнаружили многочисленные участки в Env (часто за пределами структурного эпитопа), где ранее не было известно, что мутации влияют на чувствительность bnAb.
Одно из приложений, для которого наши карты утечки вирусов могут быть незамедлительно использованы, — это оценка иммунотерапии bnAb. Во время испытаний иммунотерапии возникает множество вирусных мутаций, и важно знать, какие из них изменяют чувствительность к bnAb, используемому в исследовании (Bar et al., 2016; Caskey et al., 2015, 2017; Lynch et al., 2015a; Scheid et al., 2016; Schoofs et al., 2016). Хотя наши карты, конечно, не вполне предсказывают побег, который происходит in vivo , на который могут влиять многие стохастические факторы, включая конкретный вариант, инфицировавший человека, они определяют функциональные эпитопы, которые можно использовать для оценки потенциальной антигенной значимости. вирусных мутаций. Например, наша карта функционального эпитопа VRC01 позволила нам заново интерпретировать потенциальное значение ранее незамеченной мутации вне структурного эпитопа, которая произошла во время иммунотерапии VRC01.
Мы также изучили, как наши карты ускользания от отдельных антител сравниваются с картами, созданными с использованием пула антител — важный вопрос, поскольку комбинации bnAb являются четким будущим направлением в иммунотерапии антителами (Wagh et al., 2016, 2018). Мы составили карту побега из одного такого клинически значимого пула из двух bnAbs, 3BNC117 и 10-1074. Мы обнаружили, что не было никаких мутаций, которые надежно ускользнули бы от обоих антител. Кроме того, комбинированный отбор двух антител был подобен тому, который был предсказан по продукту индивидуальных профилей ускользания.Следовательно, антигенные карты для отдельных антител могут быть полезны для моделирования ускользания вируса от комбинаций антител.
Наши данные также позволяют количественно оценить легкость, с которой BG505 Env может ускользать от каждого антитела посредством единичных мутаций. Несмотря на то, что этот показатель легкости ускользания содержит множество предостережений, характерных для нашей экспериментальной системы (он рассматривает только отдельные мутации BG505, которые поддерживают репликацию вируса в культуре клеток), предостережения также применимы к другим методам оценки вероятности того, что вирус ускользнет от антитела. .Например, характеристика ускользания вируса во время иммунотерапии на животных моделях часто исследует только один вирусный штамм на ограниченном количестве животных. Мы обнаружили, что легкость ускользания BG505 путем единичных мутаций коррелирует с широтой антител против естественных последовательностей, но есть также различия, особенно среди самых широких антител. Мы предполагаем, что оба показателя могут быть полезны для оценки потенциальной эффективности терапии на основе bnAb.
Конечно, важно иметь в виду, что наши карты измеряют влияние только одной аминокислотной мутации на BG505.Эпистатические взаимодействия между множественными мутациями могут играть роль в приспособлении вирусов и уклонении от иммунитета (Adams et al., 2017; Dahirel et al., 2011; Lynch et al., 2015b; Otsuka et al., 2018; Da Silva et al., 2010; Troyer et al., 2009; Wu et al., 2017). Точно так же мы проводили наши эксперименты на едином генетическом фоне Env BG505, но эффекты мутаций на рост вируса и антигенность иногда различаются между вирусными штаммами (Barton et al., 2016; Falkowska et al., 2014; Haddox et al. , 2018).
Тем не менее, наличие полных антигенных карт BG505 Env по сравнению с панелью важных bnAbs дает огромное количество информации, которая может помочь в изучении эволюции ВИЧ и разработке противовирусных стратегий. Будущая работа, которая объединит эти антигенные карты с измерениями (Haddox et al., 2016, 2018) или моделями (Louie et al., 2018; Shekhar et al., 2013; Zanini et al., 2017) того, как мутации влияют на репликативную приспособленность ВИЧ. может пролить свет на эволюционную динамику вируса под давлением иммунной системы.
Методы
Создание мутантных вирусных библиотек
Ранее мы описали мутантные провирусные ДНК-библиотеки BG505.T332N и полученные функциональные мутантные вирусные библиотеки (Haddox et al., 2018). Вкратце, библиотеки мутантов BG505.W6M.C2.T332N env , содержащие рандомизированные мутации на уровне кодонов сайтов 31-702 (здесь и во всей рукописи используется нумерация HXB2), были независимо созданы и клонированы в провирус Q23.BsmBI.ΔEnv. плазмида (Haddox et al., 2018). Эти библиотеки провирусных плазмид, а также провирусные плазмиды дикого типа трансфицировали в клетки 293Т (полученные из АТСС). Затем мы пассировали супернатант трансфекции при MOI 0,01 инфекционных единиц TZM-bl на клетку в клетках SupT1.CCR5. Полученные в результате библиотеки мутантных вирусов, связанных генотип-фенотип, концентрировали с помощью ультрацентрифугирования.
Мутационное антигенное профилирование
Подход к мутационному антигенному профилированию Env был описан ранее (Dingens et al., 2017, 2018). Вкратце, 0,5 — 1 × 10 6 TZM-bl инфекционных единиц независимых мутантных вирусных библиотек нейтрализовали каждым антителом при концентрации ∼ IC 95 — IC 99,9 в течение одного часа. Затем библиотеки были инфицированы в 1 × 10 6 клеток SupT1.CCR5 в R10 (RPMI, с добавлением 10% FBS, 1% 200 мМ L-глутамина и 1% раствора 10 000 единиц / мл пенициллина и 10 000 мкг / мл). мл стрептомицина), содержащий 100 мкг / мл ДЭАЭ-декстрана. Параллельно с каждым отбором антител каждую библиотеку мутантных вирусов также инфицировали 1 × 10 6 SupT1.Клетки CCR5 без отбора антител, чтобы служить в качестве неотобранного для эксперимента контроля. Четыре 10-кратных серийных разведения каждой мутантной вирусной библиотеки также инфицировали 1 × 10 6 клеток в качестве стандартной кривой инфекционности. Клетки ресуспендировали в 1 мл свежего R10 через три часа после заражения. Клетки промывали PBS, осаждали и неинтегрированную вирусную ДНК выделяли с помощью минипрепарата через двенадцать часов после инфицирования. Затем был амплифицирован и секвенирован ген env с использованием метода секвенирования субампликона со штрих-кодом, как описано ранее (Haddox et al., 2018), который вводит уникальные молекулярные идентификаторы для уменьшения ошибки секвенирования. Количество вирусного генома в каждом образце определяли количественно с помощью кПЦР (Benki et al., 2006) или ddPCR (Dingens et al., 2017), и доля каждой выбранной библиотеки, которая выжила после отбора антител, по сравнению с невыбранным контролем составляла интерполированы из стандартной кривой инфекционности.
Анализ данных глубокого секвенирования и расчет выживающей фракции
Мы использовали dms_tools2 версии 2.2.9 (https: // jbloomlab.github.io/dms_tools2/) для анализа данных глубокого секвенирования и расчета выживающей фракции (Bloom, 2015). Расчет доли выживаемости был описан ранее (Doud et al., 2018) и подробно задокументирован на https://jbloomlab.github.io/dms_tools2/fracsurvive.html. Секвенирование плазмиды провирусной ДНК дикого типа использовали в качестве контроля ошибок при подсчете выживающей фракции. Мы взяли медианные значения для всех экспериментальных повторов для каждого антитела и нанесли данные о выживании избыточной фракции на логопрограммы, полученные с помощью dms_tools2, используя weblogo (Crooks et al., 2004) и ggseqlogo (Wagih, 2017).
Идентификация значимых участков утечки вируса
Поскольку соотношение сигнал / шум, по-видимому, различалось между антителами, мы определили статистически значимые участки утечки вируса за пределами фона индивидуально для каждого антитела. Для каждого антитела мы подгоняли гамма-распределение к значениям выживаемости фракции разделенных сайтов (средние значения для всех повторов), используя надежную регрессию (мягкая потеря L1, как реализовано в scipy). Затем мы определили сайты, которые выходили за пределы диапазона значений, ожидаемых от этого распределения, с коэффициентом ложного обнаружения 0.01 (Рисунок S2). Для целей фиг. 2B и 2C множественные сайты, которые нарушают PNG, с которыми связывается антитело, считались единым сайтом ускользания. Однако, когда антитело также контактировало со вторым или третьим остатками белка в PNG и этот сайт был местом значительного ускользания, этот сайт рассматривался как дополнительный сайт ускользания.
Структурный анализ
Сайты контакта антител определяли из структурных моделей Env-антитела. Были использованы следующие модели PDB: 5FYK для VRC01 (Stewart-Jones et al., 2016), 5V8M для 3BNC117 (Lee et al., 2017), 5T3Z для 10-1074 (Gristick et al., 2016), 3U4E для PG9 (McLellan et al., 2011), 5V8L для PGT145 (Lee et al. , 2017), 5FUU для PGT151 (Lee et al., 2016), 5I8H для VRC34.01 (Kong et al., 2016), 4G6F и 10E8 (Huang et al., 2012). Модели PGT121 с высоким разрешением, связанные с Env, недоступны; мы использовали модель PGT122 (PDB: 5FYL) (Stewart-Jones et al., 2016), которая является близкородственным «PGT121-подобным» клональным вариантом PGT121 (Mouquet et al., 2012). Контактные остатки были определены как любые остатки Env, где атом, не являющийся водородом, находится в пределах 4 Å от любого атома антитела, не являющегося водородом.Когда антитело контактировало с гликаном, N PNG этого гликана считали контактом. Для асимметричных структур антитело-тример использовали самое близкое расстояние из трех расстояний между антителом и остатками Env.
Для рисунков 3-5 мы использовали упомянутые выше модели (за одним исключением) для создания рисунков с помощью PyMol. Хотя мы использовали каркасную структуру PG9-V2 с высоким разрешением (McLellan et al., 2011) для определения сайтов контакта, мы картировали значения выживших фракций на модели умеренного разрешения PG9 в комплексе с тримером BG505 (PDB: 5VJ6) (Wang et al. al., 2017) на рис. 3F, чтобы лучше проиллюстрировать четвертичный аспект этого эпитопа вершины.
Анализы нейтрализации TZM-bl
Анализы нейтрализации TZM-bl с использованием псевдовирусов BG505.T332N, несущих одиночные дополнительные точечные мутанты, были выполнены и проанализированы, как описано ранее (Dingens et al., 2017). Анализ проводили в двух или трех независимых повторах, и кратное изменение IC 50 каждого мутанта относительно BG505. Псевдовируса дикого типа T332N рассчитывали независимо для каждого эксперимента, а затем усредняли по всем повторам.
Доступность данных и исходный код
Программное обеспечение с открытым исходным кодом для анализа наборов данных мутационного антигенного профилирования доступно по адресу https://jbloomlab.github.io/dms_tools2/. Вычислительный анализ представлен в виде исполняемого файла и записной книжки HTML Jupyter (файл S2) и по адресу https://github.com/jbloomlab/EnvsAntigenicAtlas, а значения выживших фракций и избыточных выживших фракций для каждого антитела представлены в виде файла S3. Чтения глубокого секвенирования Illumina доступны в NCBI SRA как исследование SRP157948, BioProject PRJNA486029, номера доступа SRX4553035-SRX4553088, SRX4614453.
Арипипразол улучшает симптомы депрессии и иммунологический ответ на антиретровирусную терапию у ВИЧ-инфицированного субъекта с устойчивой депрессией
Отчеты о клинических случаях в медицине 3
(и бупропион) проявляют такой эффект, что они оказывают
иммуномодулирующее и стимулирующее действие.
цитокинов и повышающие противовоспалительные цитокины [25].
Антидепрессанты, по-видимому, оказывают свое иммунологическое действие
как на центральном (ЦНС), так и на периферическом уровне (на активированных
макрофагах).Арипипразол также может быть эффективным у ВИЧ-
инфицированных пациентов в связи с определенным иммунологическим эффектом. Neu-
Ропсихиатрические расстройства у ВИЧ-инфицированных пациентов, по-видимому,
частично возникают в результате глиальной активации и высвобождения цито-
токсичных хемокинов и цитокинов, таких как интерферон-γ (IFN-γ)
[26 ]. In vitro арипипразол оказывает значительное ингибирующее действие на
-индуцированную активацию микроглии, снижая выработку
NO и TNF-α и подавляя повышение [Ca2 +] в микроглии на
[27].Мы предполагаем, что арипипразол
оказывает такое же действие in vivo на астроциты, активированные инфекцией.
Другой возможной причиной улучшения иммунного статуса пациента
является косвенное влияние зола арипипра-
на уровень кортизола, вызванное улучшением депрессии
. Нарушение регуляции функции оси HPA (например, повышение уровня адренокортикотропин-рилизинг-гормона на
и уровень кортизола на
) было связано со стрессом и депрессией
у людей, и такое нарушение регуляции может отрицательно повлиять на иммунный ответ.Имеются данные, подтверждающие тот факт, что
глюкокортикоиды могут влиять на патогенез ВИЧ напрямую,
через усиление репликации вируса или, более косвенно, через
, подавляя иммунный ответ на другие патогены. Фактически,
в 1997 году Christeff et al. [28] продемонстрировали, что высокие уровни
кортизола в плазме отрицательно связаны с количеством клеток CD4
. Таким образом, влияние арипипразола на репликацию ВИЧ и количество CD4
можно частично объяснить косвенным влиянием
на иммунный ответ, вызванным снижением уровня кортизола,
, в свою очередь, вызванным улучшением депрессивных симптомов.
В заключение, мы не можем исключить возможность того, что улучшение симптомов депрессии на
может привести к увеличению приверженности к антиретровирусной терапии на
.
На основе настоящего тематического исследования и обзора
литературы о влиянии психотропных агентов на репликацию вируса
, мы предполагаем, что использование арипипразола у ВИЧ-
инфицированных субъектов требует дальнейших исследований.
Список литературы
[1] R.Флёренс, Р. Уильямсон, Й. Цзин и др., «Систематический обзор
стратегий аугментации для пациентов с большим депрессивным расстройством
», Бюллетень психофармакологии, том. 42, pp. 57–90,
2009.
[2] М. Вуд и К. Ривилл, «Арипипразол действует как селективный частичный агонист дофаминового рецептора
», Заключение эксперта по
Investigational Drugs, vol. 16, нет. 6, pp. 771–775, 2007.
[3] М.А. Гаврилин, Л.Э. Матес, М.Поделл, «Метам-
фетамин усиливает клеточно-ассоциированный кошачий иммунодефицит
репликацию вируса в астроцитах», Journal of NeuroVirology, vol.
8, корп. 3, pp. 240–249, 2002.
[4] PJ Gaskill, TM Calderon, AJ Luers, EA Eugenin, J.
,A. Javitch и JW Berman, «Инфекция вируса иммунодефицита человека
макрофаги человека увеличиваются за счет дофамина
: мост между ВИЧ-ассоциированными неврологическими расстройствами
и злоупотреблением наркотиками », American Journal of Pathology, vol.
175, нет. 3, pp. 1148–1159, 2009.
[5] Ф. МакКенна, П. Дж. Маклафлин, Б. Дж. Льюис и др. «Экспрессия рецептора дофамина
на Т- и В-лимфоцитах человека, моно-
цитов, нейтрофилов, эозинофилов и NK-клеток: проточное цитометрическое исследование
, журнал нейроиммунологии, вып. 132, нет. 1-2, pp. 34–
40, 2002.
[6] Г.П. Кириллова, Р.Дж. Хруткай, М.Р. Шурин, Г.В. Шурин,
,И.Л. Туркова, М.М. Ванюков, «Дофаминовые рецепторы
в лимфоцитах человека: радиолиганд. связывание и количественный анализ
ОТ-ПЦР », Journal of Neuroscience Methods, vol.174, нет.
2, pp. 272–280, 2008.
[7] Л. Сантамброджио, М. Липартити, А. Бруни и Р. Даль Тосо,
«Дофаминовые рецепторы на Т- и В-лимфоцитах человека»,
Журнал нейроиммунологии, вып. 45, нет. 1-2, pp. 113–119,
1993.
[8] JW Berman, MJ Carson, L. Chang, et al., «NeuroAIDS,
, злоупотребление наркотиками и воспаление: создание совместных исследованиймероприятий. , ”Журнал нейроиммунной фармакологии, вып. 1, вып.
4, стр.351–399, 2006.
[9] M. E. Thase и A. J. Rush, «Когда сначала вы не добьетесь успеха:
последовательных стратегий для пациентов, не отвечающих на антидепрессанты», Jour-
nal of Clinical Psychiatry, vol. 58, приложение 13, стр. 23–29,
1997.
[10] Э.Г. Бинг, М.А. Бурнам, Д. Лонгшор и др., «Психиатрические
расстройства и употребление наркотиков среди людей с иммунодефицитом
, инфицированных вирусом. взрослых в Соединенных Штатах », Archives of General
Psychiatry, vol.58, нет. 8, pp. 721–728, 2001.
[11] J.Walkup, W.Wei, U.Sambamoorthi и S.Crystal, «Лечение прессантом Antide-
и соблюдение комбинированной антиретро-
вирусной терапии среди пациентов. СПИДом и диагнозом
депрессия, Psychiatric Quarterly, vol. 79, нет. 1, стр. 43–53,
2008.
[12] И.К. Энтони и Дж. Э. Белл, «Невропатология
ВИЧ / СПИД», Международный обзор психиатрии, том 20, № 1,
стр. 15–24, 2008.
[13] Дж. Дж. Дор, П. К. Коррелл, Й. Ли, Дж. М. Калдор, Д. А. Купер и
Б. Дж. Брю, «Изменения в комплексе деменции со СПИДом в эпоху
высокоактивной антиретровирусной терапии», СПИД, т. 13, вып. 10, pp.
1249–1253, 1999.
[14] С. А. Томас, «Распределение лекарств против ВИЧ в центральной нервной системе
», Current Pharmaceutical Design, vol. 10, вып.
12, pp. 1313–1324, 2004.
[15] Л. Э. Дэвис, Б. Л. Хьелле, В. Э. Миллер и др., «Ранняя вирусная инвазия
в мозг при ятрогенной инфекции вируса иммунодефицита человека
», Неврология, том. 42, нет. 9, pp. 1736–1739, 1992.
[16] S. Gartner и Y. Liu, «Понимание роли иммунной активации в нейропатогенезе ВИЧ», Journal of NeuroVirology,
vol. 8, вып. 2, pp. 69–75, 2002.
[17] К. К. Уильямс и У. Ф. Хики, «Повреждение центральной нервной системы
, моноциты и макрофаги, и неврологические расстройства
при СПИДе», Ежегодный обзор нейробиологии. т.25, pp.
537–562, 2002.
[18] CM Buckner, AJ Luers, TM Calderon, EA Eugenin и
JW Berman, «Нейроиммунитет и гематоэнцефалический барьер:
молекулярная регуляция лейкоцитов. трансмиграция и проникновение вируса
в нервную систему с акцентом на нейроСПИД »,
Journal of Neuroimmune Pharmacology, vol. 1, вып. 2, pp. 160–
181, 2006.
[19] S. Kramer-H¨
ammerle, I. Rothenaigner, H.Воль, Дж. Э. Белл,
и Р. Брак-Вернер, «Клетки центральной нервной системы как мишени
и резервуары вируса иммунодефицита человека»,
Virus Research, vol. 111, pp. 194–213, 2005.
[20] B. Arbaizar, T. Dierssen-Sotos, I. G ´
omez-Acebo, and J. Llorca,
«Арипипразол при большой депрессии и мании: метаанализ
Двухэтапный метод отбора проб для клинического надзора за лицами, получающими помощь в связи с ВИЧ-инфекцией в США, по JSTOR
АбстрактныйЦели: Цели этого исследования были двоякими: (1) описать методы составления популяционной выборки лиц, получающих помощь в связи с ВИЧ-инфекцией, и (2) сравнить данные из выборки с данными из существующих систем эпиднадзора, которые опишите помощь при ВИЧ.Методы. Авторы реализовали двухэтапный метод выборки, используя местные данные эпиднадзора за ВИЧ / СПИДом в качестве основы выборки поставщиков услуг по уходу за ВИЧ в трех штатах. У выбранных поставщиков были извлечены медицинские записи случайной выборки пациентов. Результаты: Были извлечены медицинские карты ряда пациентов, от 253 до 374 человек на штат. Демографические данные отобранных лиц и лиц, сообщенных в местную программу эпиднадзора за ВИЧ / СПИДом, были схожими; однако существовали различия в доле людей, получающих помощь в связи с ВИЧ в соответствии с руководящими принципами лечения, между выборкой и современным проектом дополнительного эпиднадзора в учреждениях.Средний эффект дизайна для исходов, собранных в выборке, составил 1,8 (диапазон = 0,5–29,6). Выводы. Этот метод обследования применим для сбора популяционных данных о пациентах, находящихся на лечении в связи с ВИЧ. Размер выборки и некоторые элементы дизайна следует изменить в будущих исследованиях, чтобы повысить точность оценок и полезность данных для местного планирования и оценки.
Информация о журналеPublic Health Reports — официальный журнал Службы общественного здравоохранения США.ASPH является партнером в публикации отчетов об общественном здравоохранении с 1999 года. Мы издаем этот рецензируемый журнал два раза в месяц — шесть выпусков, в которых публикуются статьи по трем основным направлениям: практика общественного здравоохранения, исследования и точки зрения / комментарии. За последние пять лет мы занялись такими темами, как борьба против табака, насилие среди подростков, профессиональные заболевания и травмы, иммунизация, политика в отношении наркотиков, скрининг на свинец, неравенство в состоянии здоровья и многие другие ключевые вопросы. Авторы журнала находятся на переднем крае общественного здравоохранения, и мы представляем их работы в удобочитаемом и доступном формате.
Информация об издателеСара Миллер МакКьюн основала SAGE Publishing в 1965 году для поддержки распространения полезных знаний и просвещения мирового сообщества. SAGE — ведущий международный поставщик инновационного высококачественного контента, ежегодно публикующий более 900 журналов и более 800 новых книг по широкому кругу предметных областей. Растущий выбор библиотечных продуктов включает архивы, данные, тематические исследования и видео. Контрольный пакет акций SAGE по-прежнему принадлежит нашему основателю, и после ее жизни она перейдет в собственность благотворительного фонда, который обеспечит дальнейшую независимость компании.Основные офисы расположены в Лос-Анджелесе, Лондоне, Нью-Дели, Сингапуре, Вашингтоне и Мельбурне. www.sagepublishing.com
.